对虾选择育种研究进展*
2020-08-26李旭鹏孟宪红陈宝龙曹家旺曹宝祥
孔 杰,栾 生,谭 建,隋 娟,罗 坤,李旭鹏,代 平,孟宪红,卢 霞,陈宝龙,曹家旺,曹宝祥
(1.中国水产科学研究院黄海水产研究所,山东 青岛 266071;2. 青岛海洋科学与技术试点国家实验室 海洋渔业科学与食物产出过程功能实验室,山东 青岛 266237)
选择育种是水产遗传改良工作中最为基本和常用的技术手段。其基本原理是假定生长速度、饲料转化效率和繁殖力等重要经济性状是由多个微效基因决定,然后利用数量遗传学方法构建核心育种群体并进行遗传评估,依据既定选择强度留取性能优良的个体,并在限定的近交水平下设计配种方案,持续多代定向选择,将分散在多个祖先个体中的优良基因富集到少数个体中,不断提高育种目标性状的遗传增益,最终达到育成新品种(系)的目的[1]。
人工选择在对虾育种工作中得到了广泛应用。利用群体选择方法,凡纳滨对虾(Litopenaeusvannamei)核心育种群进行多代群体选择,产量和IHHNV抗性均大幅度改良[2]。但群体选择难以剔除环境效应、非加性效应等影响因素,选种准确性较差,遗传进展缓慢;且该技术难以有效控制育种群体的近交水平,育种项目的可持续性差。1970年代,挪威育种学家以规模化家系为基础,以最佳线性无偏预测法(Best linear unbiased prediction, BLUP)为核心建立了多性状遗传评估技术[3],随后在凡纳滨对虾等选择育种项目中利用该技术也取得了较好的遗传进展。
国内自2004年开始,通过引进、消化吸收和自主创新,建立了基于规模化家系的“水产动物多性状复合育种技术”[4]。当前,该技术已在凡纳滨对虾、中国对虾(Fenneropenaeuschinensis)、日本对虾(Marsupenaeusjaponicus)等多个养殖虾类的选育研究中应用,成功培育出中国对虾“黄海5号”、罗氏沼虾(Macrobrachiumrosenbergii)“南太湖2号”、凡纳滨对虾“壬海1号”等多个新品种。
对虾在中国及世界水产养殖业中占有重要的地位。当前主要的养殖种类包括凡纳滨对虾、斑节对虾(Penaeusmonodon)、日本对虾和中国对虾等。根据FAO统计,2017年世界养殖产量超过520万t。其中凡纳滨对虾养殖产量超过445万t,是单一产值最高的水产养殖种类之一。对虾生长速度快、肉质鲜美,饲料、种虾、苗种、动保和加工产业链条完备且成熟,广大养殖户对优质苗种的需求非常迫切,是水产种业的一个典型代表种。对虾繁殖力强且性成熟周期短,可快速构建核心群和扩繁群,开展遗传改良工作具有选择强度高、遗传进展大、易于控种等优势。然而,由于核心群和扩繁群迭代速度快,少量亲本可贡献大部分后代,性状易产生近交衰退,高效的选种和配种方法等对育种项目遗传进展和可持续性的影响非常大。以规模化家系为基础的多性状综合选育技术,是支撑当前对虾种业的关键技术。本文综述了对虾生长、抗性、饲料转化效率、繁殖力等经济性状的测试与遗传评估方法,并对重要经济性状的遗传参数、基因型与环境互作效应和选择反应等进行了系统分析,包括近些年发展起来的基因组选择,间接遗传效应分析等最新结果,以期为精准、高效和可持续对虾选育提供参考。
1 选择、测试与评估方法
1.1 选择方法
1.1.1 群体选择 群体选择又称个体选择,是指基于候选个体的表型性状的一种选择策略,如生长、形状、颜色等[5]。最初的水产动物育种计划经常采用群体选择方法。对委内瑞拉凡纳滨对虾核心育种群进行11个世代群体选择后,产量增加了74%,由IHHNV(Infectious hypodermal and hematopoietic necrosis virus)导致的畸形率降低了96%[6]。还有研究表明,由于水产动物的繁殖力高,群体选择中少数个体可能对特定群体贡献了更多的后代,迅速导致高近亲繁殖率,降低了生长等重要性状的适应能力[7]。优化交配设计方案或采用分子系谱检测可以改善群体选择的结果。例如,每一世代至少选择50对亲本,每对亲本的后代数量限制在30~50,那么近交率可以控制在每代不超过1%[7]。
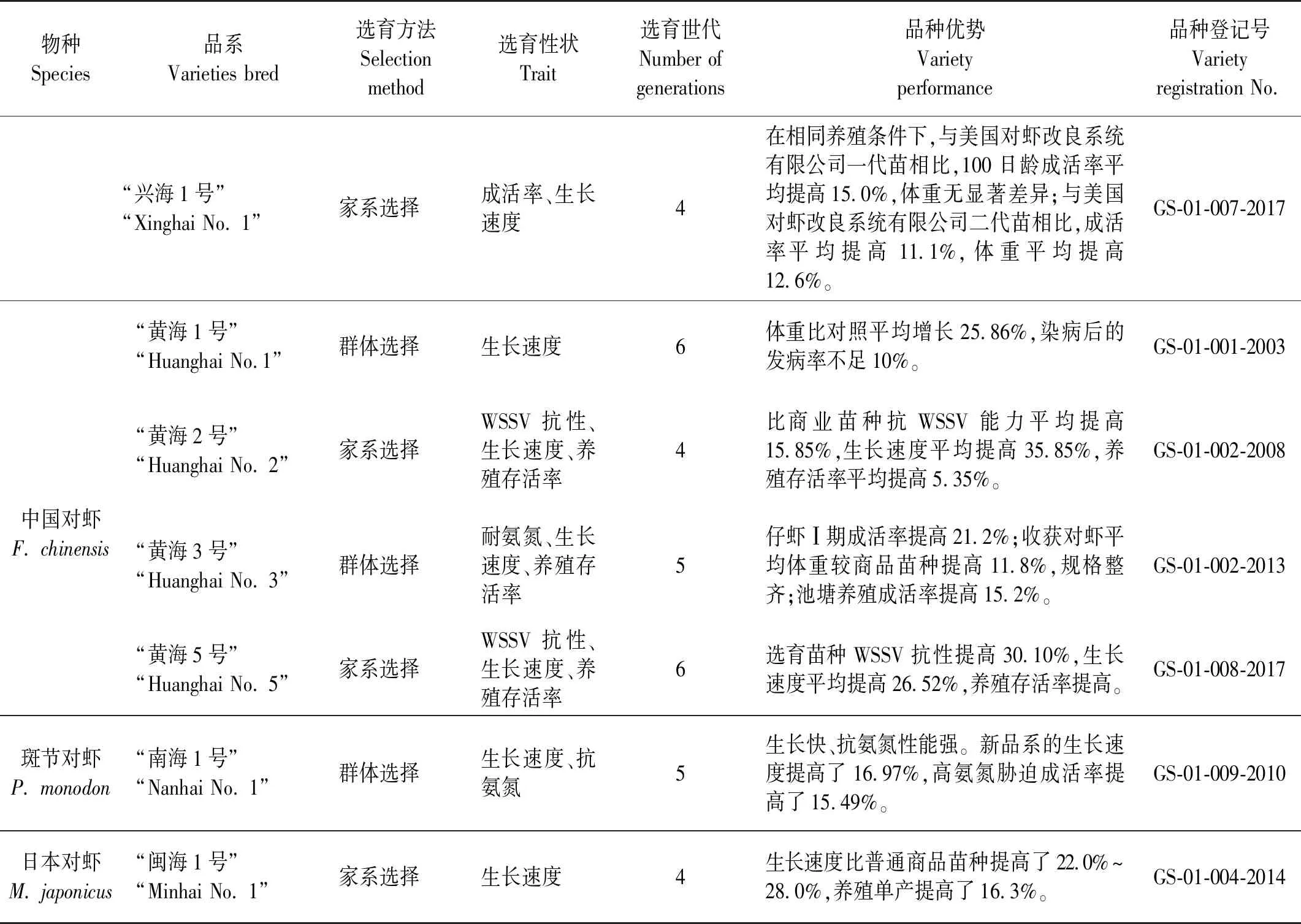
在中国对虾新品种的培育中,群体选择仍是一种重要的选择育种方法。利用该方法获得了中国第一个海水养殖动物新品种中国对虾“黄海1号”。此后,群体选择单独使用或与其他选择方法结合培育出了凡纳滨对虾“中科1号”、“中兴1号”、中国对虾“黄海3号”、斑节对虾“南海1号”等多个对虾新品种(见表1)。
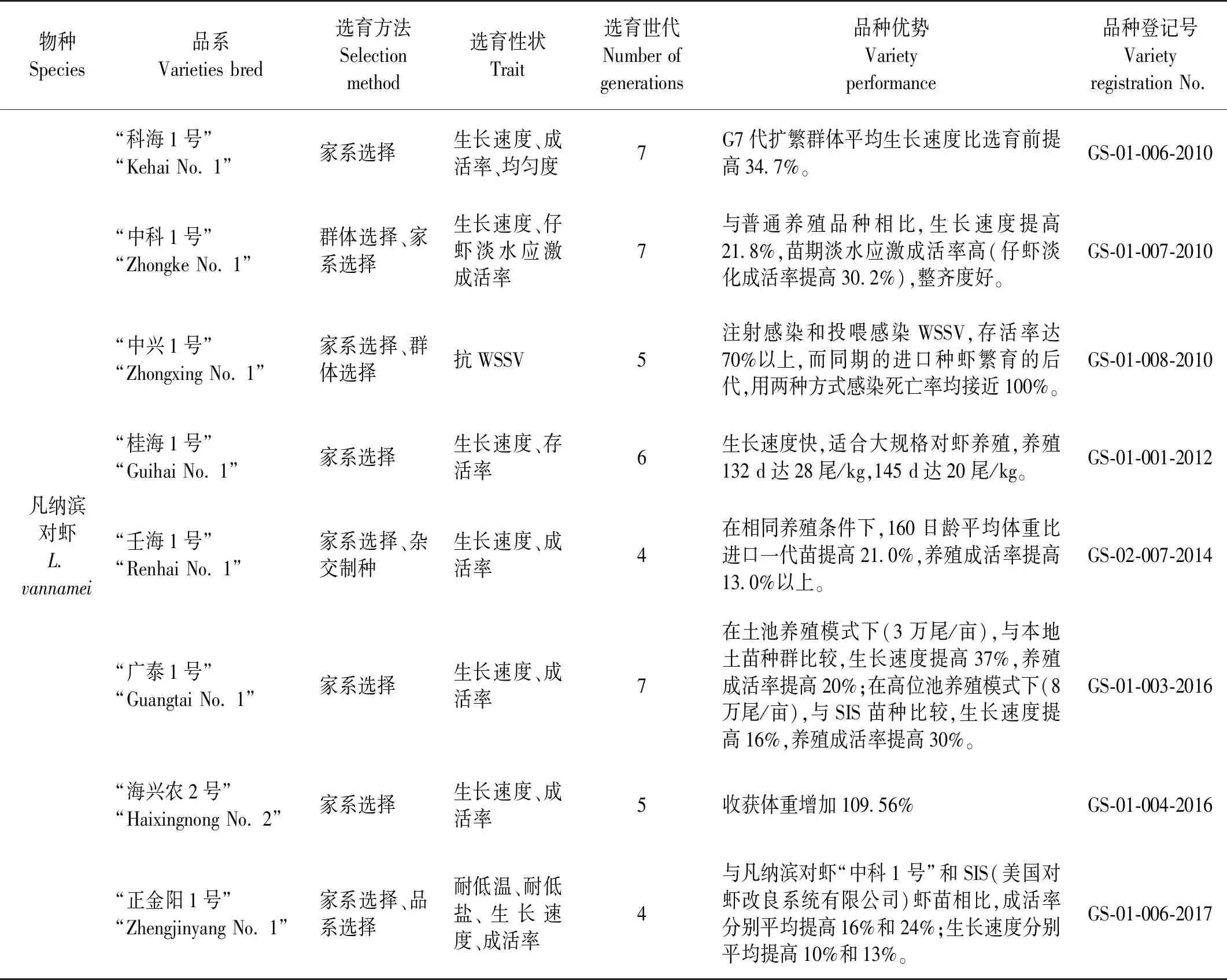
表1 中国自主培育的对虾新品种优势性状
续表1
1.1.2 规模化家系选择 规模化建立家系是开展以BLUP为核心的多性状复合育种的前提。对家系和个体标记识别,利用个体本身、同胞、祖先和后代等系谱和测定信息,通过约束极大似然法(Restricted maximum likelihood, REML)和BLUP法进行遗传评定,依据综合选择指数在家系和个体水平上选种和配种,可实现多个性状的复合选择,解决近亲交配及由此导致的种质退化问题[8]。
以家系为基础的选择方法,主要有三种:家系内选择、家系间选择和综合选择。(1)家系内选择是指对家系内优良个体的选择。该方法中多数家系都留下后代。但只能选择个体水平上的可测量性状,如生长、颜色等性状,改良性状的数量有限。(2)家系间选择是指对育种群体的优良家系的选择。依据家系的平均值,平均值高的家系入选,低的淘汰。通过同胞测试,选择一些宰杀性状,如抗性、肉质、出肉率等;群体性状,如饵料利用率等;不连续性状,如存活率、性成熟年龄等[5]。对于遗传力较低的性状,家系选择比群体选择更加有效[9]。(3)综合选择通常是对家系及家系内个体的双重选择。可利用BLUP方法,除个体本身、同胞表型数据外,结合祖先和后代等系谱信息估计个体育种值,根据不同性状的权重确定选择指数,在家系和个体水平上进行选择。“水产动物多性状复合育种技术”即为在此基础上建立的育种技术。
家系选择是目前对虾育种中最广泛使用的选择方法。在育种实践中根据实际需要可单独使用或与其他选择方法结合使用。在中国15个对虾新品种中,有12个应用了家系选择方法,具体包括凡纳滨对虾“科海1号”、 “中科1号”、 “中兴1号”、 “桂海1号”、 “壬海1号”、 “广泰1号”、 “海兴农2号”、 “正金阳1号”、 兴海1号;中国对虾“黄海2号”、 “黄海5号”;日本对虾“闽海1号”等(见表1)。
1.2 目标性状与测试方法
1.2.1 生长性状 生长性状直接影响着对虾养殖的经济效益,是对虾育种主要的目标性状。针对对虾生长性能的测试和评价,主要以体重、体长性状为主,测试的方法主要分为标记测试法和平行测试法。
标记测试法:测量家系个体的体长、体重并进行物理标记(每个家系不少于30尾),放入共同的环境中进行养殖测试,测试结束后可以获得家系或群体体长、体重性状的特定增重率、绝对增重率、变异系数等数据,并为生长性状遗传参数的估计提供了基础数据。目前,采用的物理标记主要为可视嵌入性荧光标记(Visible implant elastomer, VIE),将VIE标记注射到对虾第五、六腹节的不同部位,结合注射的不同颜色和不同部位可以在共同养殖环境条件下实现大规模家系或群体的性状测试。研究表明,对体长4 cm的中国对虾幼虾(体重1.8 g)注射VIE标记,不会影响对虾的正常生长和存活,且VIE标记的保持率很高,达99%以上[10]。VIE标记已广泛应用于对虾生长性能的测试[11-13]。
平行测试法:对于无法采用物理标记进行个体区分的对虾种类或对虾个体,通过设置多个平行组的方式进行养殖测试。通常采用在同一个养殖池内设置网格或布置网箱,保证养殖密度、投喂、换水等条件的一致性,尽可能消除养殖测试环境的差异,以确保测试数据的准确性。孙苗苗等[14]在水泥池中用围网隔成大小相同的水体开展了斑节对虾生长测试,分析了13个斑节对虾家系的特定生长率,为育种工作的开展提供了参考数据。
1.2.2 抗病性状 世界上几个主要的对虾养殖品种,包括中国对虾、凡纳滨对虾、日本对虾和斑节对虾等,都曾因病原性流行病而致使产业遭受严重损失,以1990年代暴发的病毒性流行病影响最为广泛,其中白斑综合征病毒(White spot syndrome virus, WSSV)是所有病毒性病原中传播面最广、破坏性最强的病原。
为摆脱病害困扰,对虾抗病品种的培育成为对虾病害防治的重要手段之一。世界各国先后开展了对虾抗病品种的研究,其中美国夏威夷海洋研究所(OI)培育出抗桃拉综合征病毒(Taura syndrome virus, TSV)的凡纳滨对虾品系[15],法国海洋开发研究院(IFREMER)筛选出抗IHHNV蓝对虾品系[16],中国水产科学研究院黄海水产研究所培育出抗WSSV的中国对虾“黄海2号”和“黄海5号”品种[17]。在抗性测试方法方面,对于细菌、病毒等流行性病原,一般采用人工侵染的方法测试家系及个体的抗病性能。以人工浸染WSSV为例,主要方法有三种,分别为浸浴病毒悬液、人工注射病毒粗提液以及投喂毒饵。周俊芳等[18]采用侵染WSSV的凡纳滨对虾取腹部肌肉经匀浆过滤制备了WSSV粗提液,通过人工注射粗提液开展了WSSV对凡纳滨对虾的致病性研究。李新苍等[19]通过建立实用WSSV定量检测方法开展了脊尾白虾(Exopalaemoncarinicauda)病毒侵染规律的研究。Meng等[20]建立了一种对虾抗WSSV能力的等量、定量测试方法,解决了目前对虾抗病性能测试过程中不能等量、定量侵染、不能控制对虾摄食量、不能准确分辨对虾是否摄食毒饵等无法避免的问题,提高了中国对虾抗WSSV性能测试的准确性。
1.2.3 抗逆性状 抗逆性状是对虾育种的关键目标性状之一。养殖环境中不良水质因子如异常的温度、盐度、氨氮含量、pH值以及亚硝酸盐等胁迫因子对对虾的遗传基础、抗病力以及病原微生物的致病力影响均十分显著,抗逆新品种的培育也成为对虾养殖业需要解决的问题。
目前,研究报道的与水质因子相关的抗逆性状主要以耐受pH、氨氮及亚硝酸盐居多,测试的方法主要通过设置不同浓度梯度开展致毒性、适应性研究[21-24]。潘鲁青等[25]、李润寅等[26]和马海娟等[27]通过设置不同温度梯度开展测试,分别对中国对虾、日本对虾和凡纳滨对虾的幼体、仔虾及成虾生长发育进行了研究, 获得了温度对对虾幼体发育、消化酶活力、温度适应性和耗氧速率的基础数据,为抗逆性状的选育提供了基础参考数据。对于盐度因子,主要的测试方法是针对盐度波动性、耐高盐、耐低盐等开展[28-30],以获得对虾在测试条件下的生长情况和存活情况数据。以凡纳滨对虾氨氮耐受性为例,首先通过预实验获得7 d内半致死氨氮浓度值,然后将测试家系个体物理标记后,在半致死氨氮浓度值下混合养殖测试,统计总体死亡率达到50%时各家系存活率或死亡率;或者每隔0.5到1 h捞取一次死虾,统计每尾虾的存活时间,据此进行遗传评估,选取抗逆性优良的家系及个体[31]。
1.2.4 繁殖性状 水产动物的繁殖性能决定着后代的产量和质量,是一项重要的经济指标。目前,在畜牧及家禽育种中,关于繁殖性状的遗传选育研究较多,技术手段也比较成熟。虾类开展繁殖性能的遗传选育研究起步较晚,繁殖性状的遗传力大部分为中低遗传力[32-35],对其进行遗传改良需要多代累加才能以获得更大的遗传进展。繁殖性状测试相对于其他性状测试来说,亲本培育周期长、测试时间长、人力物力成本高,因而在很多水产物种中很难开展此项工作。对虾繁殖性状测试前需要将对虾培育至性成熟,在测试开始前对所有雌虾采用眼柄环标记个体信息,并测量雌虾体重及体长,产卵后测量产卵量及卵径,孵化后统计孵化量,以及繁殖期内的首次产卵时间、产卵次数、产卵时间间隔、测试后体重等数据。Tan等以凡纳滨对虾为研究对象,开展了凡纳滨对虾繁殖力相关性状的测试,对115个家系共计1 428尾亲虾繁殖期的繁殖力和生长性能进行长达4个月的跟踪,统计产卵前和产卵后的体重、产卵次数、首次产卵日龄、产卵时间间隔等性状,结果显示产卵前和产卵后体重遗传力分别为0.58±0.08和0.52±0.08,产卵次数遗传力为0.07±0.02,首次产卵日龄遗传力高达0.92±0.08,产卵时间间隔遗传力为0.10±0.03,证实了生长速度与繁殖力间存在显著的权衡关系,为亲虾繁殖性能调控提供了重要的理论基础[36-37]。
1.2.5 饲料转化率 与畜牧动物相比,获取水产动物个体的摄食量数据更加困难,也严重限制了水产动物饲料利用效率研究的深入开展。在对虾饲料利用效率研究报道中,开展的基本是以群组为单位实施的饲料转化率测定,例如:凡纳滨对虾[38-39]、斑节对虾[40]。由于摄食方式的差异,X射线技术配合内含标记这种方法并不适用于对虾个体摄食量测定。在这种情况下,Dai等[41]研究人员首次利用循环水养殖系统,建立了大规模测定对虾个体饲料效率的方法,采用单独饲养的方式对中国对虾进行了个体摄食量测定,收集了来自中国对虾“黄海2号”育种群体中的51个家系共计400余尾对虾进行饲料效率测试,结果发现测试个体间的饲料效率比(Feed efficiency ratio, FER)存在很高的表型变异(0.055~0.593 g/d),同时,家系间也表现出较大的FER表型变异(0.115~0.388 g/d)。Dai 等[42]随后又以凡纳滨对虾为研究对象,开展了34个家系共计519尾对虾在连续两个阶段的饲料效率性状测试,结果显示个体间的FER和剩余摄食量(Residual feed intake, RFI)都存在较高的变异(FER:0.193~1.283 g/d;RFI:-0.068~0.092 g)。
1.2.6 出肉率性状 不同甲壳动物的出肉率存在较大差异,罗氏沼虾[43]、小龙虾和龙虾[44]等其他甲壳类动物的出肉率分别为37%~45%、15%~45%。同一物种在不同研究报道中的出肉率也存在一定差异,如陈晓汉等[45]测定了凡纳滨对虾的出肉率,其变化范围为53.03%~53.81%,与Kim等[46]报道的变化范围(52.1%~53.0%)相近,但低于Briggs等[47]报道的范围(66%~68%)。为了准确测定动物个体的出肉率,通常需要杀死受试个体,因此导致受试个体无法传代;这也意味着无法实施家系内个体选择,只能进行家系间选择[48]。为此,一些研究者为避免杀死受试个体,曾试图寻找更加合理的方法评估出肉率性状。各形态性状对体质量的多元分析已广泛应用于各种水产动物,常用方法主要有主成分分析、线性回归分析以及相关分析等;还有采用超声波、核磁共振等技术,通过测定肌肉横截面积、腹节周长等间接测定出肉率性状。
1.3 遗传和经济评估方法
1.3.1 方差组分估计方法 最早的方差组分估计方法是方差分析法(Analysis of variance, ANOVA),后来依次出现了Henderson方法I、II、III,最小范数二次无偏估计法(Minimum norm quadratic unbiased estimation, MINQUE)、最小方差二次无偏估计法(Minimum variance quadratic unbiased estimation, MIVQUE)、最大似然法(Maximum likelihood, ML)、REML以及贝叶斯方法等。这些方法各有特点,其中ANOVA、Henderson(I、II、III)、MINQUE和MIVQUE都存在方差组分的估计值可能为负的问题,主要见诸于早期的畜禽研究,由于对虾育种起步较晚,它们在对虾性状的方差组分估计中应用相对较少。何玉英等[49]利用两因素系统分组的方差分析计算中国对虾体长、体重等生长性状的方差组分,基于全同胞组内相关法估计获得这些性状的遗传力为0.16~0.43。徐如卫等[50]同样利用两因素系统分组的的方差分析法,估计了凡纳滨对虾的9个生长性状的方差组分,计算得到遗传力属于中高水平(0.251~0.460)。
贝叶斯方法也是目前水产动物中一种较为常用的方差组分估计方法,它结合了现有信息和先验信息,其后验密度等于先验密度与似然函数的乘积。贝叶斯方法在构建混合模型方程组时比较简便,由于模型中考虑了先验信息,可以弥补数据信息量不足的缺陷,保证其估计值更加精确,且方差较小。但贝叶斯方法存在收敛速度慢的问题,尤其在参数先验信息缺失时,如果没有附加的假设,就不能用数学描述出统计分布函数,而这些假设得不到验证将导致计算量比较大。一般在REML等方法估计方差组分出现无法收敛的情况时,可以借助贝叶斯方法实现方差组分的估计。Dai等[42]采用动物模型结合基于Gibbs抽样的贝叶斯方法估计了凡纳滨对虾剩余摄食量、饲料效率比、平均日增重等性状的方差组分,其中加性方差在表型总方差中的占比较大。一般而言,方差组分估计的准确性取决于性状分布特征、群体结构、记录的完整性、统计模型及估计方法等,所以应该根据实际的数据资料来筛选并确定最合适的方差组分估计方法。
1.3.2 育种值评估方法 目前最主流的育种值估计方法是BLUP,仅借助一个混合模型方程组既能估计出固定的环境和遗传效应,又能估算出随机的遗传效应。在使用动物模型时,BLUP方法可以充分利用亲属信息,消除固定环境及遗传效应的偏差,校正由于选配所造成的偏差,能够对来自不同年份、世代、群体、年龄、信息量的个体进行育种值估计[8]。BLUP方法在处理不平衡数据时,结合选择指数法和最小二乘法两者的优点,利用线性模型和混合方程组同时得到固定效应的最优线性无偏估计和随机效应的最优线性无偏预测,而且极大地简化了矩阵运算,提高了速度。相比直接基于表型选择,基于BLUP方法的选择要更加准确[53]。借助BLUP方法,中国目前已有多个对虾新品种问世,如罗氏沼虾“南太湖2号”、中国对虾“黄海2号”和“黄海5号”,以WSSV抗性、生长速度和养殖存活率等作为育种目标性状,采用多性状选择育种技术经多代培育而成;凡纳滨对虾“海兴农2号”是以提高凡纳滨对虾生长速率和养殖存活率等性状为选育目标,经5代的家系选育获得。
分子标记技术的发展,尤其是高通量SNP标记的出现,通过覆盖全基因组范围的分子标记信息可以计算个体间的分子亲缘相关度,构建分子亲缘关系矩阵(G矩阵),以代替基于系谱的A矩阵进行GBLUP(genomic BLUP)评估,进一步提高选择的准确度。还可以将分子标记信息构建的G矩阵与系谱信息构建的A矩阵复合成为H矩阵,进行一步法GBLUP(single-step genomic BLUP, ssGBLUP)[54]。Dai等[41]利用部分家系亲本的基因型信息结合多代系谱信息构建了H矩阵,对中国对虾“黄海2号”育种群体的平均日增重和饲料效率比进行了的育种值预测,预测准确率分别为0.36和0.27,相比只基于系谱信息的预测结果并没有显著差异。
此外,假定覆盖全基因组范围的高密度标记与 QTL处于连锁不平衡状态,也可以预测出标记的效应值,进而获得个体的基因组估计育种值[55],进行基因组选择。常用的算法包括:岭回归最佳线性无偏估计(Ride regression best linear unbiased prediction, RRBLUP)、贝叶斯算法A(BayesA)、贝叶斯算法B(BayesB)和贝叶斯算法C(BayesC)及贝叶斯收缩算法(Bayesian least absolute shrinkage and selection operator, Bayeisan LASSO)等,这些方法的主要差别在于关于标记效应方差的先验分布有不同的假设。但到目前为止,对虾中仅有少量基因组选择的研究报道。Wang等[56]利用RR-BLUP、BayesA和Bayesian LASSO三种模型预测了凡纳滨对虾体长和体重性状的基因组育种值,三种模型的平均预测准确率分别为0.296和0.411。
1.3.3 间接遗传效应评估方法 在经典的数量遗传学研究中,由于缺少可行的实验设计和分析技术,在遗传评估中仅考虑直接遗传效应(Direct genetic effect, DGE),无法评估竞争、协作等个体间的社会交互行为对育种目标性状的影响。2005年,Muir等扩展了传统的线性混合模型,评估了日本鹌鹑个体间的竞争行为,发现在遗传评估时不考虑由竞争产生的遗传效应会导致体重、存活率和饲料转化效率等性状产生负向选择反应。在现代数量遗传学中,将个体自身行为对同一组内其他同伴性状值产生的遗传效应,称为间接遗传效应(Indirect genetic effect, IGE)[57]。为了准确地剖分出IGE,在虾类中多采用多家系多分组实验设计[58-59]。实验设计为:针对g个家系,设置g个测试池;每一个家系均分为m个分组,每个分组包括n尾个体;m个分组分别放置到m个测试池中;每个测试池包括m个不同家系的个体;任意两个家系组合在同一个测试池中出现的次数最多为1次;一个家系将在m个测试池中,与m×(m-1)个不同家系组合测试。该设计方案的优点是需要的测试池少,已在大西洋鳕鱼、凡纳滨对虾和中国对虾等核心育种群体IGE分析中应用(m=3),测试池的数量100个以上[59-61]。另外一种设计方案是先将g个家系随机划分为b个区组;每个区组内m个家系两两组合在一起放入一个测试池[72]。该方案需要的测试池数量多,已在罗非鱼核心育种群体收获体重IGE分析中应用(m=2),测试池的数量超过490个[62]。已有研究表明,为了准确估计IGE的方差组分,测试池的数量需要大于250个[63]。
1.3.4 经济评估方法 动植物遗传改良的最终目的是获得更高的经济效益,早在1940年代就有学者提出综合考虑性状本身遗传力、性状之间遗传相关及相对经济重要性等参数,使用以货币为单位的综合育种值来量化育种目标[64],这是最早的经济评估理念,后来进一步发展为建立在单位生产的利润、投资效益或成本基础上的育种目标[65]。最早的经济评估出现在畜牧育种中[66-67],水产动物的此类研究最早在鱼类中开展,如针对大西洋鲑的生长速率和性成熟年龄[68],以及GIFT罗非鱼和鲤鱼的收获体重、存活率和饲料摄入量性状[69-70]等性状的遗传增益进行经济评估。
事实证明,中邮速递易在2012年率先布局智能快递柜的决定是明智的。根据国家邮政局《2017年快递市场监管报告》显示,2017年,中国快递业务量达到400.7亿件,占据世界份额超40%,连续4年稳居世界第一。同时,全年日均快件处理量1.1亿件,五年来提升超3倍。最高日处理快件量超3.3亿件,同比增31.9%。而国家邮政局公布的最新数据显示,2018年上半年预计累计完成快递业务量达220.8亿件,超过我国2015年全年快递业务量。天量的快递规模,让传统的人工配送短板尽显,智能快递柜的春天悄然而至。
大部分对虾育种项目的育种方向是围绕着提高生长、抗病和抗逆等重要经济性状的选择反应展开,对育种项目进行遗传和经济评估的前提是研究对象具备较为成熟的育种体系。目前,已在凡纳滨对虾和中国对虾中开展了经济评估的相关工作[71-72]。结合生物学、遗传学和经济学参数,采用“投入-产出”法,以收获体重、存活率和饲料摄入量为目标性状,对执行多性状复合育种方案20个世代的遗传进展和经济效益进行预测和评估,证明该育种方案可以显著地改良目标性状,产生较好的经济效益,而且利用对虾的高繁殖力特性,在核心群体中取得的遗传增益通过三级金字塔传递系统快速传递和放大,可以获得更高的经济效益。在凡纳滨对虾经济评估研究中,进一步考虑了育种目标性状在不同的区域间存在较强的基因型与环境互作效应等情况对经济效益的影响。与杂交育种、细胞工程育种和转基因等育种技术相比较,多性状复合育种技术的一个显著特点是目标性状的遗传进展会随着世代增加不断提高,这决定了在整个育种周期内目标性状累积遗传进展产生的经济效益将十分可观。
2 重要经济性状的遗传参数估计结果
2.1 遗传力
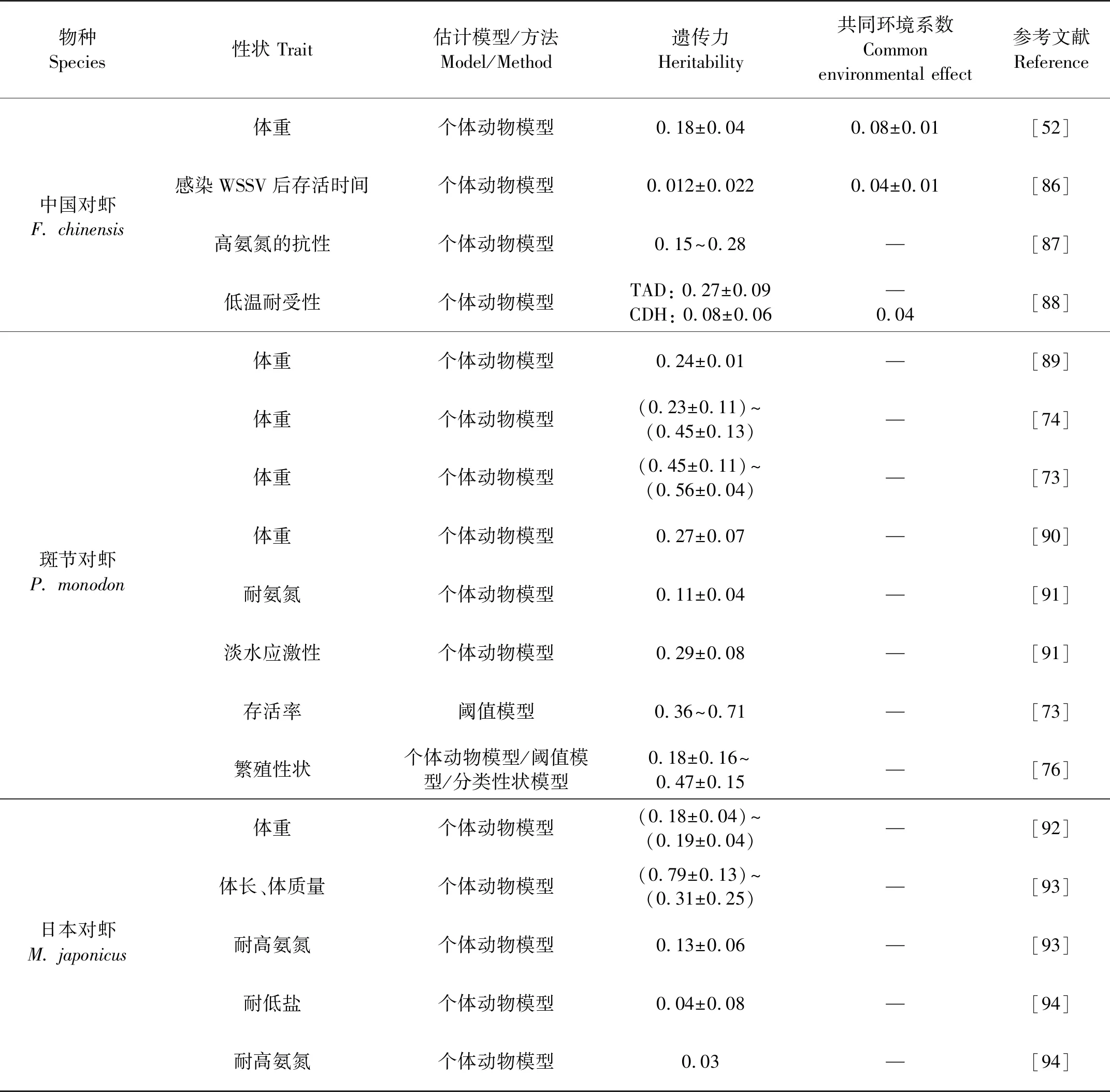
查清重要性状在育种群体中的遗传变异丰富度及相关关系,是设计育种方案,培育多性状优良新品种的基础。表2汇总了已报道的主要对虾养殖种类生长速度、WSSV和TSV抗性、氨氮耐受性、低温耐受性、出肉率、饲料利用率、繁殖力等重要经济性状的遗传力估计值。
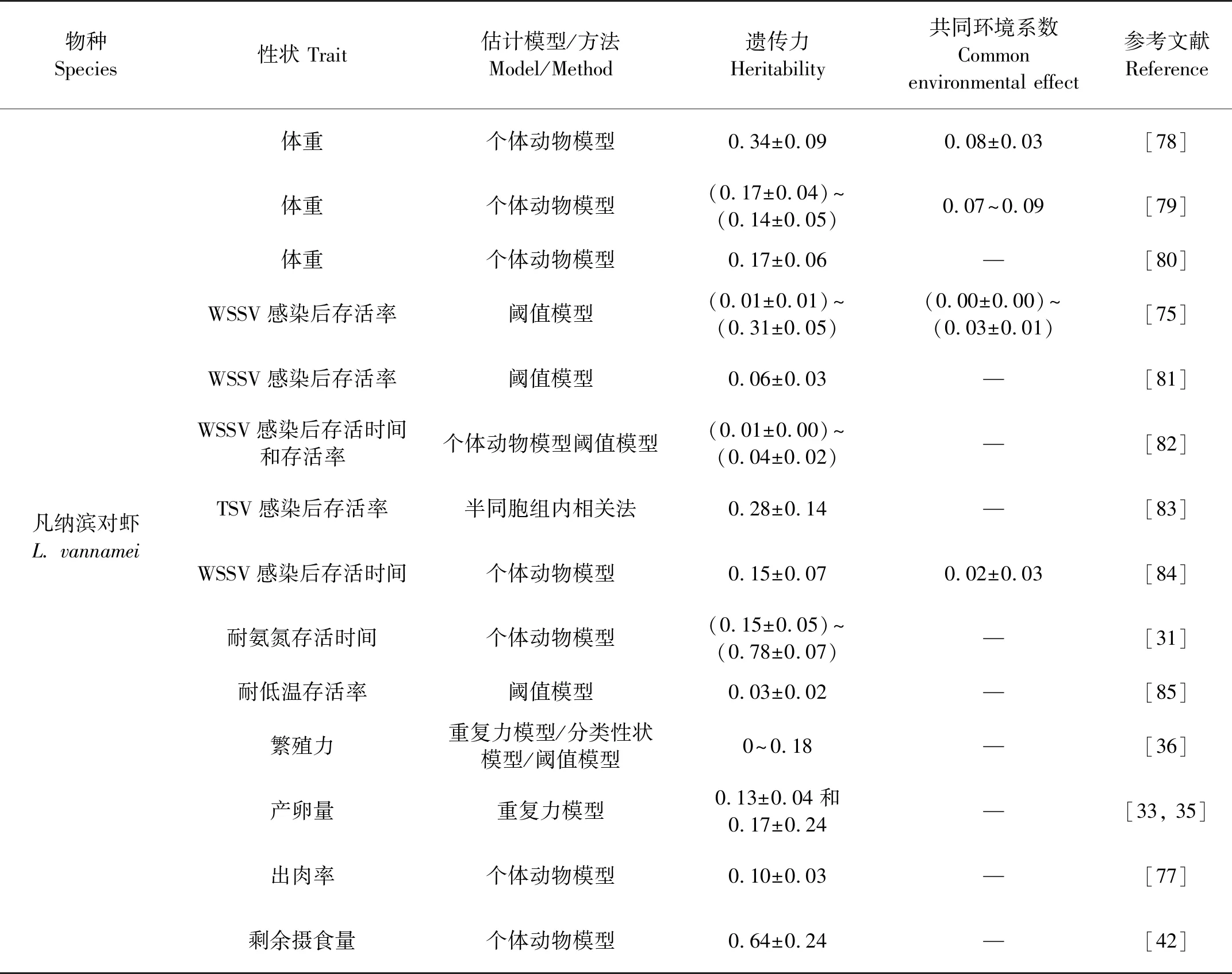
表2 对虾核心育种群体重要经济性状的遗传参数
续表2
对于体重等生长性状,其遗传力估计值在0.17~0.56之间,表现为中高遗传力水平,因此体重和生长速度等性状具有较大的选择潜力,目前已经培育成功的“快大”品系主要是针对此性状选育而成的,其生长速度表现出明显的优势。因无法剖分共同环境效应,部分斑节对虾育种群体体重遗传力估计值可能偏高[73-74]。Dai等利用个体动物模型获得了凡纳滨对虾饲料利用效率性状的遗传参数,其中饲料转化率和剩余摄食量性状的遗传力分别为0.58±0.23和0.64±0.24,该结果未剖分共同环境效应[42]。该研究首次在个体水平上报道了虾类育种群体饲料利用效率性状的遗传参数。
对于疾病抗性,如感染WSSV后存活时间或存活率,多表现为低遗传力水平(<0.15), 因此针对此性状的选育进展相对较慢,需要连续多代开展才能获得一定的遗传增益。Trang等研究表明,WSSV侵染后3 d幼虾存活率的遗传力为0.31±0.05,随着病毒侵染时间的增加,整体存活率逐渐接近50%,遗传力不断降低,侵染15 d后幼虾存活率的遗传力为0.01±0.01[75]。对虾氨氮耐受性的遗传力估计值多表现为中低遗传力水平(<0.15)。但正常盐度和低盐度环境下凡纳滨对虾仔虾(0.50 g/尾)耐氨氮存活时间的遗传力分别为0.78±0.07和0.58±0.07,均表现为高遗传力水平,该研究结果未剖分共同环境效应[31]。目前,市场上广泛推广的“高抗”系列品系主要是针对抗病、抗逆或存活等性状开展选育,表现出存活率高、抗病性强、抗应激性强等特性,适合土塘和大水面生态养殖模式。
对于繁殖力性状(雌虾产卵量、无节幼体数、卵径、产卵频次和是否产卵),大多数性状遗传力较低,在0~0.18之间[36]。因此,选育高繁殖力的品系的过程也相对较慢,需要连续多代选育以获得较高的遗传增益。Arcos[33]和Caballero[35]利用重复力模型估计的产卵量遗传力分别为0.13±0.04和0.17±0.24,同样为中低遗传力水平。斑节对虾繁殖性状(产卵次数、产卵数量、无节幼体数量)的遗传力在(0.18±0.16)~(0.47±0.15)之间,为中高遗传力,但该值未剖分共同环境效应[76]。
对于出肉率性状,基于16个SSR分型标记信息,构建了75个家系1 244尾个体间的分子亲缘相关度,利用个体动物模型获得了出肉率的遗传力为0.10±0.03[77],为低遗传力水平。
值得注意的是,部分遗传力估计值受限于群体遗传变异、家系管理标准化程度、性状特征、分析方法与模型等,估计值可能会存在偏差。尤其是水产动物养殖中存在的共同环境效应的问题,可能会影响到遗传力估计的准确性。
2.2 遗传相关和基因型与环境互作(Genotype by environment interaction, G×E )
对虾产业环节多,对品种的性状要求也相应多,性状间的遗传相关性就显得特别重要。同时,有的种类,如凡纳滨对虾,养殖区域广、养殖的环境条件、养殖模式等都呈现高度多样性,基因型与环境互作研究也备受重视。表3汇总了国外文献中报道的主要对虾养殖种类不同性状的遗传相关,以及在不同养殖环境、模式下的基因型与环境互作效应。
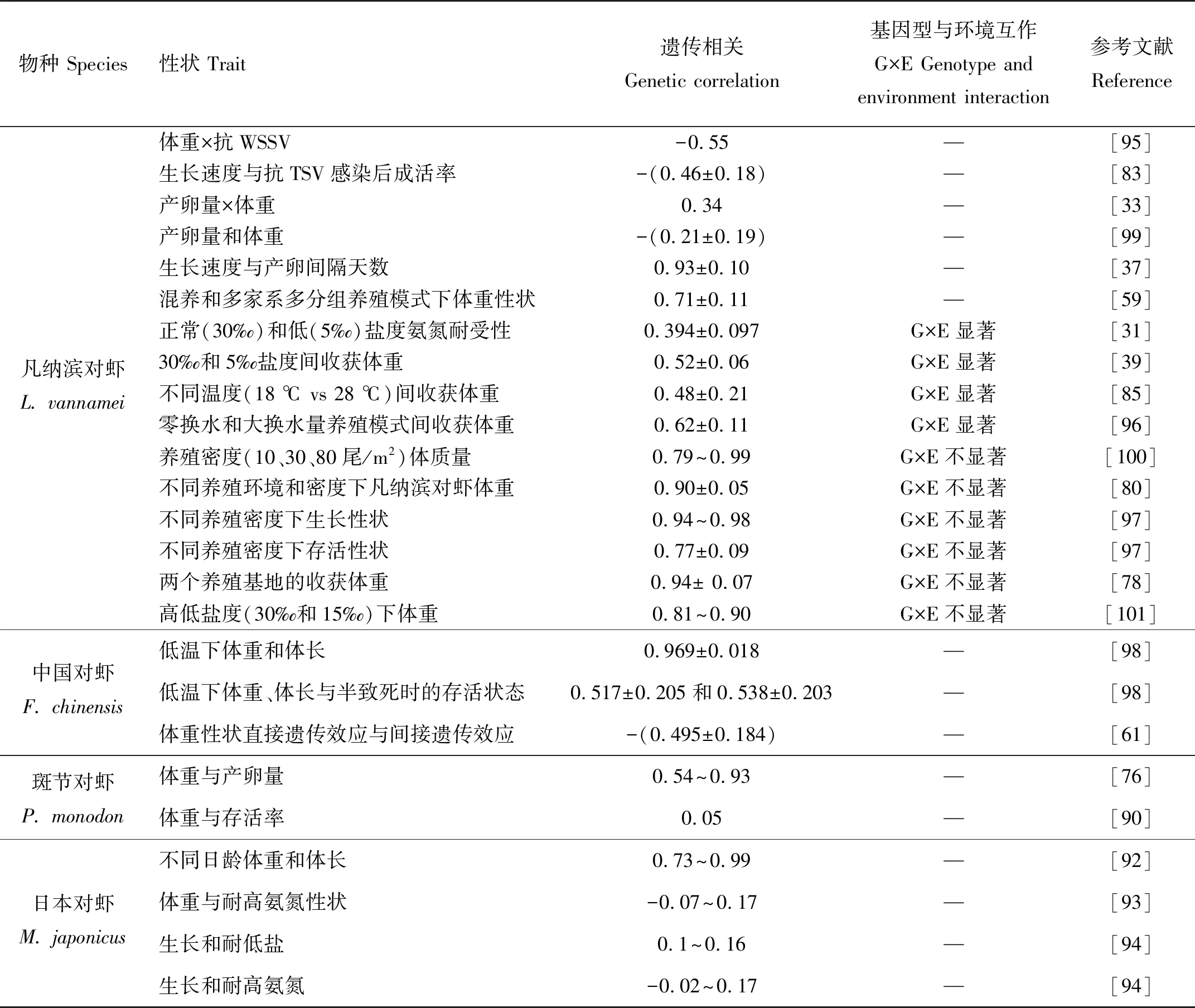
表3 对虾核心育种群重要经济性状的遗传相关
报道分析表明,对虾生长速度与抗病性状间普遍存在负相关关系。凡纳滨对虾体重与WSSV抗性间遗传相关为-0.55[95],体重与抗TSV感染后成活率之间的遗传相关为-(0.46±0.18)[83]。在生长速度与繁殖力的相关分析研究中,有报道个体体重与产卵间隔天数的遗传相关高达0.93±0.10,提示生长速度与繁殖力间存在显著的负相关关系[37]。不同生长发育阶段体重与产卵量的相关关系分析则有不同的报道,有的研究结果显示为正相关关系,也有的显示为负相关关系,但总体上相关性不强。基因型与环境互作方面,已有结果表明,养殖模式及环境间差异越大,基因型与环境互作效应越强。凡纳滨对虾仔虾在正常(30‰)和低(5‰)盐度条件下氨氮耐受性的遗传相关系数仅为0.394±0.097,表明存在较强的G×E效应;30‰和5‰盐度间收获体重表现为中低度遗传正相关(0.52±0.06),G×E效应显著[31]。凡纳滨对虾不同温度(18、28 ℃)间收获体重的遗传相关较低(rg=0.48±0.21,K>0.50),G×E效应显著[85]。凡纳滨对虾零换水和大换水量养殖模式间收获体重的遗传相关系数为0.62±0.11,也表明两种养殖模式间存在一定程度的G×E效应[96]。凡纳滨对虾在高低密度(80尾/平方米和160尾/平方米)间各性状的遗传相关介于0.94~0.98之间,为高度遗传相关,显示两个密度间存在较低的G×E效应;高低密度下存活性状的遗传相关为0.77±0.09,该值与1没有显著性差异(P>0.05),说明存在较小的G×E效应[97]。
针对中国对虾,低温下体重、体长之间的遗传相关分别为0.969±0.018,为高度正相关;低温下体重、体长与半致死时的存活状态的遗传相关分别为0.517±0.205和0.538±0.203,为中度正相关[98]。郑静静[94]对68个日本对虾家系45日龄和75日龄体重和体长性状的遗传相关估计值介于(0.505 2±0.028 0)~(0.989 6±0.003 4)。蒋湘等[93]估计了日本对虾基础群体的体长、腹长、体质量与耐高氨氮性状的遗传相关,体长、腹长、体质量与耐高氨氮性状的表型相关与遗传相关系数分别为-0.082~0.08和-0.067~0.17,检验结果不显著。周发林等[91]以高氨氮和淡水胁迫致死时间为衡量指标,估计了斑节对虾耐氨氮和淡水应激性状的遗传参数,斑节对虾耐氨氮和淡水应激性状的家系表型值相关系数为0.15,表现为低度线性正相关。斑节对虾耐氨氮和淡水应激性状的家系育种值相关系数为0.57,表现为中度线性正相关。
2.3 间接遗传效应
尽管对虾的行为学研究资料匮乏,但同时出生的对虾,在相同的环境中养殖,其个体大小有时表现出很大变异,个体间的社会交互行为产生的间接遗传效应可能是重要原因,其研究越来越受到重视。以凡纳滨对虾核心育种群G1代105个家系为研究对象,在低密度养殖模式下,采用多家系多分组设计,利用传统动物模型和包含间接遗传效应的扩展动物模型估算了收获体重的遗传参数。在虾类育种群体中获得了个体间由社会交互产生的IGE方差,占总遗传方差的比例为54%;总遗传方差占表型方差的比例为73%。基于多家系多分组新模式测定的总遗传变异是传统模式的2倍多[59]。进一步采用多家系多分组设计,限制配合饲料投喂量为正常投喂量的50%,对中国对虾G11代核心育种群103个家系进行IGE分析,结果表明:在竞争性环境下,IGE对收获体重总遗传方差的贡献度达75%;较大的负DGE-IGE协方差(-2.20)导致总遗传方差变小,与表型方差的比值仅为52%,小于传统的遗传力估计值(62%);负的DGE-IGE相关系数(-0.50±0.18)表明测试群体内个体间存在较强的竞争交互行为。研究表明,在饲料投喂量受限制的竞争性环境下,对虾个体间较强的竞争交互行为对中国对虾收获体重间接产生了可遗传的效应,减少了可利用的总遗传变异[61]。上述研究首次在虾类育种群体中,定性定量解析了由对虾个体间社会交互产生的间接遗传效应,发现了隐藏的新遗传变异,为对虾精准育种奠定了理论和技术基础。
3 遗传增益
选择导致的群体平均值的变化称为选择反应(Selection response),是中选亲本的后代与选择前亲本世代之间的平均表型值之差。选择反应除以亲本群体的平均表现型值,所得百分率,为遗传增益(Genetic gain)[102-103]。
3.1 国外文献报道
表4汇总了外文文献中报道的对虾选择育种项目中重要经济性状的遗传增益。
3.1.1 生长性状 在14项对虾遗传增益估计的研究中,有9项是关于体重遗传增益的研究。体重每代遗传增益2.3~21.0%,平均8.10%。其中5项研究的遗传增益是基于单性状开展的选育项目,4项研究是基于多性状开展的选育项目。多性状选育的平均增益是8.3%,与所有生长性状的遗传增益估计值的总体平均值非常接近。与传统家畜育种的结果相比,这是一个显著的高遗传增益[104]。对虾育种中,生长的遗传增益与鱼类相比相对稍低,在23种甲壳类育种研究中,平均每代遗传增益为9.7%,而在44种鱼类中,平均每代遗传增益为14.3%[104]。根据对虾育种的遗传增益,经过7代的生长选择,生长速度可以翻一番。而对于草鱼和大西洋鲑鱼等具有长世代间隔的其他水产物种,可能需要长达28年的时间才能达到这一进展。
3.1.2 抗病性状 对虾中特定疾病抗性的选育进展集中在凡纳滨对虾中,针对不同疾病每代遗传增益1.7~18.4%,平均10.6%。其中WSSV的抗性选育进展较低,每代平均遗传增益仅4%,TVS遗传增益15.4%。抗病性状的高遗传增益显示了针对特定的病原培育高耐受性或者可能真正抗性品系是可行的。而养殖存活率性状虽然受到多种环境因子的影响,仍可以达到5.7%的遗传增益。对多数养殖者来说,疾病是对虾养殖中面临的主要问题。目前研究表明基于感染测试结果进行选育是非常有效的[105]。这种方法结合QTL以及标记辅助或基因组选育,非常有希望在未来降低对虾的死亡率。
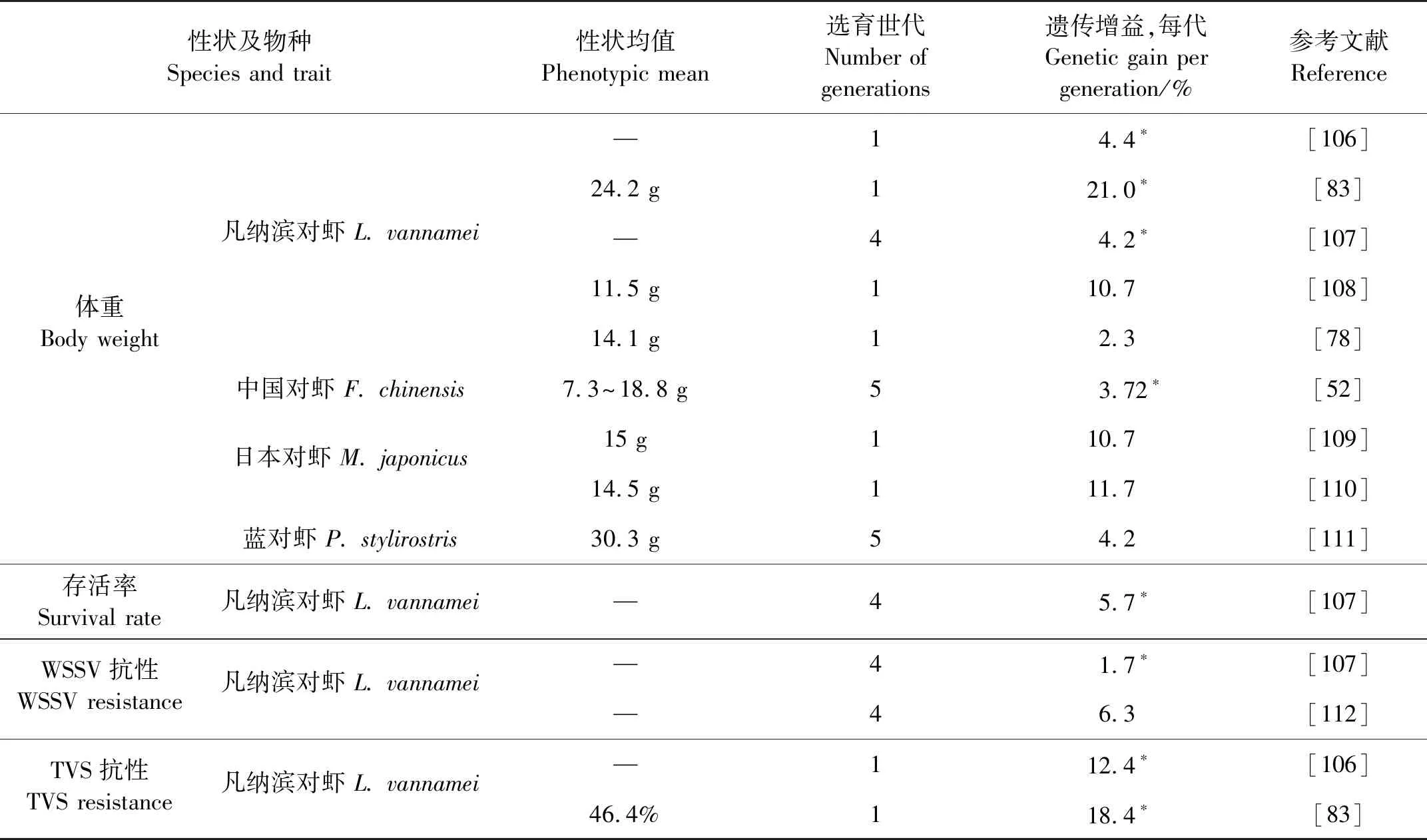
表4 对虾核心育种群重要经济性状的遗传增益
3.2 国内遗传增益的报道
到目前为止,中国自主培育的对虾新品种共有15个,其中凡纳滨对虾9个,中国对虾4个,斑节对虾1个,日本对虾1个(见表1)。选育性状以生长速度为主,囊括了成活率、抗病、抗逆等多类性状。其中,凡纳滨对虾以家系选育为主要选择方法,结合群体选育、品系选育、杂交制种等多种方法进行新品种培育,各性状每代平均遗传增益为生长性状5.74%,成活率3.90%,WSSV抗性6.00%;中国对虾以家系选育和群体选育为主要选择方法进行新品种培育,各性状每代平均遗传增益为生长性状5.01%,成活率2.72%,WSSV抗性4.49%;斑节对虾以群体选育为主要选择方法进行新品种培育,各性状每代平均遗传增益为生长性状3.39%,氨氮胁迫成活率3.10%;日本对虾以家系选育为主要选择方法进行新品种培育,生长速度每代平均遗传增益为4.08%。这些新品种的培育为中国对虾养殖业自给自足奠定了基础。
4 展望
4.1 高通量性状测定技术
利用精确高效的性能测定系统,高通量、高质量和可重复地获取性状数据,是开展选择育种研究的前提基础。在植物、畜牧育种研究中,利用遥感、机器人技术、计算机视觉和人工智能等技术,已经实现了自动化测量,极大提高了育种效率[113]。而水产动物,特别是对虾,个体小、养殖密度高、数量多、环境复杂,难以实时监控、自动化、规模化获得个体的表型数据。当前,人工测量是获得对虾个体表型性状的主要方法,但存在耗时长、易产生人为误差等问题,难以规模化采集数据,已经成为阻碍对虾选择育种发展的一个重要问题。近些年,机器视觉技术已在鱼类和贝类中应用,测量外型、大小、重量性状的准确度可达到> 0.95;近红外光谱和高光谱成像技术,应用在脂肪、蛋白、纹理、颜色、新鲜度等品质性状的测量中,准确度可达到0.8[114]。与硬件设施配套的图像识别算法,如基于机器学习的卷积神经网络算法,可进一步提高测量的准确度。可以预期,逐步发展起来的表型组学将是突破未来对虾育种研究和应用的关键研究领域,通过高通量表型分析来描述重要经济性状可以为育种决策提供大数据支持,大幅度提升选育效率。
4.2 基因组选择技术
传统的BLUP选育,对于中高遗传力性状,选择反应大,遗传增益显著;而对于抗病、抗逆等低遗传力性状以及一些限性性状,难以准确剖分出加性遗传变异,选育进展缓慢。抗病和抗逆等复杂性状是由多基因和非遗传因素共同作用的复杂性状,遗传机制复杂,受环境影响大,不同种类的抗性存在较大差别。对于抗性、肉质等性状,传统育种方案多通过同胞测试方法进行家系选择,仅可利用一半的加性遗传变异[115],进一步降低了选育效率。利用覆盖全基因组的分子标记,通过基因组选择方法,可以准确刻画全同胞家系内个体间的亲缘关系,提高家系选择的效率[55]。当前,限制基因组选择在对虾选择育种中应用的主要因素有两个:(1)与牛、猪等大动物相比,单尾对虾的商品价值较低,且种虾的使用寿命短;(2)目前缺少可用的商业化SNP芯片,测序分型的成本仍然比较高。但从另外一个角度看,对虾的繁殖力高,每尾雌虾平均产卵量高达30~50万粒。因此选育出的优良种虾,其遗传增益可以迅速传递到生产群,其价值要远高于作为普通商品虾的价值。且随着凡纳滨对虾基因组的破译[116]以及wssGBLUP等新方法的不断发展[54],可以同时利用分型个体和未分型个体系谱和性状测定信息进行评估,因此仅对部分个体进行高通量SNP分型也可以提高遗传评估的准确性,降低了分型成本。此外,随着液相与固相分型技术[117]、GBS(Genotyping-by-sequencing)等高通量SNP分型与填充技术[118]的不断发展,个体高通量SNP分型的成本也在不断降低。未来2~3年,预期每个对虾样本10 K量级的SNP分型成本可控制在100元之内,每个世代可对千级规模的样本开展基因组选择育种,进一步提高抗性和屠宰等性状的选育效率。
4.3 高强度选择模式
当前已报道的大部分对虾规模化家系选育计划,家系数量和家系内测试个体数量大多在百级水平,存在进一步突破的空间。遗传进展主要取决于目标性状的遗传力和选择强度。在已完成基础群体构建,遗传力等参数已固定的情况下,扩大选育群体的规模,提高选择强度,是进一步提高遗传进展的主要途径之一。对虾繁殖力高,开展家系内高强度选择,可进一步增加选择反应。凡纳滨对虾模拟研究表明[119],与对照方案(家系数量100个、家系内测试个体数100个、体重遗传力0.35)相比,增加参与测试的家系内个体数(100~5 000),不断提高选择强度,核心育种群收获体重育种值均值增加了48%(62.75~92.87 g)。在实际育种计划中,可以结合已有设施,分阶段多次执行家系内数量标准化,不断提高选择强度;也可结合SSR、SNP等分子标记,在仔虾期大规模混养家系个体,通过亲子鉴定等方法高强度筛选育种目标性状优良的个体作为留种亲本,增加选择反应。
