广西巴马小型猪内源性反转录病毒整合后传代次数对HEK293细胞中HERV-W表达和变异的影响
2020-11-06秦树英白安斌陈凤莲覃绍敏刘金凤吴健敏
马 玲,钟 华,秦树英,白安斌,陈凤莲,覃绍敏,刘金凤,吴健敏*
(1.广西壮族自治区兽医研究所 广西兽医生物技术重点实验室,广西 南宁 530001;2.广西民族大学,广西 南宁530006)
近年来,异种器官移植取得了重大进展,如移植转基因猪心脏的狒狒可以存活195 d,新西兰科学家已经成功制备SPF猪可以用于异种器官移植等。如果猪→人异种器官移植可以应用于临床,将极大地缓解器官移植供体严重短缺的现状。但整合在猪基因组中的猪内源性反转录病毒(porcine endogenous retrovirus,PERV)却一直制约着异种器官移植的发展[1-3]。PERV属于反转录病毒科γ反转录病毒属,与小鼠白血病病毒(murine leukemia virus,MLV)、猫白血病病毒(feline leukemia virus,FeLV)等病毒同属,可以在体外感染猪、犬、猫、蝙蝠及多种灵长类动物细胞和十余种人源细胞。此外,PERV还可以在体内感染重度联合免疫缺陷(severe combined immunodeficiency,SCID)裸鼠,纯化的病毒TM蛋白可以显著抑制人类免疫细胞功能,PERV的感染可以改变宿主细胞因子的表达水平,以及2002年滴度更高、嗜性更广的PERV-A/C重组病毒的发现,都引起人们对异种器官移植的担忧——PERV是否可以突破种间屏障,通过异种器官移植造成猪→人的跨种传播[2,4-9]。
对PERV另一方面的担忧,是PERV与人逆转录病毒(human endogenous retrovirus,HERV)的重组及其对HERV表达的影响。从2000年开始,有报道PERV在人类细胞基因组中整合后,有可能与HERV发生重组[7,10];MACHNIK等[11]和马玲等[12]研究表明PERV的整合会影响HERV-W的表达。HERV-W是HERV家族中与γ反转录病毒同源关系较高的,其编码syncytin-1蛋白的ERVWE1位于7号染色体,而PERV-BM在7号染色体上恰有2个整合位点,PERV-BM的整合如何影响不同代次细胞感染模型中的HERV-W,尤其syncytin-1的表达,是否会造成HERV-W的变异还有待深入研究[13-14]。
有研究表明,广西巴马小型猪来源的PERV(PERV-BM)整合到HEK293细胞后,宿主细胞的细胞周期、细胞凋亡和相关调节基因的表达会发生改变,且与细胞感染模型的传代次数有关[15-16]。本试验在马玲等[12]研究的基础上,扩大检测范围,检测不同代次HEK293-PERV-BM中HERV-W各结构基因mRNA和syncytin-1蛋白表达水平;同时测定不同代次HEK293-PERV-BM中HERV-W 各结构基因的序列,监测PERV-BM整合后HERV-W的变异情况;为进一步明确PERV在异种移植中的安全性提供参考。
1 材料与方法
1.1 细胞系和细胞感染模型HEK293、PK15细胞由本实验室保存;PERV-BM感染HEK293细胞模型(HEK293-PERV-BM)和PK15细胞来源猪内源性反转录病毒(PERV-PK15)感染HEK293细胞模型(HEK293-PERV-PK)均由本实验室构建、培养、保存[11]。待各细胞、细胞感染模型长成单层后,进行细胞传代,约3~4 d传代1次,每5代收集细胞,用于检测。
1.2 主要试剂DMEM培养基、胎牛血清为Thermo Fisher Scientific产品;QIAamp DNA Mini Kit 为QIAGEN Bioinformatics产品;MiniBEST Universal RNA Extraction Kit、PrimeScriptTMRT Master Mix和SYBR Premix Ex Taq Ⅱ为TaKaRa宝生物工程(大连)有限公司产品;RIPA裂解液、Bradford蛋白浓度测定试剂盒为上海碧云天生物技术有限公司产品;100×蛋白酶抑制剂复合物、100×磷酸酶抑制剂混合物Ⅰ、anti-β-actin鼠单克隆抗体、HRP标记的山羊抗小鼠IgG为生工生物工程(上海)股份有限公司产品;anti-syncytin-1抗体和山羊抗兔IgG抗体为Abcam艾博抗(上海)贸易有限公司产品;增强型DAB显色试剂盒(20×)为北京索莱宝科技有限公司产品;其他化学试剂均为国产分析纯。
1.3 不同代次HEK293-PERV-BM中HERV-W mRNA实时荧光定量PCR检测在1~35代HEK293-PERV-BM细胞感染模型中,每隔5代抽提1次RNA,采用已建立的HERV-W 实时荧光定量PCR方法,检测其中HERV-W各基因mRNA相对表达量,同时设HEK293细胞为对照,每个样品进行3次平行试验,并对检测结果进行数据分析。
1.4 不同代次HEK293-PERV-BM中syncytin-1 Western blot检测根据HERV-W实时荧光定量PCR检测结果,收集mRNA相对表达量改变明显代次的细胞,取等量蛋白样品,进行Western blot检测,分析各代次中syncytin-1蛋白表达水平。
1.5 不同代次HEK293-PERV-BM中HERV-W结构基因克隆与分析
1.5.1引物合成 根据GenBank收录的HERV-W序列(GenBank登录号:AY101582、AY101583、AH013147、AF520540、AF127229、AF072506、AF123881),设计合成扩增HERV-W的gag、pol和env结构基因全长的特异性引物(表1),并由北京博迈德基因技术有限公司合成。
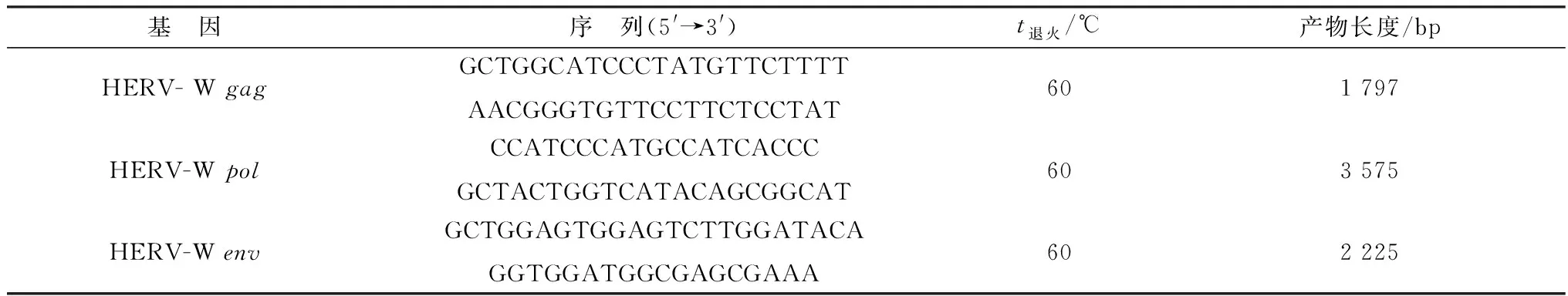
表1 HERV-W gag、pol和env基因扩增引物信息
1.5.2DNA抽提、目的基因扩增及序列比对 每隔5代抽提HEK293-PERV-BM P1~P55细胞基因组DNA,以各代次细胞感染模型的基因组DNA为模板,扩增HERV-W结构基因gag、pol和env全长,克隆扩增产物纯化回收后送生工生物工程(上海)股份有限公司测序。测序结果分别与HERV-W序列(GenBank登录号:AY101582、AY101583、AH013147、AF520540、AF127229、AF072506、AF123881)进行比对,并与AY101582进行相似性分析。
2 结果
2.1 不同代次HEK293-PERV-BM中HERV-W mRNA表达分析应用SYBR GreenⅠ实时荧光定量PCR分别检测HEK293和HEK293-PERV-BM中HERVgag、HERVpol、HERVenv、syncytin-1和humanβ-actin的mRNA表达水平。结果如图1所示,与HEK293细胞相比,PERV整合后,不同代次HEK293-PERV-BM中HERV-Wgag、HERVpol、HERVenv和syncytin-1的mRNA相对表达水平出现不同程度的改变,且改变趋势相似,即从P1开始出现轻微改变,P10或P20后,gag、pol、env和syncytin-1相对表达量升高,至P30代最高,达3.72%~37.27%,之后开始下降至与P10或P20代相近的水平。
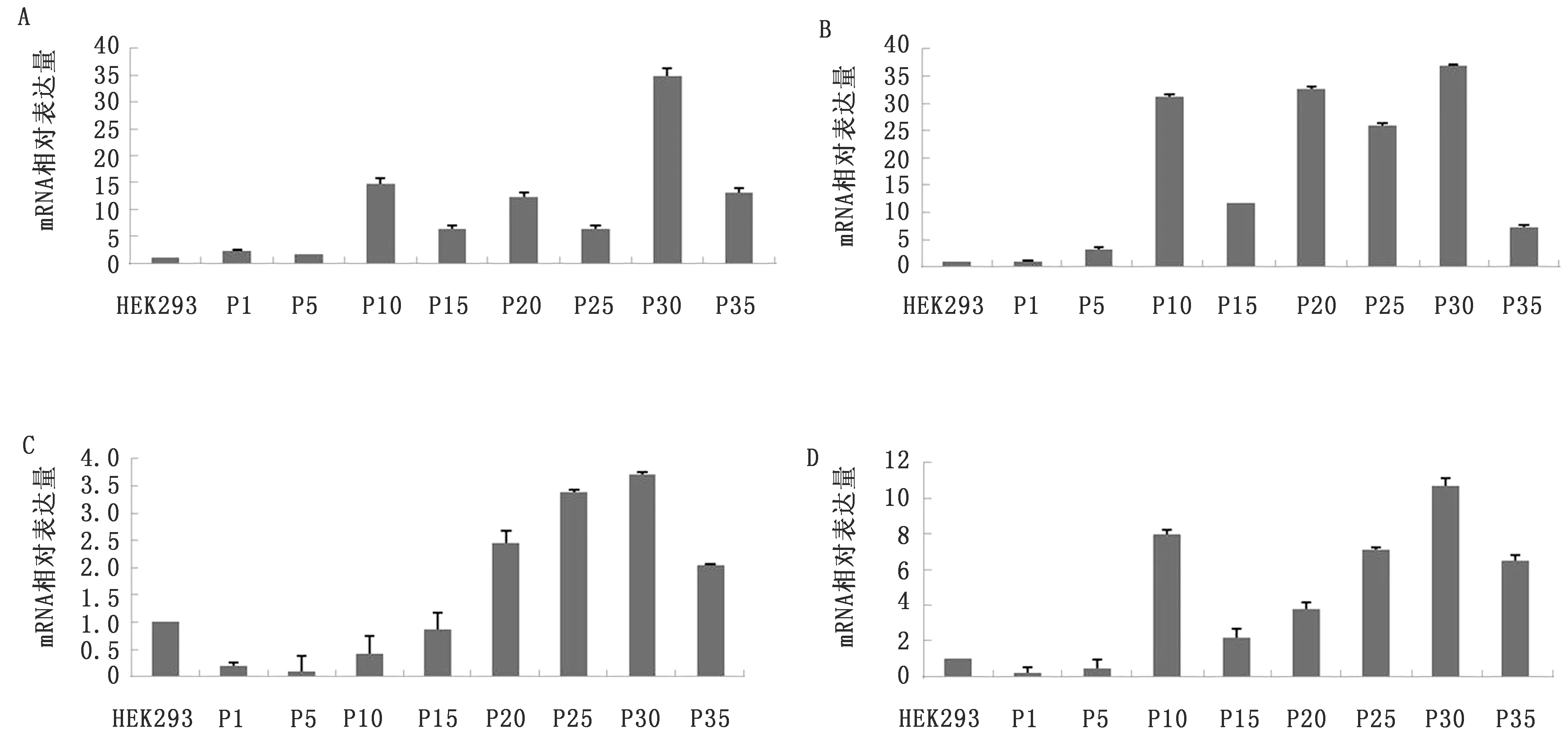
图1 各代次HEK293-PERV-BM细胞感染模型中HERV-W gag(A)、HERV-W pol(B)、HERV-W env(C)和syncytin-1(D) mRNA相对表达量
2.2 不同代次HEK293-PERV-BM中syncytin-1蛋白表达水平分析按照HERV-W实时荧光定量PCR结果,选取相对表达量差别较大的第1,15,25,30代HEK293-PERV-BM进行Western blot检测,同时设HEK293和HEK293-PERV-PK为对照。结果如图2所示,试验组和对照组均出现55 000的HERV-W syncytin-1特异性条带和42 000的β-actin特异性条带。与HEK293相比,HEK293-PERV-BM P1的syncytin-1条带较弱、HEK293-PERV-BM P15表达量相当、HEK293-PERV-BM P25和P35表达量稍高,而HEK293-PERV-PK蛋白表达量稍高。比较发现,除P1代外,各代次间HERV-W syncytin-1蛋白表达量的差异不大,均较相应mRNA差异小。
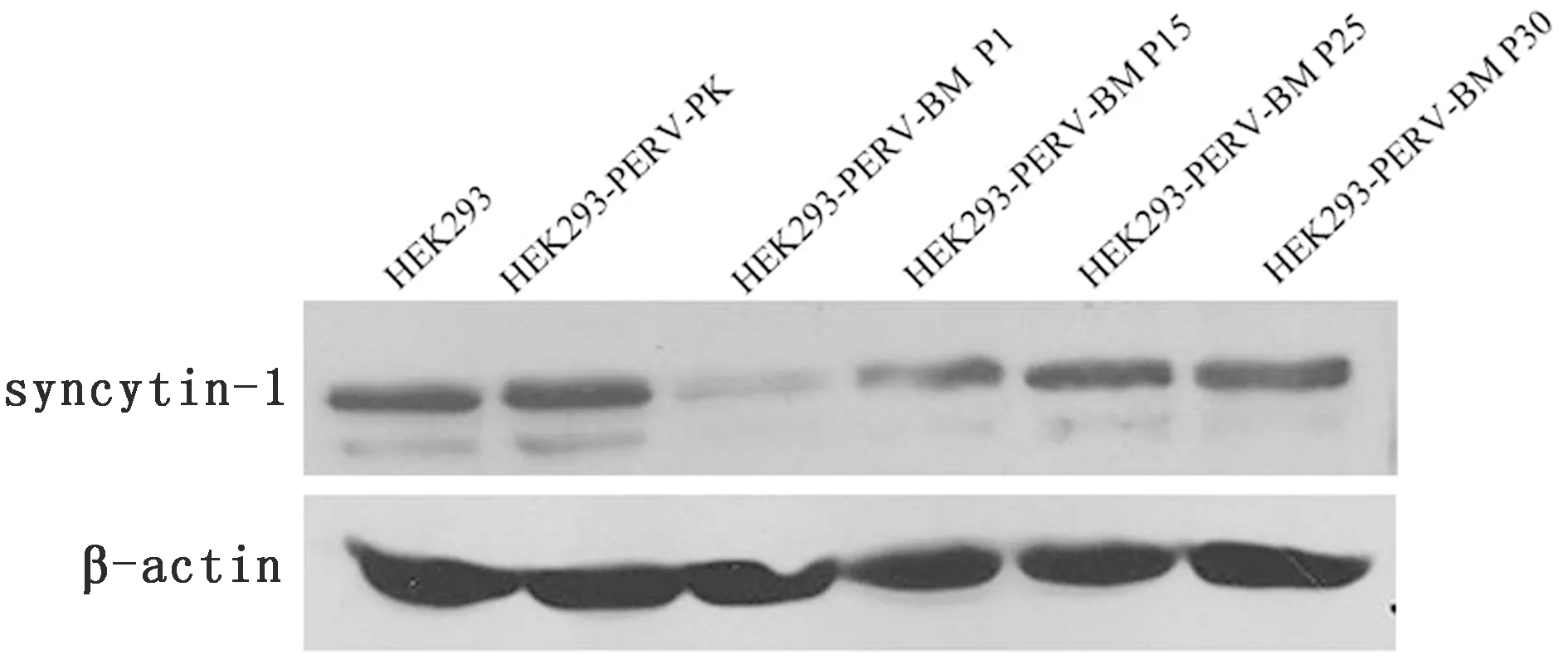
图2 不同代次HEK293-PERV-BM细胞感染模型中syncytin-1的Western blot检测
2.3 不同代次HEK293-PERV-BM中HERV-W结构基因克隆与变异分析以每隔5代抽提HEK293-PERV-BM细胞感染模型P1~P55基因组DNA为模板,分别扩增HERV-Wgag、pol、env基因。如图3 所示,各代次HEK293-PERV-BM细胞感染模型的HERW-Wgag、pol和env基因序列与AY101582相似性分别为100.0%,98.5%~100.0%和99.5%~100.0%。从第1~55代细胞感染模型中,HERV-Wgag、pol和env基因的变异主要表现在个别位置的点突变,各代次中gag基因相似性为100%,无变异;pol的变异主要集中在P5代的1 626~1 811 bp;env基因的变异主要从P20代开始,P40和P45中均有个别位点发生突变;而在920位,各代次HERW-Wenv均为A,而AY101582为G,这可能是HEK293细胞株的特点。由此可见,1~55代HEK293-PERV-BM中HERV-W 结构基因仅发生个别位置的点突变,未见发生固定位置的点突变或基因片段的缺失、插入及重组。

图3 不同代次HEK293-PERV-BM细胞感染模型中HERV-W gag(A)、pol(B)、env(C)序列比对
3 讨论
本试验发现,PERV-BM整合后,不同代次HEK293-PERV-BM中HERV-W各基因mRNA和syncytin-1蛋白表达均发生不同程度的改变,且与代次有关。另外,HEK293-PERV-BM P1~P55中HERV-Wgag、pol和env结构基因仅发生个别位置的点突变,未出现规律性点突变或基因片段的缺失、插入及重组。
病毒感染会影响HERV的转录,如流感病毒A/WSN/33、单纯疱疹病毒感染后可以激活HERV-W的转录,乙型肝炎病毒(hepatitis B virus,HBV)可以通过NF-κB诱导HERV-Wenv基因的过表达;此外,人类免疫缺陷病毒1型(human immunodeficiency virus-1,HIV-1)感染后可以激活HERV-K 前病毒,造成mRNA和蛋白表达水平升高,并可以在患者血液中检出病毒样粒子;人4型疱疹病毒也可以激活HERV-W和HERV-K18的表达[17-21]。PERV-PK的整合不会影响HERV-K的转录谱,但是却会影响HERV-W各基因mRNA和syncytin-1蛋白的表达,而且在特定代次之间存在显著性差异;PERV-BM的整合也会造成HERV-W各基因mRNA表达水平明显升高,上述结果与本试验结果类似,而且于P10或P20开始升高,至P30达到最高,但syncytin-1蛋白的表达,则是除P1外,其他代次差异不大[11-12]。此外,HERV-Wenv基因与自身免疫性疾病、精神分裂症、Ⅰ型糖尿病等多种人类疾病的发生发展有直接关系,其抗原表位的W、Q和T肽可以诱导强烈的T细胞免疫应答,抗HERV-Wenv的单克隆抗体GnbAC1由于对Ⅰ型糖尿病有明显治疗效果,正处于临床开发阶段[22-23]。HERV-Wenv基因可以编码syncytin-1蛋白,位于7q21.2,是一种编码合胞素基因蛋白,在人胎盘的生长过程中至关重要,其低表达可导致胎盘发育不良、自然流产等,而该蛋白的异常表达也可见于乳腺癌、子宫内膜癌、T细胞淋巴瘤等多种恶性肿瘤中[11]。除此之外,syncytin-1还具有抗凋亡、介导细胞(滋养细胞和癌细胞)融合、抑制识别相同受体的外源性逆转录病毒感染细胞等功能[13,24-26]。所以PERV整合后,HERV-W各基因表达的改变可能对人类存在一定风险。
OLDMIXON等[9]在NIH小型猪中发现了A型和C型PERV重组形成PERV-A/C病毒,其为滴度和感染性更强的重组病毒,而且PERV-BM在7号染色体上的整合与HERV-W位置较近,更加剧了对PERV与HERV重组的担忧。本试验结果表明,PERV-BM整合后,HERV-W 结构基因仅发生个别位置的点突变,未见发生固定位置的点突变或基因片段的缺失、插入及重组,这与SULING等[27]推测PERV与HERV-W、K、E、R等发生重组的几率较低的结果一致。但是随着时间的推移,HERV积累的突变有可能会破坏其编码能力,所以仍应引起重视[28]。
