E.tenella 河北株SO7基因在毕赤酵母系统中的表达及免疫保护力评价
2020-11-06闫艳娟李蕴玉张东林李佩国庞洪泽张香斋
闫艳娟,李蕴玉,张东林,李佩国,庞洪泽,张香斋
(河北科技师范学院 河北省预防兽医学重点实验室,河北 秦皇岛 066600)
鸡球虫病是由一种或几种艾美耳球虫引起的寄生性原虫病,该病严重危害养禽业,造成巨大的经济损失[1-3]。目前鸡球虫病的防治仍以化学药物为主,其中最常用的抗球虫药一类是聚醚类离子载体药物,如马杜霉素、拉沙霉素等;另一类即为化学合成类的抗球虫药,主要有地克珠利等[4]。但由于化学药物使用不当,导致耐药虫株的出现及药物残留威胁人类的健康,因此,核酸疫苗的研究成为了热点。有研究表明球虫多种细胞器所分泌的蛋白质与虫体侵入宿主细胞有重要联系[5]。
SO7基因是在子孢子和第1代裂殖体阶段都存在的一种折光体抗原,有研究表明E.tenellaSO7基因的表达产物对E.tenella有一定的免疫保护作用[6-7],但关于E.tenellaSO7基因在毕赤酵母系统中的表达产物对艾美耳球虫的免疫保护效果还未见报道。本试验对E.tenella河北株SO7基因进行克隆,并与IL-2基因及真核表达载体pPIC9串联,转染毕赤酵母细胞GS115,进而研究体外重组蛋白SO7对鸡E.tenella的保护作用,为新型鸡球虫疫苗的研制提供依据。
1 材料与方法
1.1 虫株、质粒、细胞E.tenella河北株孢子化卵囊、IL-2基因、真核表达载体质粒pPIC9及毕赤酵母细胞GS115均由河北省预防兽医学重点实验室保存。
1.2 主要试剂与仪器质粒提取试剂盒、胶回收试剂盒购自OMEGA公司;Ex Taq DNA 聚合酶、DL2000 DNA Marker、限制性内切酶SacⅠ、EcoRⅠ、BamHⅠ、SacIⅠ、T4DNA 连接酶、His标签单克隆抗体、HRP标记山羊抗小鼠IgG抗体均购自 TaKaRa生物工程(大连)有限公司。His选择镍-亲和层析柱购自GE Healthcare公司;梯度PCR仪由德国eppendorf AG公司生产;MIKRO 220R型台式高速冷冻离心机由德国Heettich公司生产。
1.3E.tenella河北株SO7基因的克隆与鉴定根据GenBank登录的E.tenellaSO7基因的序列(KX826909.1),设计1对特异性引物,P1:5′-ATACGCGTGCAGACCTCTTCAGCGGAC-3′;P2:5′-ATCCGCGGTTACTTATCGTCGTCATCCTTGTAATCCCAGAAGAAGCCGGAG-3′。由生工生物工程(北京)股份有限公司合成,用RT-PCR方法克隆E.tenellaSO7基因,回收目的基因片段与pMD18-T载体连接,转化到DH5α感受态细胞中,筛选阳性质粒,用EcoRⅠ和BamHⅠ限制性内切酶双酶切鉴定。
1.4 真核表达载体pPIC9-IL-2-SO7的构建将IL-2基因片段与真核表达载体pPIC9于37℃ EcoRⅠ/BamHⅠ同时双酶切,回收酶切产物,用T4DNA连接酶进行连接,转入DH5α感受态细胞中,挑取阳性克隆进行PCR检测、序列测定,无突变和移码即得到重组表达载体pPIC9-IL-2。再将回收的SO7基因片段与重组表达载体pPIC9-IL-2同时用MluⅠ/SacⅡ进行双酶切,回收酶切产物,以T4DNA连接酶连接,转染DH5α感受态细胞,挑取阳性克隆进行PCR检测、序列测定,测序正确即得到真核表达载体pPIC9-IL-2-SO7。
1.5 毕赤酵母细胞的转化及阳性菌株的筛选和鉴定利用限制性内切酶SacⅠ 对真核重组质粒pPIC9-IL-2-SO7进行线性化处理,LiCl转化法转化毕赤酵母GS115感受态细胞,取50 μL制备好的毕赤酵母GS115感受态细胞。加入50% PEG3350 240 μL、1 mol/L LiCl 36 μL、2 g/L单链Salmon sperm DNA 25 μL和线性化的质粒 50 μL,30℃水浴30 min,42℃水浴热休克20~25 min,6 000 r/min离心收集菌体。用1 mL YPD置30℃摇床孵育1~4 h 后,离心取100 μL并涂RDB平板,培养3~4 d 至长出白色菌落。同时转化空载体pPIC9作为对照。提取酵母细胞基因组,针对酵母细胞AOX1基因设计特异性引物P1:5′-GACTGGTTCCAATTGACAAGG-3′,P2:5′-GGCAAATGGCA-TTCTGACATCCT-3′,PCR筛选阳性菌株。同时用毕赤酵母GS115感受态细胞作为空白对照。
1.6 重组蛋白SO7的表达和纯化验证筛选出阳性菌株后,接种于BMMY中,用甲醇诱导表达,采用His选择镍-亲和层析柱分离纯化,获得的蛋白按1/4体积比例加入4×Loading Buffer,充分混匀后煮沸10 min,进行SDS-PAGE电泳检测;应用Western blot方法验证目的蛋白的免疫原性,以His标签单克隆抗体为一抗,以HRP标记山羊抗鸡IgG为二抗。所表达纯化的蛋白即为重组蛋白SO7。
1.7 动物保护性试验将150只7日龄雏鸡逐只测体质量,按其体质量离散度随机分为5组,每组3个重复,每个重复10只,1,2组分别为阴性对照组和阳性对照组,3~5组每只鸡在7,14,21日龄时分别胸肌注射50,100,150 μg的重组蛋白SO7,28日龄时除阴性对照组外的所有雏鸡口服8×104个E.tenella孢子化卵囊,期间观察记录各组鸡只的精神状态、食欲及排泄物状况;对死亡鸡只剖检并记录死亡原因。试验至42日龄时,测体质量,每组取5只鸡剖杀,参照文献[8]方法进行盲肠病变计分和盲肠内容物卵囊值,计算其抗球虫指数(ACI)。ACI在180以上为优秀,160~180为良好,120~160为较差,120以下为无效。
2 结果
2.1E.tenella河北株SO7基因的克隆与鉴定以E.tenellaSO7基因的引物P1/ P2扩增的SO7基因,10 g/L琼脂糖凝胶电泳分析RT-PCR产物,可见1条约600 bp的条带,与预期条带大小相符(图1)。对筛选得到的阳性重组质粒pMD18-T-SO7应用BamHⅠ和EcoRⅠ进行双酶切鉴定,结果显示有2条特异性条带,分别为约600,2 600 bp的载体片段,与理论结果一致,说明E.tenella河北株的SO7基因成功克隆到pMD18-T载体中(图2)。
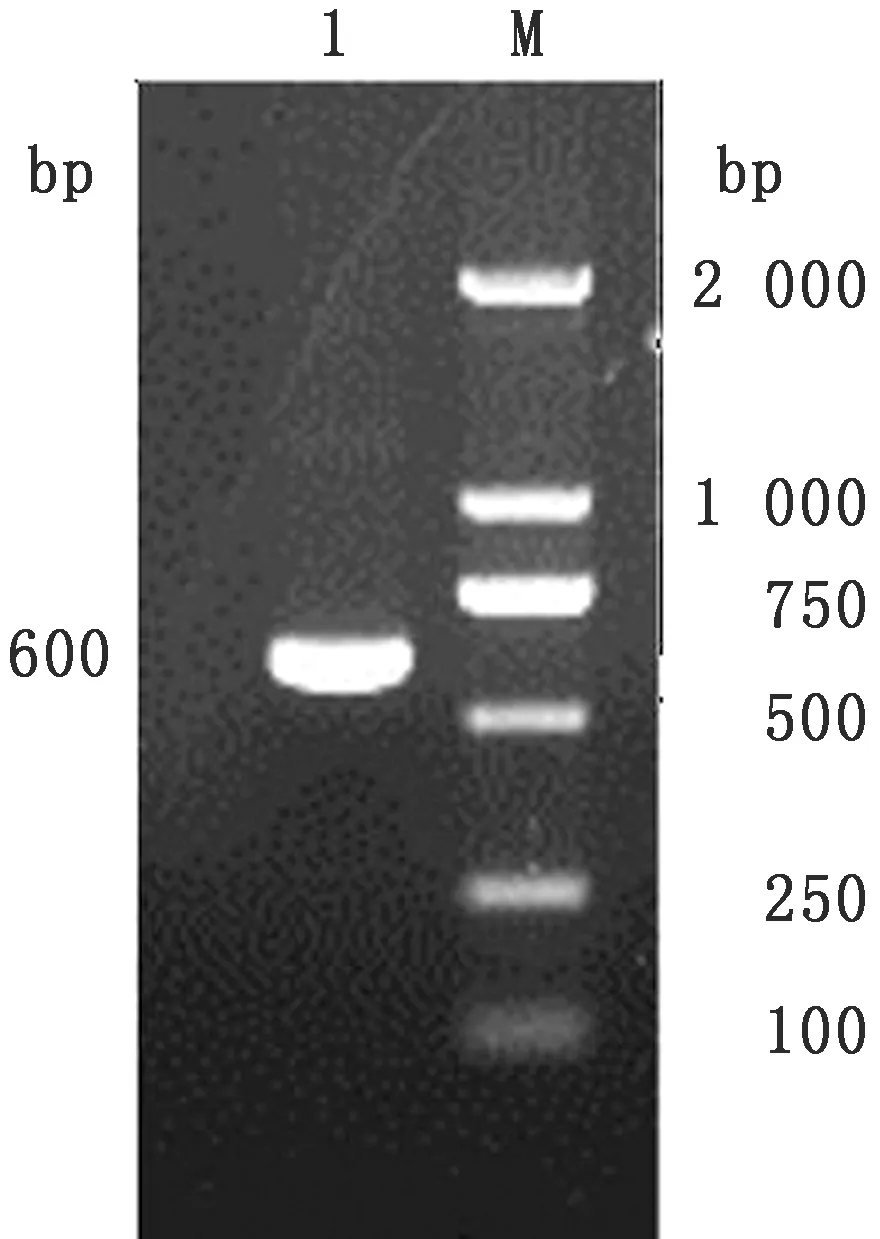
图1 E.tenella 河北株SO7基因扩增结果 1.E.tenella SO7基因;M.DL2000 DNA Marker
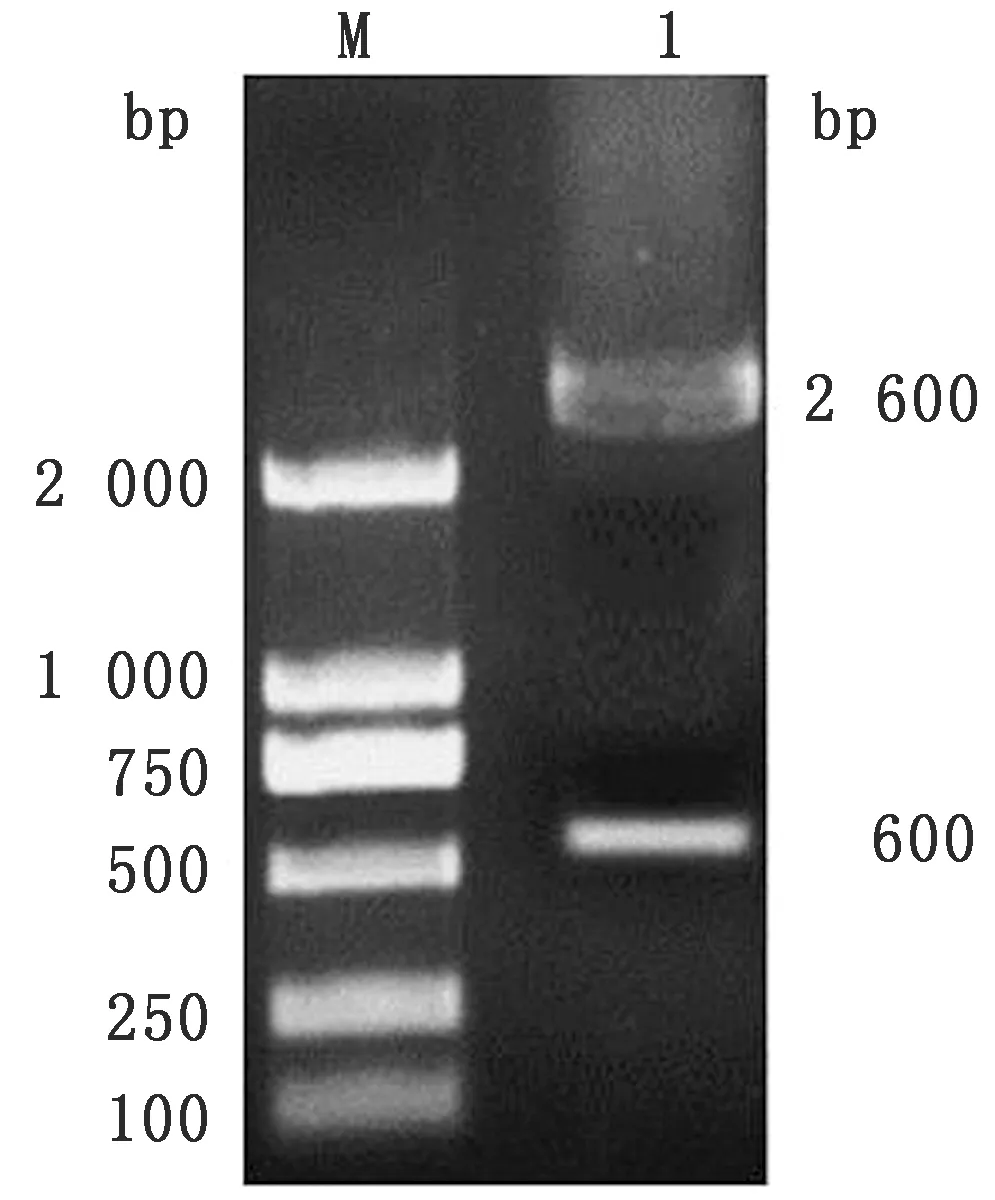
图2 E.tenella 河北株SO7基因双酶切鉴定结果 1.SO7基因双酶切产物;M.DL2000 DNA Marker
2.2 真核表达载体pPIC9-IL-2-SO7的构建将IL-2基因用T4DNA连接酶连接到表达载体pPIC9中,构建真核表达载体pPIC9-IL-2,转入DH5α感受态细胞中,挑取阳性克隆进行PCR检测,获得大小为432 bp的目的基因片段(图3),序列测定结果该基因无突变和移码。将SO7基因酶切回收产物用T4DNA连接酶与真核表达载体pPIC9-IL-2相连,构建真核表达载体pPIC9-IL-2-SO7转入DH5α感受态细胞中,挑取阳性克隆进行PCR检测,获得约600 bp的目的基因片段(图4),序列测定结果该基因无突变和移码,表明真核表达载体pPIC9-IL-2-SO7构建成功。
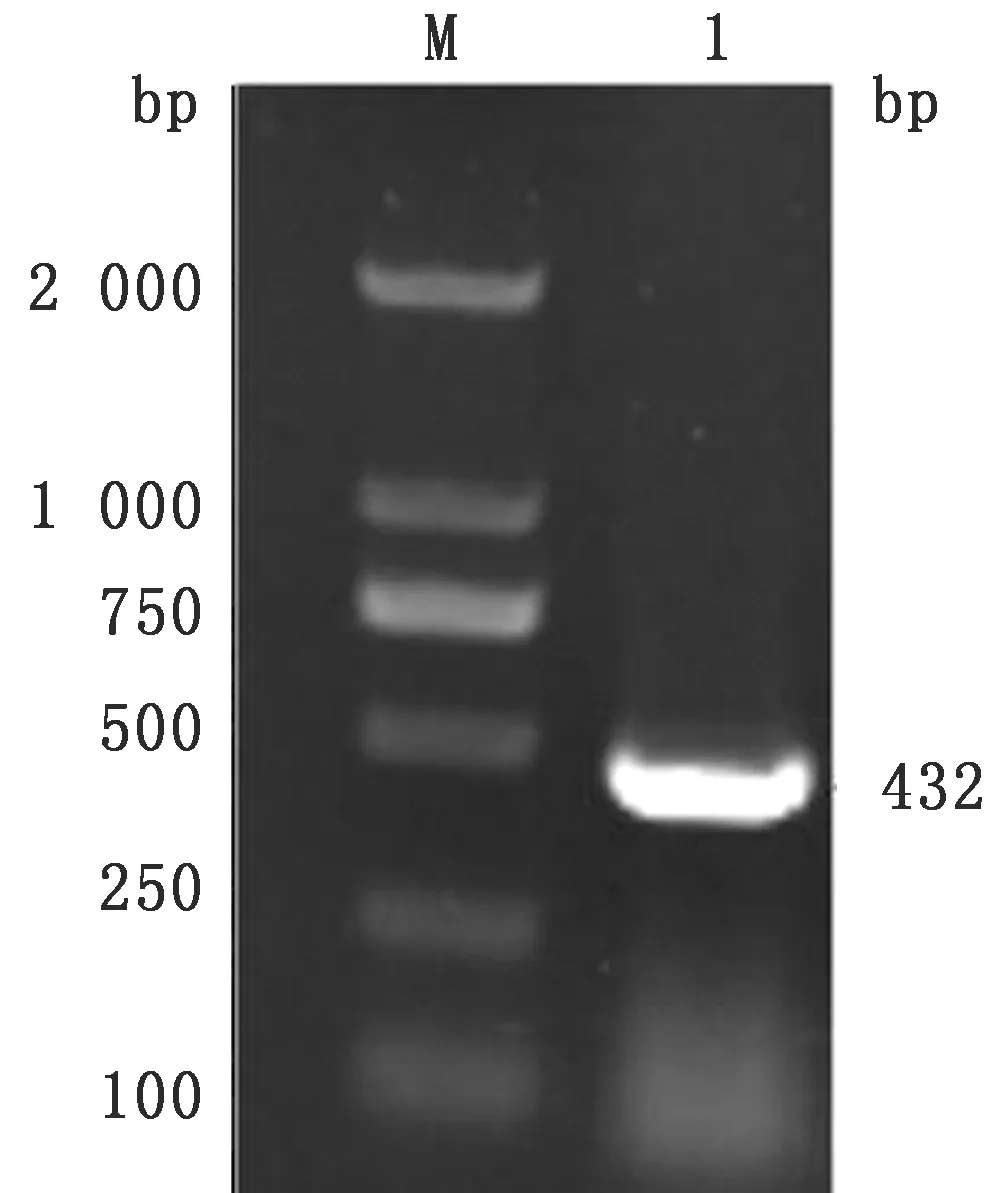
图3 真核表达载体pPIC9-IL-2的PCR鉴定结果 M.DL2000 DNA Marker;1.IL-2基因

图4 真核表达载体pPIC9-IL-2-SO7的PCR鉴定结果 1.SO7基因;M.DL2000 DNA Marker
2.3 毕赤酵母表达载体GS115-pPIC9-IL-2-SO7的构建转化酵母细胞GS115后,经过RDB培养基的筛选,得到转化后的阳性菌株。用酵母细胞特有的引物AOX1进行PCR鉴定,结果显示,E.tenella河北株SO7基因重组到酵母细胞基因组的阳性菌株有2条带,分别为2 200,1 100 bp,对照组毕赤酵母细胞GS115有1条带,为2 200 bp(图5);表明将E.tenella河北株 SO7基因成功整合到毕赤酵母细胞的基因组中,其阳性菌株命名为GS115-pPIC9-IL-2-SO7。
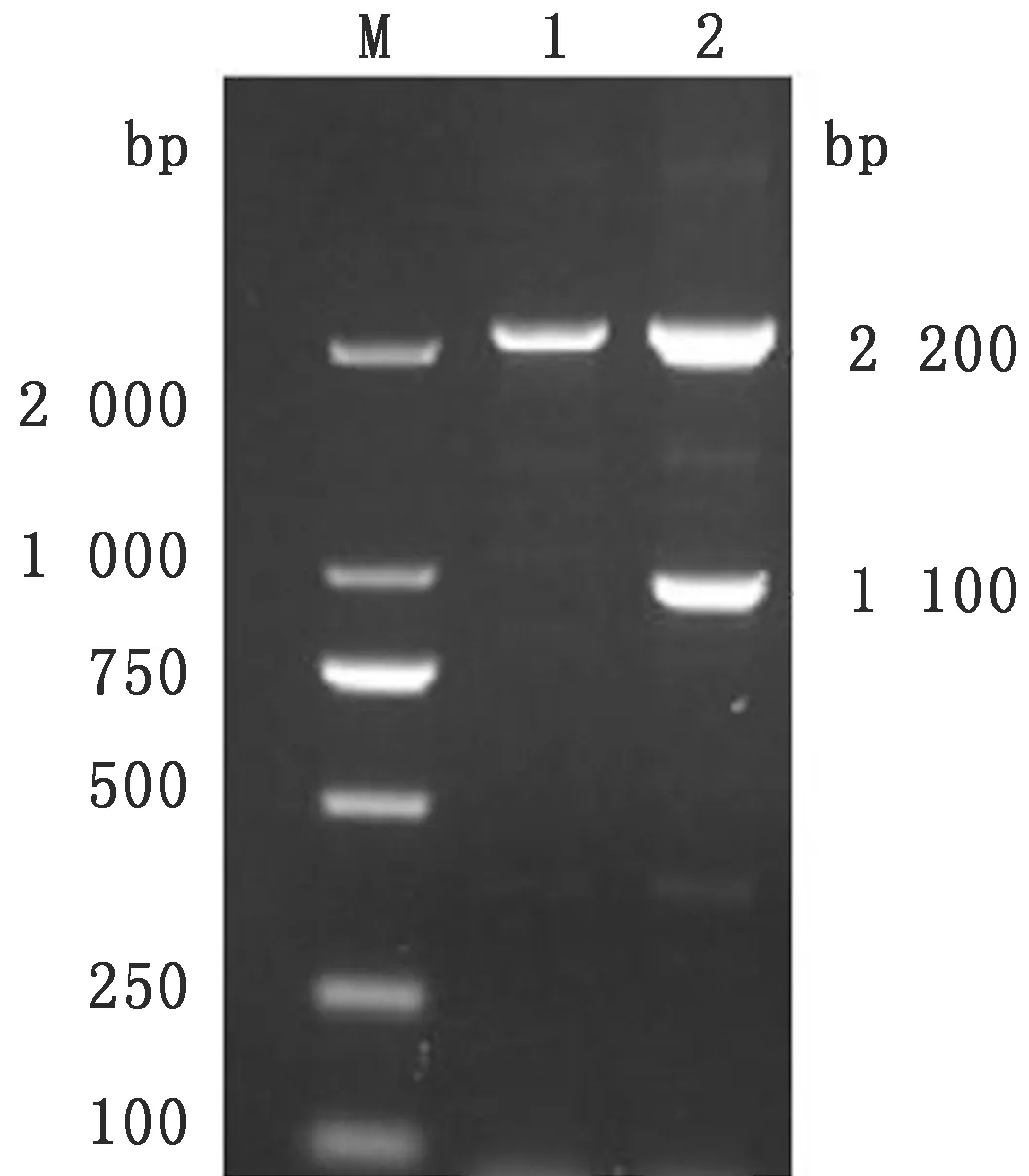
图5 阳性菌株GS115-pPIC9-IL-2-SO7的PCR鉴定结果 M.DL2000 DNA Marker;1.GS115对照;2.阳性菌株GS115-pPIC9-IL-2-SO7
2.4 重组蛋白SO7的表达和纯化验证甲醇诱导毕赤酵母细胞表达蛋白,采用His选择镍-亲和层析柱对所表达的蛋白进行纯化,应用SDS-PAGE电泳对所表达及纯化蛋白进行检测。以鸡源HIS标签单克隆抗体为一抗,山羊抗鸡IgG-HRP为二抗进行Western blot检测。SDS-PAGE电泳检测结果显示,纯化前蛋白条带中存在与预期蛋白大小相符的条带,但具有多条杂带,纯化后除目的蛋白外基本无其他杂带(图6A)。Western blot检测结果显示,在与理论值大小一致的位置出现了清晰条带,无其他杂带(图6B),表明已成功表达了重组蛋白SO7,且重组蛋白具有很好的生物活性。

图6 重组蛋白SO7表达及纯化的SDS-PAGE(上)及Western blot(下)检测结果 M.蛋白 Marker;1.空载体;2.1次纯化;3~5.多次纯化;6.纯化前;7.重组蛋白SO7
2.5 免疫保护力评价由表1可以看出,在试验期间,各组存活率均为100%,1组即阴性对照组鸡只精神状态、食欲及粪便无异常,其他各组在攻虫后出现不同程度的精神沉郁、血便,但未出现死亡。与阳性对照组比较,3~5组的相对增重率分别提高了39.82%,44.15%,33.66%;病变值和卵囊值分别降低了16.67%,25.00%,16.67%和90%,90%,0%。若以ACI作为抗球虫效果综合评价指标,其ACI比阳性对照组分别提高了29.66%,33.33%和19.72%,其中按50,100 μg/只的剂量进行免疫时,ACI均高于160,具有良好的抗球虫水平。

表1 抗球虫指数(ACI)
3 讨论
鸡球虫具有复杂的生活史和庞大的基因组,并且在不同的发育阶段具有不同的抗原和免疫原性,有研究证实,鸡球虫在裂殖子和子孢子阶段均具有高免疫原性,能使机体产生保护性免疫[9]。SO7基因是在球虫子孢子阶段出现的一种折光体抗原,因其表达产物对鸡具有免疫保护作用,可以作为构建核酸疫苗候选基因[10]。王卉等[11]扩增出E.tenella杨凌株的SO7基因,并进行原核表达,结果显示SO7重组蛋白可以很好地保护鸡只免受球虫侵害,卵囊产量下降率为46.81%,ACI达160.8。
IL-2是T淋巴细胞产生的一种细胞因子,有研究表明IL-2具有促进机体免疫反应,增强疫苗效果的作用,国内先后有使用IL-2作为免疫增强佐剂,增强疫苗效果并取得成功的先例[12]。SHAH等[13]将IL-2等细胞因子与堆型艾美耳球虫pcDNA3-1E联合应用,结果表明,IL-2等细胞因子可明显增强pcDNA3-1E免疫的效果。SONG等[14]构建了2种DNA疫苗pVAX-SO7和pVAX-SO7-IL-2,分别免疫鸡只,观察2种疫苗对抗球虫的效果,结果显示2种疫苗可提高增重率,降低盲肠病变计分,同时对2种DNA疫苗进行了对比,发现串联IL-2可以增强疫苗的免疫活性。李蕴玉等[15]将IL-2基因与EtMIC-2进行串联,构建了重组质粒pcDNA3.0-IL-2、pcDNA3.0-EtMIC-2、pcDNA3.0-IL-2-EtMIC-2并进行动物保护性试验,结果表明,IL-2可增强EtMIC-2基因的免疫原性和抗氧化能力。本试验将IL-2与pPIC9载体和球虫的SO7基因进行串联构建重组质粒pPIC9-IL-2-SO7,并成功转染毕赤酵母细胞GS115。毕赤酵母表达系统是一种外源蛋白真核表达系统,具有蛋白加工、折叠、翻译后修饰等优点,该表达系统调控产生过氧化物酶的启动子AOX1、AOX2等可以驱动外源基因在毕赤酵母中表达,其外源蛋白可以在信号肽α因子或蛋白自身信号肽的引导下跨膜运输分泌到胞外,为下一步的蛋白分离与纯化提供了便利[16]。E.tenella与毕赤酵母在种属分类具有一定的差异性(原虫与真菌),两者均属于单细胞真核生物,但是与细菌、植物等表达载体相比,其蛋白质翻译修饰系统更相近,适合于E.tenella相关蛋白的表达。
经Western blot检测表明重组蛋白SO7具有免疫原性。用纯化的重组蛋白SO7进行动物保护性试验,当免疫剂量为50,100 μg/只时,能显著抑制卵囊的繁殖,降低盲肠病变计分,提高鸡的相对增重率,ACI高于160,具有良好的抗球虫效果,为今后柔嫩艾美耳球虫核酸疫苗的发展奠定基础。关于重组蛋白SO7的抗球虫作用机理及其与抗球虫药物和活球虫疫苗的保护作用对比还有待进一步研究。
