斑石鲷和条石鲷线粒体基因组密码子使用分析
2020-09-23张志勇张志伟张曹进陈淑吟贾超峰曹广勇
孟 乾,张志勇,张志伟,张曹进,陈淑吟,祝 斐,贾超峰,曹广勇
( 江苏省海洋水产研究所,江苏省海水鱼类遗传育种重点实验室,江苏 南通 226007 )
在生物体内的蛋白质翻译过程中,编码同一种氨基酸的不同密码子称为同义密码子,而不同基因在蛋白质编码过程中会倾向于使用一些特定的同义密码子,叫作同义密码子的使用偏性,不同物种及不同功能基因的密码子使用偏性均存在差异[1-5]。线粒体DNA(mtDNA)具有进化速率快、核苷酸替代率高及母性遗传等特点,已成为鱼类进化、鉴定及种质遗传等研究的重要遗传分子标记,mtDNA的密码子偏性研究有助于推进鱼类的鉴定、发育及遗传育种等研究。
斑石鲷(Oplegnathuspunctatus)和条石鲷(O.fasciatus)均属鲈形目、石鲷科、石鲷属,在我国沿海均有分布,且具有美观、营养丰富、生长快及抗逆性强等特点,具有较高的观赏与经济价值,其海水养殖产业具有发展前景[6-9]。目前,国内外对条石鲷和斑石鲷的研究主要集中在选择育种、人工繁殖技术、病害及分类等方面[7-15],而在mtDNA水平研究较少。孙鹏等[16]以mtCOX1和Cytb片段探讨了条石鲷野生种群多样性。此外,斑石鲷和条石鲷间已实现人工杂交,二者在基于mtCOX1、Cytb基因和D-loop序列片段的遗传差异比较中显示亲缘关系较近[17-18]。笔者以斑石鲷和条石鲷mtDNA编码蛋白基因序列为研究对象进行密码子偏好性分析,并与斑马鱼(Daniorerio)、金鼓鱼(Scatophagusargus)、黑鲷(Acanthopagrusschlegelii)和真鲷(Pagrosomusmajor)mtDNA编码蛋白质的基因序列进行比对和密码子偏好性聚类分析,探讨石鲷科与其他物种间的亲缘关系,以期为探讨两者间的近缘杂交育种、种质资源等研究提供理论依据。
1 材料与方法
1.1 序列材料
本研究所用斑石鲷、条石鲷、斑马鱼、金鼓鱼、黑鲷和真鲷的线粒体基因组基因序列均来源于美国国立生物技术信息中心GenBank数据库(表1)。

表1 斑石鲷、条石鲷及其他物种的登录号
1.2 密码子分析指标和方法
相对密码子使用度,是指对于某一特定的密码子在编码对应氨基酸的同义密码子间的相对概率,可直观地反映密码子使用的偏好性。当某一密码子相对密码子使用度=1,代表该密码子的使用没有偏好性;相对密码子使用度>1时,表明该密码子为使用相对较多的密码子,反之亦然。第i个氨基酸的第j个密码子的相对密码子使用度(RSCUij)的计算公式[3]如下:
式中,xij是编码第i个氨基酸的第j个密码子的出现次数,ni是编码第i个氨基酸的同义密码子的数量(值为1~6)。
用CodonW对斑石鲷和条石鲷mtDNA中编码蛋白基因序列分别进行分析,得出每条序列中各密码子的相对密码子使用度以及密码子的第3位碱基中A、U、G、C的含量在第3位碱基总量中所占的比率(分别为A3s、U3s、G3s、C3s);用Microsoft Excel 2006统计分析数据。
1.3 因子相关性及聚类分析
将各密码子含量以标准得分存为变量作为横轴,相对密码子使用度值以1为原点作为纵轴,以Microsoft Excel 2006作散点图;每个物种的mtDNA编码蛋白质的基因作为一个对象,将密码子的相对密码子使用度作为变量,以偏好性密码子的相对密码子使用度(大于1)对基因的密码子使用偏好性进行聚类分析;去除斑石鲷等6个物种的mtDNA的ATP8、ND3和ND4L(序列长度较短),每个基因(ND1、ND2、COX1、COX2、ATP6、COX3、ND6、ND4、ND5、Cytb)各自看成一类并以Mega 5.1进行聚类分析。
2 结果与分析
2.1 斑石鲷与条石鲷mtDNA编码蛋白质基因的密码子偏好性
斑石鲷和条石鲷的mtDNA编码蛋白基因中的C3s分别为31.21%和35.86%,均大于A3s、U3s、G3s(表2),可见斑石鲷和条石鲷的mtDNA编码蛋白基因偏爱使用以C碱基结尾密码子,避免使用以G结尾密码子。
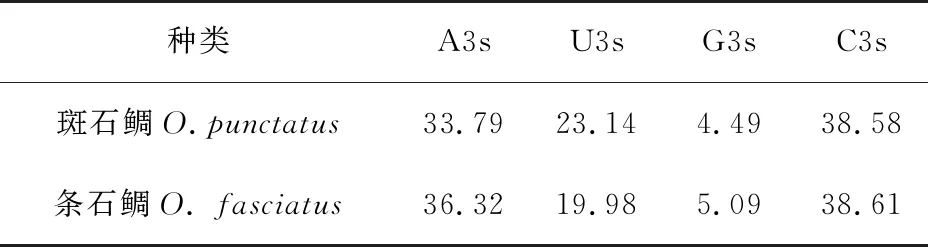
表2 斑石鲷和条石鲷mtDNA编码蛋白质基因密码子的A3s、U3s、G3s、C3s %
斑石鲷以UUC、ACC等30个密码子的相对密码子使用度均大于1(表3),为斑石鲷mtDNA编码蛋白基因的偏好性密码子,条石鲷比斑石鲷的mtDNA编码蛋白基因的偏好性密码子少2个(CCA和GUC),其中尤其偏爱使用以C结尾的密码子(斑石鲷除AUC外,条石鲷除AUC和GUC外,其余的均为偏好性密码子),其次偏爱以A结尾的密码子(特别是CGA,其相对密码子使用度分别为2.96和3.32);尽量避免使用以G结尾的密码子(除斑石鲷CGG的相对密码子使用度值为0.86,其余相对密码子使用度均小于0.5)。

表3 斑石鲷和条石鲷线粒体编码蛋白基因序列偏好密码子的相对密码子使用度

(续表3)

(续表3)
依据密码子编码氨基酸原理进行计算,斑石鲷和条石鲷线粒体中的各种氨基酸含量基本一致,含量较高的4种氨基酸依次为亮氨酸>异亮氨酸>丙氨酸>苏氨酸,其中除异亮氨酸偏好密码子为AUU,其余3种氨基酸的偏好密码子也均以AC结尾(图1)。
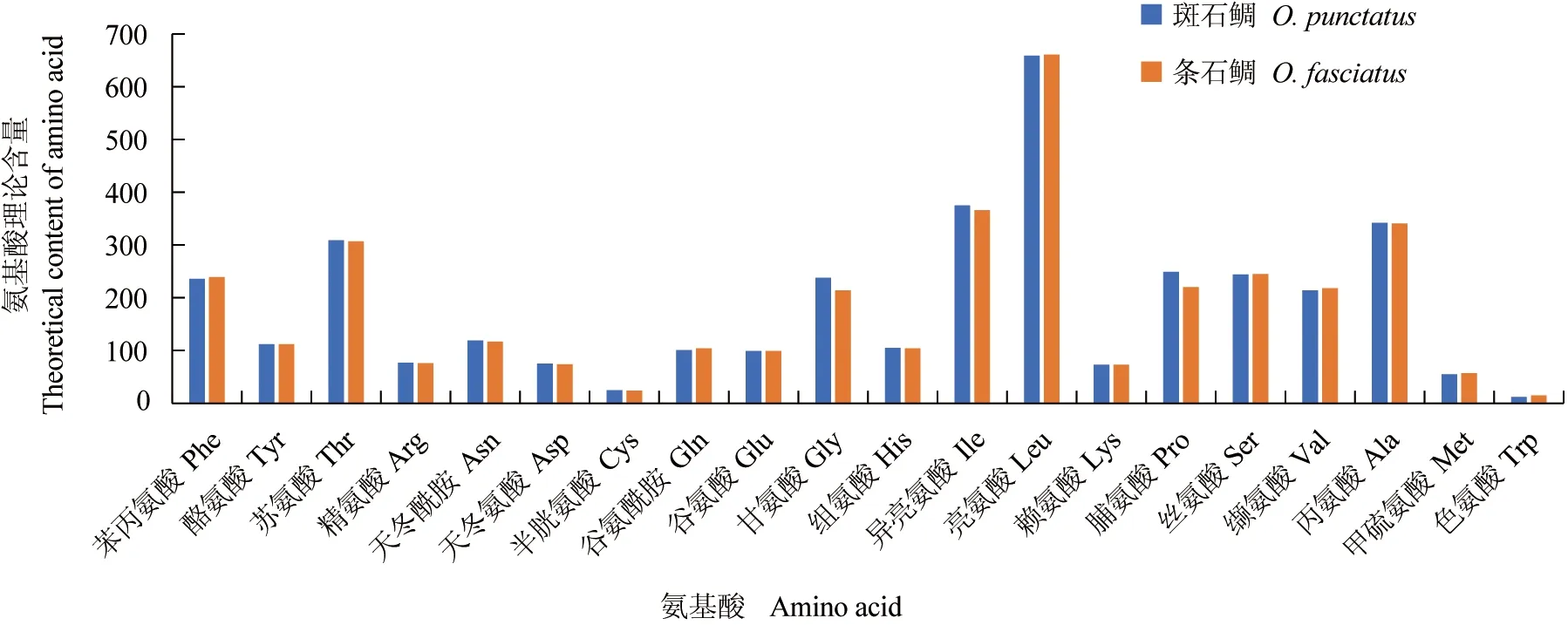
图1 斑石鲷和条石鲷各种氨基酸含量
2.2 因子相关性分析
斑石鲷与条石鲷线粒体编码蛋白序列因子相关性分析x、y两轴的分析结果表明,该2种鱼线粒体编码蛋白序列中,AC结尾的密码子与GU结尾的密码子明显分开,距离中心较近的AC结尾与GU结尾密码子比例相近,但A、C含量相对较高(图2),表明该2种鱼较偏好使用AC结尾的密码子。

图2 斑石鲷和条石鲷线粒体编码蛋白序列的因子相关性分析
2.3 偏好性密码子及基因序列的聚类分析
在对基因密码子使用概率进行聚类分析的过程中,将每个物种的mtDNA编码蛋白质的基因作为一个对象,将密码子的相对密码子使用度作为变量。除去相对密码子使用度始终为1的编码色氨酸和甲硫氨酸的密码子UGG和AUG,再除去3个不编码氨基酸的终止密码子,取余下的59个中偏好性密码子的相对密码子使用度(大于1)对基因的密码子使用偏好性进行聚类分析。
聚类结果见图3,大致分为2类:斑石鲷与条石鲷首先聚类,然后与金鼓鱼聚为一类;金鼓鱼和黑鲷首先聚为一类,说明它们的密码子使用的偏好性相似。此外,斑石鲷与金鼓鱼幼鱼阶段形态相似:金鼓鱼体黄褐色,体扁、吻短,散布许多黑圆斑,腹部银白;斑石鲷体色较淡呈灰褐色,各鳍暗色,头部、体侧、胸鳍及各奇鳍基部密布许多黑斑(图4)。

图4 斑石鲷和金鼓鱼外形
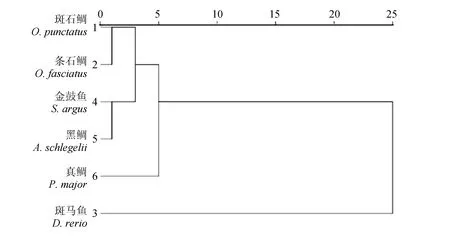
图3 斑石鲷、条石鲷与其他物种的mtDNA编码蛋白质的基因密码子偏好性聚类
将所研究的6个物种的每个mtDNA编码蛋白质的每个基因(ND1、ND2、COX1、COX2、ATP6、COX3、ND6、ND4、ND5、Cytb)各自看成一类,对每个基因序列进行聚类分析。其结果为斑石鲷和条石鲷均首先聚为一类,它们的距离相对较近;斑石鲷、条石鲷2种鱼在ATP6、ND2、COX1、ND5中首先聚类后,其次与金鼓鱼聚为一类,这与密码子偏好性聚类结果一致(图5)。

图5 最大似然法对斑石鲷、条石鲷及其他物种mtDNA编码蛋白基因序列构建的进化树
3 讨 论
3.1 密码子偏好性分析
基因是生物遗传信息的载体,蛋白质是发挥生物功能的主要分子。密码子是联系两者的元件,记录了长期进化过程中物种内在的功能适应性变化,其使用情况必然具有重要的生物学意义。斑石鲷和条石鲷mtDNA编码蛋白的序列偏好使用以AC结尾的密码子,最高使用频率的密码子是CGA(相对密码子使用度分别为2.96和3.32),其次是AAA(1.97和1.89) ,但总体而言以C结尾比A结尾密码子偏好性略高,这与分析金鱼(Carassiusauratus)[19]转录组偏好使用GC结尾密码子分析结果存在差异,说明物种密码子使用在进化过程中存在保守性,相近物种的密码子使用偏好性相似,反之则密码子的使用存在差异。斑石鲷和条石鲷mtDNA以G结尾密码子避免使用,且与其线粒体COX1、Cyt b和D-loop 3种片段上组成分布一致[18]。以密码子编码氨基酸的原理计算,理论上亮氨酸的氨基酸含量最高,且含量较高的亮氨酸(CUA、CUC、CUU)、异亮氨酸 (AUU)、苏氨酸(ACC、ACA)及丙氨酸(GCC、GCA)4种氨基酸均偏爱相同的密码子,表明不同氨基酸组成也一定程度影响密码子使用。
3.2 聚类分析
宋乔乔等[20]对牛亚科动物线粒体基因密码子偏好性进行聚类分析,发现基于密码子偏好性差异可以揭示物种间的亲缘关系,可用于物种的分类研究。不同物种的密码子偏好性差异与其亲缘关系也密切相关,密码子偏好性差异小,则亲缘关系近;反之,亲缘关系远[21]。Duan等[22]比较了14种鲌亚科鱼类密码子偏好性的差异,发现属间物种的密码子偏好性是具有物种特异性,主要是受到进化过程中的突变与自然选择的影响。基因突变具有低频性,故近缘物种密码子在使用偏好性上总体相似,又有轻微差别。在本研究中,密码子偏好性的聚类分析和基因序列的进化分析结果显示,密码子偏好性的聚类分析中斑石鲷和条石鲷聚为一类;基因序列的进化分析结果也显示,黑鲷与真鲷在ATP6、ND2、Cytb、ND1、ND4、ND5、ND6聚为一类,表明斑石鲷与条石鲷、黑鲷与真鲷亲缘关系较近,这与传统分类学一致。斑石鲷、条石鲷与斑马鱼在基因序列(除ND4)及偏好密码子聚类中均遗传距离最远,说明石鲷科与斑马鱼在进化过程中亲缘关系较远。斑石鲷、条石鲷2种鱼在密码子偏好性聚类中首先聚为一类后,其次与金鼓鱼聚为一类;基因序列的分析结果也显示,在ATP6、ND2、COX1、ND5上存在相同结论,且金鼓鱼和斑石鲷在幼鱼阶段外观相似,表明石鲷科与金鼓鱼亲缘关系较近,与传统分类学存在一定差异。密码子偏好聚类中,黑鲷与金鼓鱼先聚为一类,后与真鲷聚为一类,体现了金鼓鱼较真鲷与黑鲷亲缘关系更近,这与发育进化树及传统分类学[23]存在较大分歧。Christianson[24]认为,基于密码子的偏好性进行聚类,其结果与真实的系统发育关系不一定完全一致,这一指标在较小的分类单元里比较可靠,但是在分歧较大的分类单元里,这一方法主要还是作为传统系统树构建方法的补充和参考。
3.3 研究意义
不同物种密码子偏好性差异,产生原因可能是各物种生存环境不同,承受压力不同,导致突变和选择在物种之间程度有差异[25-26],此外,基因序列长度[27-28]、编码区GC含量[29]、翻译效率[30]和编码蛋白质的结构与功能[31]等均可引起密码子使用偏性,至于这些因素具体如何影响同义密码子偏好性仍有待进一步研究。同时有研究表明,密码子的使用偏性与基因的表达强弱有关,高效表达的基因所使用的密码子同低表达的基因相比较,它们具有明显不同的基因使用频率,高效表达的基因在密码子使用上比非高效基因更有偏向性趋势,它们通常使用一套偏好性同义密码子,同时发生偏好性使用的密码子还对应着含量最丰富的tRNA,提高翻译效率[31-33]。通过优化与改造密码子,可促进翻译效率及提高生物中外源基因的表达,如斑石鲷偏好密码子的基因作为外源基因,可以不经密码子优化,在条石鲷中获得高效表达;若依据斑石鲷的密码子偏好性信息进行密码子改造,可提高以斑石鲷偏好密码子的外源基因在其他模式生物中的表达效率。对于今后石鲷科鱼类转基因过程中对构建合适的转基因表达系统具有重要的指导意义,从而提高特定蛋白的表达量,增强转基因育种的效果。
同时,研究密码子偏好性还有很多的意义。研究密码子使用偏好性有利于改善靶基因在受体表达系统中的表达效果,通过对密码子进行改造优化已经成为提高外源基因在宿主细胞中的表达效率最有效且成功的途径之一;通过对密码子偏好性形成的主要因素探究,对于进一步了解生物基因或基因组特征、分子进化压力及生态适应等均具有重要启示;还有许多其他方面的意义,如鉴定基因的表达水平高低,判断是否发生基因重组,发现新基因等[34-36]。
4 结 论
本研究利用分析密码子偏好性及聚类分析等生物信息学方法,在mtDNA水平上对斑石鲷和条石鲷编码蛋白序列的密码子使用特征进行分析,明确了这2种鱼的mtDNA密码子偏好性,并比较与其他鱼类密码子使用偏好性差异。本研究结果可为探讨斑石鲷和条石鲷新基因预测、功能基因组学与其他水生动物在进化中亲缘关系等研究提供理论基础。
