大兴安岭次生林蒙古栎种群的空间格局
2020-10-09韩雪敏张秋良边玉明任建华
韩雪敏, 张秋良, 边玉明, 任建华
(内蒙古农业大学林学院,内蒙古 呼和浩特 010018)
种群是同一生物个体在一定的空间和时间内的总和,年龄结构和空间分布格局是种群在特定时间和空间中的真实模式表现,是衡量种群动态的重要指标[1-4].年龄结构是指个体在种群不同年龄阶段中所占的比例,年龄动态及时间序列预测可以较为直观地反映不同生长阶段种群的存活、死亡趋势以及未来发展状态.种群的空间分布格局是种群与外界环境相互适应的结果,对其进行分析有利于掌握种群动态变化规律及其生态过程[5-6].
近年来,种群结构及其动态变化逐渐成为植物生态学的研究热点[7].岳永杰等[8]对北京松山自然保护区蒙古栎林的分布格局进行研究,结果表明该林分的空间分布格局为聚集分布,但林木聚集程度和聚集规模较低;赵清格[9]对内蒙古赛罕乌拉国家级自然保护区的蒙古栎进行研究,发现蒙古栎种群表现为增长型;张会儒等[10]对蒙古栎过伐林的空间结构进行研究,结果表明蒙古栎的单种聚集明显,胸径比和树高比分化较为严重;陈科屹等[11]对吉林省汪清林业局塔子沟林场蒙古栎种群空间分布格局进行研究,发现蒙古栎种群的分布格局随尺度增大由聚集分布转向随机分布;李艳丽等[12]对北京市怀柔区渤海镇三岔村蒙古栎——糠椴天然混交林进行研究,结果表明蒙古栎种群的径级结构呈增长型分布,并随尺度增加趋向于随机分布.
蒙古栎(Quercusmongolica)是壳斗科栎属,为阳性树种,喜光.蒙古栎是内蒙古大兴安岭地区的主要硬阔叶树种之一,以中、幼龄林为主,多生长在干旱瘠薄的土壤环境中,有较强的适应能力.蒙古栎次生林是对原始林的不合理采伐或火烧之后形成的,粗放经营造成蒙古栎材质次、干形差、长势弱,导致林分生产力低下、结构不合理.目前,对内蒙古大兴安岭地区蒙古栎次生林的研究甚少,因此本文以内蒙古大兴安岭蒙古栎次生林为研究对象,用径级代替龄级的方法研究蒙古栎种群年龄结构的变化趋势;利用点格局分析方法研究蒙古栎种群的空间格局,以期为今后该地区蒙古栎次生林的科学经营提供重要的参考依据.
1 研究区概况
研究区位于内蒙古大兴安岭吉文林业局境内,地理位置为122°37′21′′—123°44′13′′E,49°45′34′′—50°41′45′′N,最高海拔1 220 m,最低海拔391 m.该地区属于寒带大陆性季风气候,昼夜温差大,四季变化显著.年平均气温-2.6~-0.8 ℃,自西向东递增.无霜期90 d,年平均降水量451.2~499.3 mm.土壤种类较多,有棕色针叶林土、潜育棕色针叶林土、灰色针叶林土、沼泽土、草甸土等.以兴安落叶松为优势的针叶林占主导地位,其次是白桦纯林和兴安落叶松、白桦混交林,以及柞树和黑桦林等.
2 研究方法
2.1 样地设置及调查
在内蒙古大兴安岭吉文镇吉库林场115林班,设置一块90 m×40 m的固定样地.由表1可知,样地内树种组成单一,蒙古栎为主要树种,黑桦(Betuladahurica)为伴生树种.对样地内胸径≥5.0 cm的蒙古栎进行每木检尺,用多样性测度仪测量每株树的具体坐标(x,y).

表1 样地信息汇总
2.2 种群年龄结构及动态
采用径级代替龄级的方法[13],根据测得的胸径数据,以5.0 cm为一个径级[14],将蒙古栎划分为7个等级,推测蒙古栎林年龄结构的发展趋势.Ⅰ级:5.0~10.0 cm.Ⅱ级:10.0~15.0 cm.Ⅲ级:15.0~20.0 cm.Ⅳ级:20.0~25.0 cm.Ⅴ级:25.0~30.0 cm.Ⅵ级:30.0~35.0 cm.Ⅶ级:>35.0 cm.根据径级结构,将蒙古栎种群分为小树(Ⅰ~Ⅱ级)、中树(Ⅲ~Ⅳ级)和大树(Ⅴ~Ⅶ级)[12]3个年龄段.
根据陈晓德[15]种群动态数量化方法,对种群动态进行定量描述:
(1)
(2)
式中,Vn表示种群从n到n+1级的个体数量变化;Sn、Sn+1表示第n级、第n+1级种群的个体数;Vpi表示整个种群结构的数量变化动态指数;k表示种群径级数量.式(2)仅适用于在没有外部环境干扰的情况下种群结构动态的比较.存在外部干扰时,式(2)应修正为:
(3)
2.3 种群静态生命表的编制及时间序列分析
根据蒙古栎种群不同龄级的个体数,采用径级代替龄级的方法统计各龄级株数,编制种群静态生命表,进而分析其动态变化.采用一次移动平均法[16]来模拟预测蒙古栎种群的龄级结构.
(4)
式中,n为需要预测的年限;Xk为当前k龄级的种群大小;t为龄级.t分别取种群经历2、4、6 年的时间段,对蒙古栎种群的未来发展趋势进行模拟预测.
2.4 种群空间分布格局及空间关联性
以RipleyK函数为基础,分析处于不同径级的蒙古栎个体在不同尺度范围内的分布格局.K函数的公式[17]表示为:
(5)
式中,A为样地面积;n为样地内物种个体总数;r为空间尺度;Wij为以点i为圆心、uij为半径的圆落在面积A中的弧长与整个圆周的比值,可校正边界效应引起的误差;uij为点i到点j的距离;I为指示函数.为了更直观地解释实际的空间格局,常用Ripley L函数代替K函数,其公式为:
(6)
应用RipleyK双变量K12(r)函数对两个物种的空间关联性进行点格局分析.本文通过2个不同物种的点格局分析得出同一物种不同年龄段间的空间关系[17],表示为:
(7)
式中,n1、n2为2个不同年龄段蒙古栎植株总个体数;i和j为2个不同年龄段的植株;A、Wij和Ir(uij)含义同RipleyK函数式.同理,对K12进行平方根转换得L12函数,公式为:
(8)
利用生态学软件ADE-4对蒙古栎种群的空间格局进行分析.通过1 000次Monte-Carlo拟合得到99%的置信区间.L(r)的最大值和最小值,分别为上下包迹线的坐标值.以r为横坐标,以上下包迹线为纵坐标绘图.L(r)值若在上下包迹线以内,符合随机分布;若在上包迹线之上,为聚集分布;若在下包迹线之下,则为均匀分布.同理,仍用Monte-Carlo拟合计算方法检验不同年龄段间是否相关,L12(r)值若在上包迹线之上,为正相关;若在下包迹线之下,为负相关;若在上下包迹线以内,则不相关.
3 结果与分析
3.1 年龄结构特征
由图1可知,胸径≥5.0 cm的蒙古栎个体共有178株,小树所占比重较大.经统计,Ⅰ、Ⅱ龄级(小树)个体共82株,占总个体数的46%;Ⅲ、Ⅳ龄级(中树)个体57株,占总个体数的32%;Ⅴ、Ⅵ、Ⅶ龄级(大树)个体共39株,占总个体数的22%.说明该地区蒙古栎低龄级个体补充充足,大树个体相对较少,新生个体数会随着年龄增长而减少,年龄结构呈增长型.随着时间的推移,蒙古栎林内Ⅱ龄级、Ⅲ龄级和Ⅳ龄级的个体数量逐渐增加,种内竞争不断增强.
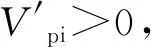

表2 蒙古栎种群龄级结构的动态变化指数
3.2 种群静态生命表及存活曲线
根据调查数据编制了大兴安岭蒙古栎种群的静态生命表(表3).由表3可知,蒙古栎种群的死亡率随径级的增长呈先下降后上升的趋势,说明蒙古栎种群的生长受到自身生物学特性以及种内竞争的影响.其中,第Ⅱ级和第Ⅲ级的死亡数及死亡率出现了小于0的情况,表明该阶段内蒙古栎种群处于一个稳定增长的时期.从第Ⅳ级开始,随着龄级的增加,蒙古栎种群的死亡率呈增长趋势.蒙古栎种群的生命期望随龄级的增加呈递减趋势,蒙古栎种内竞争加强进而导致生命期望逐渐降低.

表3 蒙古栎种群静态生命表1)
蒙古栎种群的存活状况可通过种群存活曲线来反映.其中,DeeveyⅠ型(凸曲线)表示该种群的全部个体都可以达到生理寿命;DeeveyⅡ型(直线)表示该种群各年龄时期的死亡率是相同的;DeeveyⅢ型(凹曲线)表示该种群幼期的死亡率高,之后死亡率低并且呈稳定趋势[18].
由图2A可知,蒙古栎种群的存活曲线接近DeeveyⅠ型(凸曲线).其中,Ⅰ龄级到Ⅱ龄级的存活率迅速下降;Ⅱ龄级到Ⅳ龄级存活率又有所上升;从Ⅴ龄级开始蒙古栎年龄接近其生理年龄,存活率下降明显.说明在低龄级时期蒙古栎种群的死亡率较低,种群数量大幅度增长;到达一定时间后,存活数量减少,种群开始衰退,最终趋于稳定.由图2B~2C可知,蒙古栎种群死亡率和消失率的变化曲线基本一致.从图2可以看出,随着群落演替的不断进行,蒙古栎个体逐渐趋于成熟,对环境的要求也越来越严格,导致蒙古栎个体种内竞争加强,使得蒙古栎种群的死亡率逐渐升高.由于Ⅱ、Ⅲ龄级内幼苗数量较少,导致该龄级的死亡率出现负值,对蒙古栎种群的更新会产生一定的影响.
3.3 种群动态变化时间序列预测
从表4可以看出,2年后Ⅲ龄级的蒙古栎个体的数量保持不变,Ⅳ龄级的数量将会减少,而其余龄级的数量均增加;4及6年后各龄级的数量都会逐渐增加.可以推断出,蒙古栎种群在未来将呈现出稳定增长的趋势.

表4 蒙古栎种群动态变化的时间序列预测
3.4 种群点格局分析
蒙古栎种群个体空间分布如图3所示,横轴表示样地长90 m,纵轴表示样地宽40 m,用实测坐标值(x、y)来表示蒙古栎个体的空间位置.可以初步看出,蒙古栎种群存在丛生和萌生的现象,这与实地调查的结果一致.
图4为样地中蒙古栎种群点格局分析的结果.在进行分析时,将r的间隔设置为1 m,最大值为20 m.从图4可以看出,在0~20 m间蒙古栎种群呈明显的聚集分布,聚集强度在0~6 m间线性增强,在13~20 m间线性减小;当r=9 m时,聚集强度和聚集规模最大,分别为4.43和254.34 m2.
从图5可知,样地的林分面积为0.36 hm2,蒙古栎种群内小树的个体数为82,密度为228株·hm-2;中树的个体数为57,密度为159株·hm-2;大树的个体数为39,密度为109株·hm-2.株数密度大小表现为小树>中树>大树.
从图6可知,在0~20 m内小树呈聚集分布,聚集强度在0~6 m间线性增强,当r=6 m时,聚集强度和聚集规模最大,分别为3.85和113.04 m2;在0~20 m内中树呈聚集分布,聚集强度在0~7 m间线性增强,当r=7 m时,聚集强度和聚集规模最大,分别为4.51和153.86 m2;在0~1 m和18~20 m间大树呈随机分布,在1~18 m间呈聚集分布,当r=8 m时,聚集强度和聚集规模最大,分别为5.43和200.96 m2.
3.5 空间关联性分析
由图7可知,蒙古栎种群小树与中树、小树与大树以及中树与大树之间均不相关.
4 讨论与结论
本研究发现,蒙古栎种群的年龄结构呈相对稳定的状态,种群存活曲线接近于DeeveyⅠ型,说明在低龄级时期蒙古栎种群的死亡率较低,种群增长较快;当达到一定的时间以后,种群进入衰退阶段,并随着时间推移最终趋于稳定.从蒙古栎种群的静态生命表和存活曲线可以看出,目前种群内的幼苗数量较少,难以满足种群的更新要求.随着种群的不断发展,如果幼苗得不到及时补充,种群将会慢慢走向衰退.经调查发现,蒙古栎林内萌生株数较多,而实生株数较少,出现这一现象的原因可能是蒙古栎成熟后掉落的种子被动物啃食,使实生苗的存活受到影响,进而导致蒙古栎种群出现了天然更新不良的现象.
本研究结果表明,蒙古栎种群呈聚集分布,这是由于受到小径阶和中径阶林木的影响,这与陈科屹等[11]对蒙古栎种群空间分布格局的研究结果相近,与赵清格[9]对赛罕乌拉自然保护区蒙古栎种群的研究结论也十分相似.这种现象与蒙古栎的繁殖特性和种子扩散方式有关.由于蒙古栎的根系具有很强的萌蘖能力,种子质量较大,扩散能力差,成熟后会集中散落在母树附近,这些原因都会导致蒙古栎呈现聚集分布的空间格局.
在进行点格局分析时,取样尺度越大越好,可以较为全面客观地反映种群在不同尺度上的空间分布格局,使分析结果更加真实准确.本研究由于受到蒙古栎分布区域以及地形地势条件等因素的限制,取样面积仅为90 m×40 m,点格局分析得出的结论也仅仅局限于20 m的尺度范围内,因而研究结果具有一定的局限性.
通过对大兴安岭蒙古栎种群年龄结构及空间分布格局的研究发现,蒙古栎虽然为增长型种群,但幼苗数量较少,种群的径级分布呈左偏单峰山状,不符合理想的异龄林的反“J”型分布曲线;且蒙古栎种群呈聚集分布,不同龄级间各自相互独立,小树与中树、小树与大树及中树与大树之间都没有相关性.
