取食和交配对苹小吉丁飞行能力的影响
2020-10-15马志龙卡德艳卡德尔阿地力沙塔尔张元明
马志龙, 彭 彬, 卡德艳·卡德尔, 阿地力·沙塔尔,*, 张元明
(1.新疆农业大学林学与园艺学院, 乌鲁木齐 830052;2.中国科学院新疆生态与地理研究所, 乌鲁木齐 830011)
苹小吉丁Agrilusmali国外分布于俄罗斯和东亚等国,在我国主要分布在北部以及西北地区,主要危害苹果、海棠、桃等经济果树(崔晓宁等, 2015)。苹小吉丁传入伊犁地区后在山区野果林及平原栽培果园迅速扩散蔓延,对新疆苹果产业健康发展造成了极大威胁。苹小吉丁以幼虫钻入韧皮部和木质部进行取食危害,化蛹于木质部,成虫羽化后在树皮上咬一“D”形羽化孔后爬出枝条。危害苹果树初期可造成大量的枝条干枯,大暴发时常面临毁园的威胁(李孟楼和张正青, 2017; 阎雄飞等, 2017);成虫喜欢在晴天或气温稍高的时候绕树冠飞行,阴雨天或气温较低时常静伏于枝叶上(崔晓宁等, 2015)。在饲喂苹果叶片的条件下苹小吉丁雄虫平均存活9~14 d,雌虫平均存活15~22 d(李孟楼等, 2017)。在饥饿条件下,苹小吉丁雄虫在第3天开始出现死亡高峰,雌虫于第4天出现死亡高峰(李孟楼等, 2017)。苹小吉丁通过苗木调运传入新疆伊犁州(季英等, 2004),人为传播是主要传播因素,但也会依靠自身的飞行在林间扩散蔓延。昆虫自身的传播主要与其飞行能力有关(Tayloretal., 2010)。飞行能力是昆虫迁飞、扩散等行为的重要参考指标。飞行距离、飞行时间和飞行速度是判断昆虫飞行能力强弱的重要参数。这些参数一般依靠昆虫飞行磨进行测量,这为评价昆虫的飞行能力提供了参考(王世磊, 2011)。利用飞行磨对吉丁类昆虫飞行能力测定已有相关研究,如对杨十斑吉丁Melanophilapicta和花曲柳窄吉丁Agrilusplanipennis不同性别、交配状态和取食情况条件下成虫的飞行能力的研究已有报道(Tayloretal., 2010; 孙栋等, 2014; 王辉, 2014)。
苹小吉丁传入伊犁地区后在山区野果林及平原栽培果园迅速扩散蔓延,对新疆苹果产业健康发展造成了极大威胁。但关于苹小吉丁飞行扩散能力如何,现在还尚不清楚。因此,本研究以采自新疆伊犁地区的苹小吉丁为材料,对不同性别和不同日龄苹小吉丁成虫的飞行能力进行了测定,同时探究了影响飞行能力的重要因素,研究结果可为苹小吉丁潜扩散区域预测和防控工作提供参考。
1 材料与方法
1.1 试虫来源
试虫采自新疆巩留县综合农场栽培果园及库尔德宁镇、七十二团八连和哈拉布拉乡萨哈野果林分布区,于6月中下旬开始将带有苹小吉丁老熟幼虫、蛹及未羽化成虫的苹果枝条从基部截取,装入带有透气窗的塑料罐中。塑料罐为透明PET材质(罐底宽15.5 cm,高24.0 cm,口径11.2 cm),罐子底部铺一层厚3~5 cm的湿润细沙,将苹果枝条插入细沙以保持水分。于罐体四面各截取一个直径约10 cm的圆形窗口,窗口粘贴100目的纱网作为通风透气口,同时防止羽化成虫逃逸。将羽化罐置于通风透光处进行自然羽化,羽化成虫按雌雄分别收集装入塑料饲养瓶中,瓶口贴明羽化时间。
1.2 测试仪器
通过使用SUN-FL型智能昆虫飞行信息系统即飞行磨(北京鹏程电子科技中心生产)对苹小吉丁虫潜在飞行能力进行测定,该系统由飞行磨采集控制主机、飞行磨盘和PC机软件组成。
1.3 飞行能力测定
1.3.1飞行能力吊飞测试方法:在测试前将飞行磨吊臂取下,将试虫(不麻醉)前胸背板涂抹少许502胶或粘虫胶,轻轻固定在飞行磨吊臂一端的吊针上保持试虫与吊杆垂直,调整吊臂平衡,并使试虫飞行方向与飞行轨迹相切。试虫固定后用手轻触或吹气,查看试虫是否能够正常展翅飞行,如能正常飞行,将飞行磨吊臂安装回飞行轴,然后打开飞行测试软件,设置参数,让试虫绕中轴进行飞行。每次测试24 h,测试计算试虫的飞行速度、飞行持续时间和飞行距离(丁吉同等, 2014; 丁吉同, 2015)。测试环境温湿度分别由三菱重工恒温恒湿控制机组(型号:MDC265A/MAUC265)控制在28±1℃和35%~45% RH。光强为600~700 lx,光周期为14L∶10D。
1.3.2苹小吉丁不同日龄成虫飞行能力测定:按1.3.1节的吊飞测试方法,分别取羽化后1, 2, 3, 4, 5, 6, 7, 8, 9, 11, 13和15 d共12个日龄的成虫进行测试,每个日龄雌雄成虫分别测试15头。
1.3.3取食对苹小吉丁飞行能力的影响测定:用采集于野苹果林的新鲜野苹果叶片饲喂试虫至3日龄。苹果叶片叶柄用沾水的脱脂棉进行包裹防止水分迅速蒸发以保持叶片的新鲜程度,叶片每天更换一次。同时另一组成虫不饲喂任何食物作为对照。按1.3.1节的吊飞测试方法,对两组试虫进行吊飞测试,分析取食情况对苹小吉丁飞行能力的影响。取食组和对照组分别测定30头。
1.3.4交配对苹小吉丁飞行能力的影响测定:将苹小吉丁雌雄成虫分为两组,第1组进行两两配对,时间为1~2 d。第2组雌雄分开不进行交配。按1.3.1节的吊飞测试方法,对两组试虫于6日龄时进行测飞,测飞时间为24 h。每个处理测试15头,分析交配情况对苹小吉丁飞行能力的影响。
1.4 数据分析
数据使用Excel 2010进行初步处理,运用SPSS 17.0对不同日龄苹小吉丁飞行能力各指标进行单因素方差分析(one-way ANOVA),同时对数据进行Duncan氏多重比较测定在5%水平下的差异显著性。对不同交配和取食处理条件下苹小吉丁的飞行能力差异进行T检验。
2 结果
2.1 不同日龄苹小吉丁成虫飞行能力
对不同日龄苹小吉丁成虫的飞行能力进行了测定,由表1和表2可见:苹小吉丁雌虫各飞行指标均表现出随着日龄的增加先上升后逐渐下降的趋势,24 h内平均飞行距离、平均飞行时间、最长飞行距离、最长飞行时间及最大飞行速度的峰值均出现于11日龄,分别为0.387 km, 0.419 h, 0.4165 km, 0.4582 h和2.4639 km/h,而平均飞行速度的变化趋势稍有不同,最大值出现在4日龄,为0.975 km/h。苹小吉丁1日龄雌成虫的各飞行指标均较低,平均飞行距离和平均飞行时间分别为0.016 km和0.017 h,显著低于7日龄以上的成虫(P<0.05)。苹小吉丁不同日龄雄成虫各飞行指标与雌成虫的变化趋势较为一致,均随着日龄的增加先上升后降低,但稍有差异。平均飞行距离、平均飞行时间、最长飞行距离及最大飞行速度的峰值同样出现于11日龄,分别达到0.301 km, 0.370 h, 0.3559 km和1.8561 km/h。平均飞行速度的最大值也出现在4日龄,达到0.856 km/h。而最长飞行时间与雌虫不同,最大值出现在9日龄,为0.4873 h。1日龄雄成虫各飞行指标依然最低,平均飞行距离、平均飞行时间和平均飞行速度分别仅为0.019 km, 0.010 h和0.595 km/h,且平均飞行距离和平均飞行时间显著低于7日龄以上的雄成虫(P<0.05)。

表1 苹小吉丁不同日龄成虫平均飞行距离、时间和速度
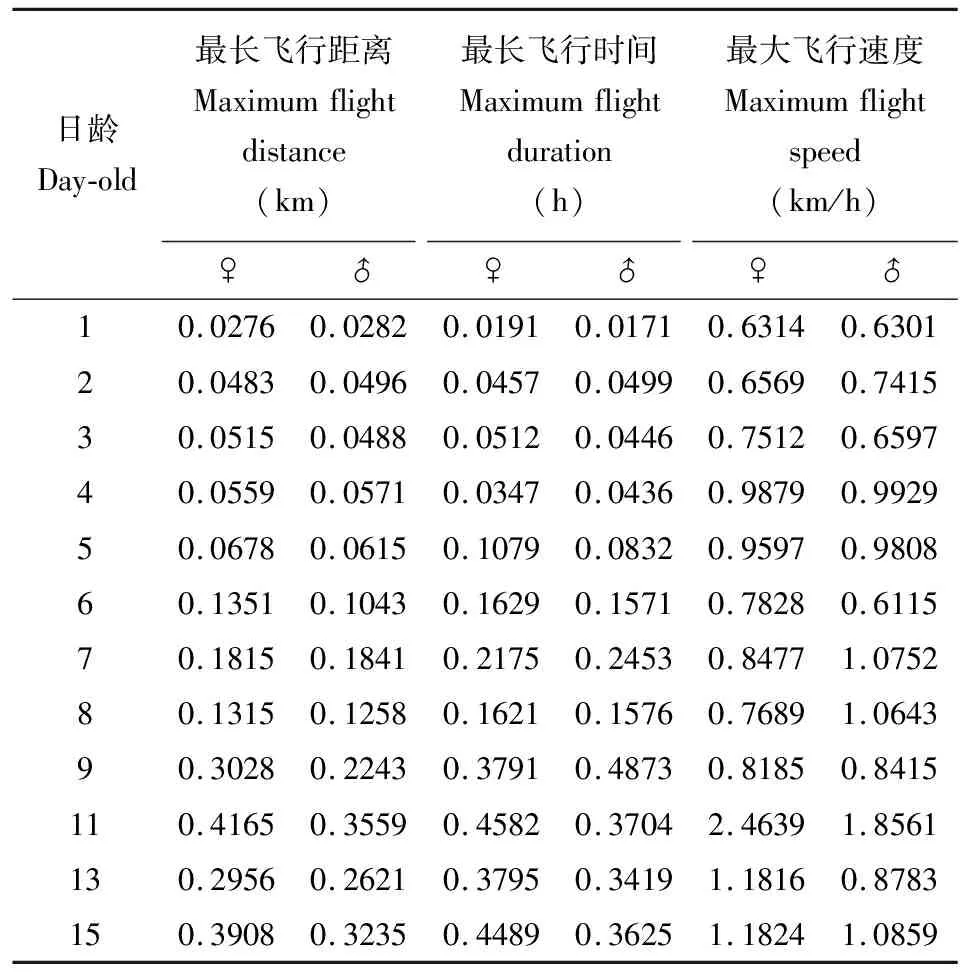
表2 苹小吉丁不同日龄成虫最大飞行距离、速度和最长飞行时间
2.2 取食对苹小吉丁飞行能力的影响
饲喂野苹果叶片的苹小吉丁雌雄成虫的平均飞行距离和平均飞行时间均显著高于未取食成虫。经过取食的苹小吉丁雌雄成虫的平均飞行距离分别为0.047和0.044 m,而未取食的雌雄成虫分别为0.016和0.013 m,差异显著(t=24.020,t=17.759;df=28;P<0.05);经过取食的雌雄成虫平均飞行时间依次为0.048和0.042 h,而未取食的雌雄成虫仅分别为0.013和0.012 h,且差异显著(t=62.749,t=48.684;df=28;P<0.05);从平均飞行速度来看,经过取食的雌雄成虫平均飞行速度分别为0.703和0.620 km/h,未取食的雌雄成虫平均飞行速度分别为0.590和0.576 km/h,经过取食的雌雄试虫平均飞行速度更快,且经T检验发现两者之间差异也达到显著水平(t=5.694,t=3.188;df=28;P<0.05)(图1)。
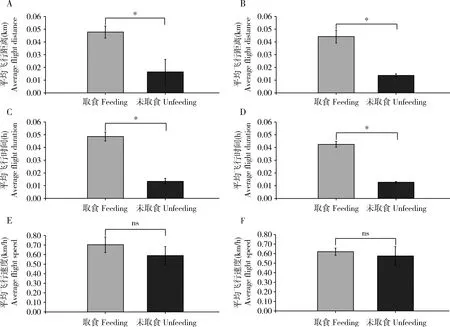
图1 取食对苹小吉丁3日龄成虫平均飞行距离、时间和速度的影响
2.3 交配对苹小吉丁飞行能力的影响
从图2中可以看出,苹小吉丁6日龄雌成虫交配后的平均飞行距离、平均飞行时间和平均飞行速度分别为0.145 km, 0.169 h和0.802 km/h,未交配的分别为0.124 km, 0.158 h和0.710 km/h,交配后雌成虫的各飞行指标稍高于未交配雌虫,且差异显著(t=5.048,t=4.660,t=4.420;df=28;P>0.05);苹小吉丁6日龄雄成虫交配后的平均飞行距离、平均飞行时间和平均飞行速度分别为0.087 km, 0.135 h和0.569 km/h,未交配的分别为0.090 km, 0.142 h和0.586 km/h,与雌虫的飞行能力不同,交配后的雄虫平均飞行距离和时间略低于未交配雄虫,但T检验结果表明两者之间只有平均飞行速度差异显著,其他两个指标间差异不显著(t=-0.897,t=-1.609,t=-2.589;df=28;P=0.377,P=0.119,P=0.015)。

图2 交配对苹小吉丁6日龄成虫平均飞行距离、时间和速度的影响
3 讨论
昆虫飞行能力的评价一般是在室内条件下通过使用相应的吊飞仪器即飞行磨对其飞行时间、速度和距离等指标进行测定,而昆虫的飞行能力通常会由于日龄、性别、交配状态等条件的不同而产生差异(武怀恒, 2006; 吴孔明等, 2006; 郭江龙等, 2016)。本研究结果表明,苹小吉丁的飞行能力随着日龄的增加表现出先上升后逐渐降低的趋势,初羽化的成虫飞行能力较弱,11日龄成虫的飞行能力达到最强(表1和2)。日龄对苹小吉丁成虫的飞行能力影响作用显著,可能是由于昆虫的飞行需要糖类和脂类等营养物质为其提供能量支撑,而不同日龄所积累的糖元和甘油三脂等物质量的不同导致了飞行能力的差异,这在甜菜夜蛾Spodopteraexigua和二点委夜蛾Athetislepigone飞行过程对能源物质需求中已有报道(江幸福等, 2002; 郑作涛, 2014)。再如,初羽化的绿盲蝽Lyguslucorum成虫需要不断地补充营养来促进飞行肌的发育(Luetal., 2007)。但是,鳞翅目昆虫舞毒蛾Lymantriadispar的飞行能力则表现不同,舞毒蛾成虫期不需要进行补充营养,初羽化的成虫飞行能力较强,随着日龄的增加飞行能力逐渐减弱(郭虹, 2012; 杨帆, 2013)。
成虫在饥饿状态下或长期营养不足时飞行能力会显著降低(张蕾等, 2006; 郭文超等, 2012),甚至幼虫期的营养状况也会对成虫期的飞行能力产生一定的影响(王凤英等, 2010; 李霞等, 2013)。本研究吊飞试验发现,取食对苹小吉丁的飞行能力产生了显著的影响,饲喂苹果叶片的雌雄试虫飞行能力均高于未饲喂试虫(图1),提示苹小吉丁成虫羽化后是否能够及时进行取食将决定其能否通过自身进行较远距离的飞行扩散。迁飞行为是昆虫选择生存环境的一种生态适应性表现,与其内在的生理条件如交配、产卵等具有一定的关联,且与雌虫的性状发育更为密切(武怀恒, 2006; 黄学飞等, 2010)。本研究还发现,苹小吉丁已交配雌虫的飞行能力更强,平均飞行距离、平均飞行时间和平均飞行速度均高于未交配雌成虫;已交配雄虫平均飞行距离、平均飞行时间和平均飞行速度均略低于未交配雄成虫(图2)。与日龄和取食对苹小吉丁飞行能力的影响作用相比,交配情况对雄虫的影响作用稍弱(图2),说明苹小吉丁的飞行能力主要受到日龄和取食的调控。由于苹果小吉丁发育时间不整齐,所以供试虫源获取较为困难,样本数量相对较少。在条件允许情况下可以继续补充虫源进行测试,以获取更精准的飞行数据。
昆虫的体型大小也会引起飞行能力的差异,花曲柳窄吉丁和绿盲蝽雌成虫的飞行能力要强于雄虫,就是因为雌虫的体型普遍大于雄虫(Luetal., 2007; 王辉等, 2015)。与同为吉丁科的杨十斑吉丁(孙栋等, 2014)和花曲柳窄吉丁(王辉, 2014)的飞行能力相比,苹小吉丁的飞行能力相对稍弱,可能是由于其体型较小、体重较轻,为飞行所提供的能量物质相对较少所致。因此,幼虫期饲喂食物的种类及不同取食量对苹小吉丁成虫体型大小、营养物质积累和飞行能力影响方面还值得深入研究。
