槐角黄酮通过调控miR-188-5p/PRMT5基因表达对肺癌细胞增殖、迁移和侵袭的影响及其机制
2020-11-02侯从岭雷小婷芦晓帆周超锋李彬
侯从岭 雷小婷 芦晓帆 周超锋 李彬
(1河南省中医院肺病科,河南 郑州 450002;2河南中医药大学第二临床医学院肺病科;3河南省中医院肿瘤科)
肺癌发病率和死亡率在世界各地均在上升,肺癌的发生与吸烟饮酒密切相关〔1〕。大多肺癌患者确诊时已是晚期,癌症已发生转移,脑转移患者平均生存时间仅1~2个月,放化疗等方法预后较差,患者负担重〔2〕。中药因可延长患者生存期并改善患者生存质量,已成为肺癌治疗的重要组成部分〔3〕。槐角黄酮可能通过上调Bcl-2相关X蛋白(Bax)、激活半胱氨酸天冬氨酸蛋白酶(caspase)-3及下调B细胞淋巴瘤(Bcl)-2来抑制人胰腺癌细胞增殖,诱导细胞凋亡〔4〕。与癌旁组织相比,miR-188-5p在胃癌组织、前列腺癌和肝癌组织及细胞中表达下调,过表达miR-188-5p可显著抑制癌细胞的增殖和转移或促进癌细胞凋亡〔5~7〕。蛋白质精氨酸甲基转移酶(PRMT)5在肺癌、胃癌及肝癌组织和细胞中表达上调,干扰或下调PRMT5表达可抑制癌细胞的增殖〔8~10〕。但槐角黄酮对肺癌细胞的影响及机制尚不清楚。本研究以人肺癌细胞系A549为研究对象,研究槐角黄酮对肺癌的影响及miR-188-5p和PRMT5在此机制中的作用。
1 材料与方法
1.1材料 人肺癌细胞株A549购自美国模式菌种收集中心(ATCC);槐角购自中药批发市场;胰蛋白酶Trypsin、牛血清白蛋白(BSA)、二甲基亚砜(DMSO)和四氮唑蓝(MTT)购自Sigma-Aldrich公司;F-12K培养基和胎牛血清(FBS)购自Gibco公司;Transwell板购自美国Corning公司;双荧光素酶报告系统(Dual-Luciferase Reporter Assay System)购自美国Promega公司;Lipofectamine 2000转染试剂、RNA提取试剂Trizol、 real-time PCR 试剂盒、反转录试剂盒(RT-PCR)购自美国Invitrogen公司;抗PRMT5抗体、细胞周期蛋白(Cyclin)D1抗体、p21抗体、抗基质金属蛋白酶(MMP)-2抗体、抗MMP-9抗体和抗GAPDH抗体购自Abcam;显微镜、发光仪、酶标仪及Real-time PCR仪购自美国Bio-Rad公司;流式细胞仪购自美国BD公司;二喹啉甲酸(BCA)蛋白检测试剂盒购自Thermo。
1.2方法
1.2.1细胞培养、药物处理 细胞培养:A549细胞培养于含10%FBS、1%青-链霉素的F-12K培养基中,置于湿度保持95%、37℃且5%CO2培养箱中培养。待细胞培养至对数生长期,消化传代。药物处理:槐角黄酮采用文献〔11〕方法提取和制备。将槐角黄酮以10 mg/ml的质量浓度溶解于F-12K培养基中,并稀释成1 mg/ml和5 mg/ml,过滤除菌。96孔板培养细胞至贴壁状态,将培养液吸出,换为含不同浓度槐角黄酮的培养液,培养48 h,收集细胞进行实验。
1.2.2细胞转染 将传代后的A549细胞用培养液稀释至(1~2)×106个细胞/ml,以2×105细胞/孔的密度接种于6孔板中,培养细胞至融合度为80%~90%,进行转染。取等体积用无血清培养液稀释的脂质体和各组片段及载体,混合均匀,室温孵育20 min,加入到培养好的A549细胞中,混匀,培养6 h,换为F-12K完全培养基,转染48 h,收集细胞,进行实验。
1.2.3Real-time PCR检测mRNA的表达 收集各组转染或(和)槐角黄酮处理的A549细胞,用Trizol试剂提取细胞总RNA,合成cDNA,合成的cDNA测定浓度和纯度后置于-80℃保存。取cDNA按照real-time PCR的说明书进行反应,反应程序为:94℃ 4 min;94℃ 30 s、58℃ 45 s、72℃ 30 s,35个循环;72℃10 min。引物如下:miR-188-5p:5′-CATCCCTTGCATGGTGGAGGG-3′;PRMT5上游:5′-CCTGTGGAGGTGAACACAGT-3′,下游:5′-AGAGGATGGGAAACCATGAG-3′。运用Bio-Rad PCR系统进行数据分析。
1.2.4MTT实验检测细胞增殖 收集转染或处理后的A549细胞,消化,稀释细胞,以2×103个/孔接种于96微孔板中,继续培养至24、48、72 h时进行MTT实验,每孔加入20 μl(5 mg/ml)MTT,培养4 h,弃培养上清,每孔再加入150 μl DMSO,室温混匀5 min,酶标仪测定490 nm 吸光度(A)值。
1.2.5Transwell实验检测细胞迁移和侵袭 迁移实验:将转染或处理后的各组A549细胞培养至对数生长期,收集细胞,加入含10 g/L BSA的无血清F-12K培养基,将细胞稀释为2×105个/ml。Transwell下层培养孔加入500 μl含10%FBS的F-12K培养基,Transwell上层小室中加入100 μl细胞,将上层小室放入下层培养孔中,培养48 h,取出用棉签拭去基质胶和上层小室的细胞,冰甲醛固定细胞,染色,显微镜计数。侵袭实验:以4℃无血清培养基1∶3比例稀释Matrigel,加入上层Transwell小室,烘干,以下步骤同迁移实验,上层加入100 μl稀释的细胞,下层加入500 μl含10%FBS的F-12K培养基,培养48 h,固定细胞,染色,计数。
1.2.6双荧光素酶报告实验 A549细胞转染48 h,收集细胞,Trypsin消化,计数,以1×104个细胞/孔接种于24孔板中,培养24~48 h,观察若细胞融合度达到80%~90%,进行转染,将构建的PRMT5的野生型(WT-PRMT5)和突变型(MUT-PRMT5)双荧光素酶报告载体分别共转染miR-NC或miR-188-5p,转染后培养48 h,收集细胞,加入裂解液室温裂解30 min,离心收集上清,加入荧光素酶底物,发光仪检测荧光素酶活性。以海肾荧光素酶活性为内参照,计算相对萤火虫荧光素酶活性。
1.2.7Western印迹实验 将转染48 h后的各组A549细胞用放射免疫沉淀试验(RIPA)裂解液裂解,超声破碎细胞,收集蛋白并检测浓度。将蛋白样本进行十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE),转膜,封闭1 h,分别加入一抗,抗PRMT5抗体(1∶1 000)、CyclinD1抗体(1∶500)、p21抗体(1∶800)、抗MMP-2抗体(1∶500)、抗MMP-9抗体(1∶800)和抗GAPDH抗体(1∶1 000),4℃孵育过夜。洗膜2次,然后加入稀释的二抗,室温孵育2 h。以GAPDH为内参照,分析蛋白水平。
1.3统计学处理 采用SPSS21.0软件进行t检验、方差分析。
2 结 果
2.1槐角黄酮对肺癌A549细胞增殖和迁移侵袭的影响 与对照组相比,槐角黄酮1 mg/ml组、5 mg/ml组和10 mg/ml组肺癌细胞A549的增殖蛋白CyclinD1表达明显下降,p21表达明显上升,侵袭相关蛋白MMP-2和MMP-9表达显著下降,细胞抑制率显著上升,迁移细胞数和侵袭细胞数均显著下降(均P<0.05),且呈现显著的剂量依赖趋势。见图1,图2,表1。

图1 Western印迹检测增殖、迁移侵袭相关蛋白表达

图2 槐角黄酮对肺癌A549细胞迁移、侵袭的影响(结晶柴染色,×200)

表1 槐角黄酮对肺癌A549细胞增殖和迁移侵袭的影响
2.2槐角黄酮对肺癌A549细胞中miR-188-5p和PRMT5表达的影响 与对照组相比,槐角黄酮1 mg/ml组、5 mg/ml组和10 mg/ml组A549细胞中miR-188-5p表达水平显著上升,PRMT5 mRNA和蛋白表达量显著下降(均P<0.05)。见表2,图3。
表2 槐角黄酮对肺癌A549细胞中miR-188-5p和PRMT5表达的影响

图3 Western印迹检测PRMT5蛋白表达
2.3miR-188-5p靶向调控PRMT5的表达 PRMT5的3′-UTR序列中含有与miR-188-5p互补的核苷酸序列,见图4A。与miR-NC组(1.00±0.09,n=6)相比,miR-188-5p组野生型WT-PRMT5的萤火虫荧光素酶相对活性显著下降(0.32±0.03,n=6,t=17.558,P=0.000);而突变型MUT-PRMT5的萤火虫荧光素酶相对活性无明显变化(miR-NC组0.98±0.08、miR-188-5p组0.99±0.09,n=6,P>0.05)。与miR-NC组(0.61±0.03,n=12)相比,miR-188-5p组的PRMT5蛋白表达量显著下降(0.23±0.03,n=12,P<0.05);与anti-miR-NC组(0.60±0.06,n=12)相比,anti-miR-188-5p组的PRMT5蛋白表达量显著上升(0.95±0.08,n=12,P<0.05),见图4B。
2.4miR-188-5p过表达可抑制肺癌A549细胞增殖和迁移侵袭 与miR-NC组相比,miR-188-5p组的miR-188-5p表达量显著升高,增殖蛋白CyclinD1表达下降,增殖抑制蛋白p21表达明显上升,侵袭相关蛋白MMP-2和MMP-9表达显著下降,细胞增殖抑制率显著上升,迁移细胞数和侵袭细胞数均显著下降(P<0.01,P<0.001)。见图5,表3。

A:PRMT5的3′UTR中含有与miR-188-5p互补的核苷酸序列;B:PRMT5蛋白表达

图5 Western印迹检测增殖、迁移侵袭相关蛋白表达

表3 miR-188-5p过表达对肺癌A549细胞增殖和迁移侵袭的影响
2.5抑制PRMT5表达可抑制肺癌A549细胞增殖和迁移侵袭 与si-NC组相比,si-PRMT5组的PRMT5蛋白表达显著下降,增殖蛋白CyclinD1表达显著下降,增殖抑制蛋白p21表达显著上升,侵袭相关蛋白MMP-2和MMP-9表达显著下降,细胞增殖抑制率显著上升,迁移细胞数和侵袭细胞数均显著下降(均P<0.001),见表4,图6。
表4 抑制PRMT5表达对肺癌A549细胞增殖和迁移侵袭的影响

图6 PRMT5和增殖、迁移侵袭相关蛋白表达
2.6抑制miR-188-5p表达逆转了槐角黄酮(10 mg/ml)对肺癌A549细胞增殖和迁移侵袭的作用 与对照组相比,槐角黄酮(10 mg/ml)组的A549细胞PRMT5蛋白表达量显著上升,增殖蛋白CyclinD1表达显著下降,增殖抑制蛋白p21表达显著上升,侵袭相关蛋白MMP-2和MMP-9表达显著下降,细胞增殖抑制率显著上升,迁移细胞数和侵袭细胞数均显著下降(均P<0.05);与槐角黄酮组+anti-miR-NC组相比,槐角黄酮组+anti-miR-188-5p组的A549细胞PRMT5蛋白表达量显著下降,增殖蛋白CyclinD1表达上升,增殖抑制蛋白p21表达降低,侵袭相关蛋白MMP-2和MMP-9表达显著上升,细胞增殖抑制率显著降低,迁移细胞数和侵袭细胞数均显著上升(均P<0.05)。见图7,表5。
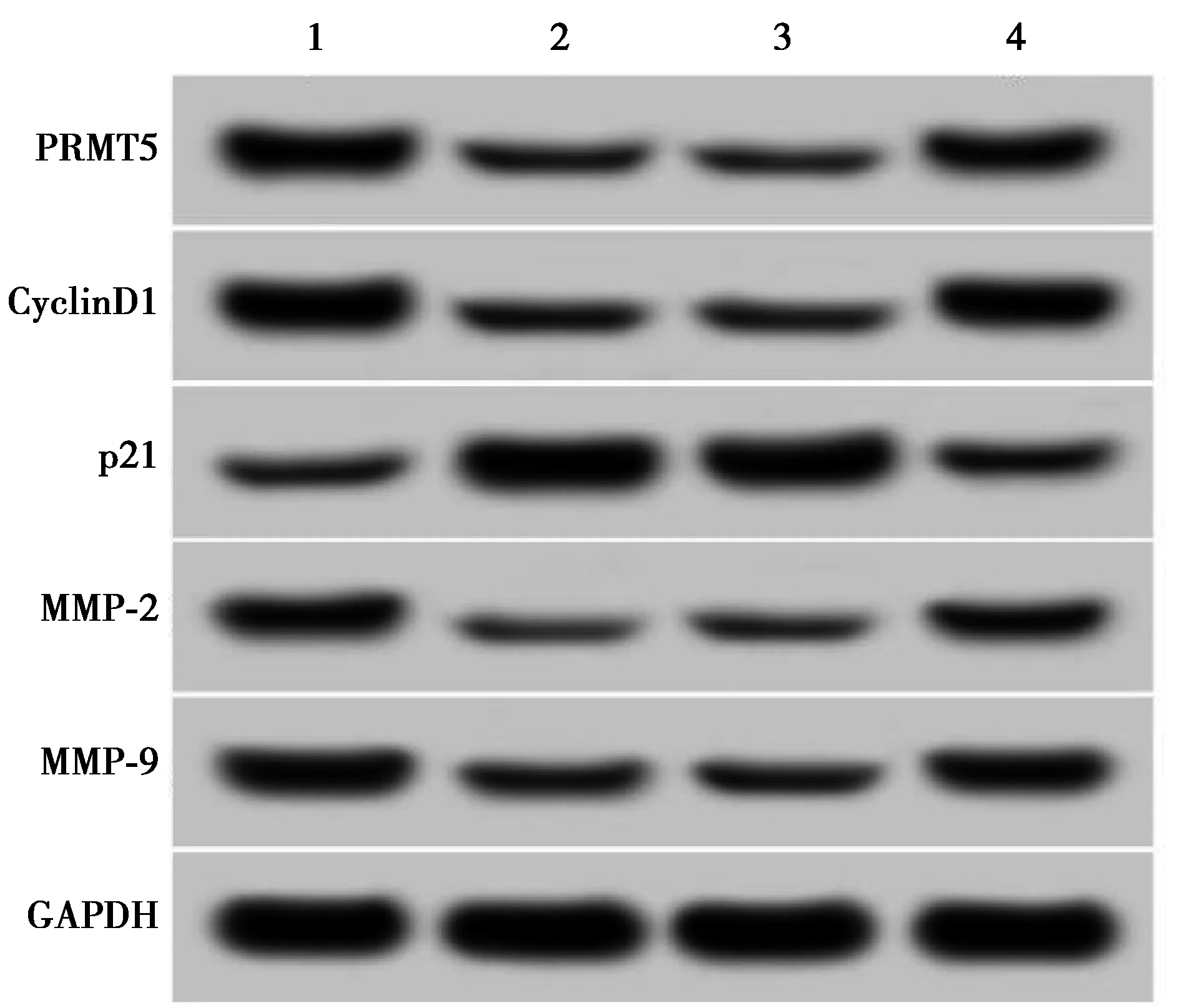
1~4:对照组、槐角黄酮组、槐角黄酮+anti-miR-NC组、槐角黄酮+anti-miR-188-5p组
表5 抑制miR-188-5p表达逆转了槐角黄酮对肺癌A549细胞增殖和迁移侵袭的作用
3 讨 论
肺癌是全世界癌症死亡的主要原因,75%的患者在确诊时已发展为晚期(Ⅲ/Ⅳ期),2014年英国国家统计局报告称,远处转移性肺癌(Ⅳ期)患者1年生存率仅为15%~19%〔12〕。尽管肺癌的诊断和治疗(手术、放疗、化疗和靶向治疗)有很大进步,但是晚期患者预后仍然很差〔11〕。中药在治疗肺癌中起着越来越重要的作用,通过提高患者免疫力,改善微循环,抑制肿瘤新生血管形成及促进细胞凋亡等,达到控制和抑制肿瘤的作用,改善放化疗的不良反应,提高患者生存质量〔13〕。但中药对肺癌的分子抑制机制,尚不完全清楚。
槐角是豆科植物槐(Sophora japonica L)的果实,具有凉血止血的功效,槐角富含异黄酮类化合物,对骨质疏松和癌症等具有较好的预防和治疗作用,其中的化合物包括染料木素、槐属苷、染料木苷、槐属双苷、刺芒柄花素、刺芒柄花苷、大豆苷等,其中的染料木素在合适的浓度对肺癌A549和胃腺癌BGC-823细胞抑制率分别可达82%和91%〔14〕。槐角转化产物染料木素和异樱黄素对乳腺癌细胞具有抑制增殖和促进凋亡的作用〔15〕,槐角黄酮对胰腺癌也有显著的抑制作用〔4〕。本研究发现,槐角黄酮可抑制肺癌细胞A549的侵袭、迁移和增殖,且呈显著的剂量依赖性,又一次验证了槐角黄酮的抗肿瘤作用。
miR-188能产生miR-188-5p和miR-188-3p两种miRNA,在多种肿瘤组织中表达异常,调控肿瘤的发生和发展。Zhang等〔16〕研究发现,miR-188-5p在转移性前列腺癌组织中表达下调,过表达miR-188-5p可通过抑制LAPTM4B抑制磷酸酰肌醇3激酶/蛋白激酶B(PI3K/AKT)信号通路,抑制前列腺癌细胞增殖和转移。Wang等〔17〕最新发现,在胃癌组织和细胞中,miR-188-5p表达异常升高,miR-188-5p通过上调癌基因婆罗双树转录因子(SALL)4,促进胃癌细胞增殖和迁移,同时抑制抑癌基因PTEN(第10号染色体同源丢失性磷酸酶-张力蛋白基因)表达。本研究发现,槐角黄酮可促进A549中miR-188-5p的表达,过表达miR-188-5p可以抑制A549细胞增殖、迁移和侵袭,与Zhang等〔16〕研究结果类似;抑制miR-188-5p可逆转槐角黄酮对A549细胞增殖、迁移和侵袭的抑制作用,说明miR-188-5p在槐角黄酮的抑癌机制中发挥作用。
此外,本研究通过Targetscan预测发现,PRMT5的3′-UTR序列中含有与miR-188-5p互补的核苷酸序列,预示miR-188-5p与PRMT5之间可能存在结合位点或者调控关系。PRMT5可能参与槐角黄酮对肺癌A549的抑制机制。PRMT5是蛋白-精氨酸甲基转移酶家族中的一员,它形成独特的同质寡聚体,主要位于细胞质,催化蛋白中精氨酸残基对称二甲基化的形成,与维持干细胞干性有关,肿瘤干细胞的出现有助于耐药性的形成〔18〕。PRMT5通过调控乳腺癌干细胞来决定其对化疗药物的敏感性〔19〕,同时其也是胶质母细胞瘤的药物治疗靶点〔20〕。Sheng等〔21〕发现,PRMT5在肺癌和前列腺癌中通过调控编码生长和抗生长因子的一组特定基因的表达,包括受体酪氨酸激酶和抗增殖蛋白,进而调控癌细胞的增殖。Jing等〔22〕总结大量研究结果发现,PRMT5作为潜在的致癌基因,通过组蛋白和其他蛋白甲基化参与细胞增殖、分化、侵袭、迁移和凋亡,对肺癌细胞的生长和转移至关重要,其高表达通常提示肺癌临床预后较差。Jing等〔23〕研究表明,PRMT5 在人肺癌组织中经常高表达,其通过组蛋白H4R3的对称二甲基化抑制miR-99家族的转录,从而增加FGFR3的表达,进而激活Erk1/2和Akt,导致肺癌细胞生长和转移,敲除PRMT5可通过阻断miR-99家族的组织学修饰抑制肺癌转移。本研究结果与上述结果一致。说明槐角黄酮通过抑制PRMT5表达抑制肺癌。
此外,本研究证实在肺癌A549细胞中miR-188-5p和PRMT5之间具有调控关系,也说明槐角黄酮通过miR-188-5p靶向抑制PRMT5进而抑制肺癌。
