γ-氨基丁酸对乌鳢生长性能、肠道消化酶和抗氧化酶活性的影响
2020-08-12彭思博陈秀梅孔雨昕王桂芹
彭思博 陈秀梅 孔雨昕 王桂芹
(吉林农业大学动物科学技术学院,吉林长春130118)
乌鳢(Channa argus),属鲈形目(Perciformes)、鳢科(Channidae)、鳢属(Channa),又称黑鱼、乌鱼、财鱼等,广泛分布于我国南北水域以及亚洲南部、东南部国家,乌鳢、斑鳢及其杂交鳢是主要养殖品种[1]。目前,我国主要乌鳢养殖省份为广东、山东、江西、湖南、浙江和湖北。因其生长快、生命力强、没有肌间刺,深受消费者的青睐,被喻为高投入、高产出、高效益的养殖品种[2]。但在水产集约化养殖的今天,养殖户在获得最大经济效益的同时,也造成了各种各样问题,如环境污染、水质恶化、鱼类疾病频发等[3]。其中,乌鳢作为凶猛肉食性鱼类,因投喂冰鲜野杂鱼容易造成环境污染、浪费饲喂原料等原因,一般采用驯化摄食配合饲料的方法来替代冰鲜野杂鱼进行养殖,但是存在摄食率低、抗病力弱等问题[4]。基于此,寻求一种高效安全的饲料添加剂,通过营养调控手段提升乌鳢的抗应激能力势在必行。
γ-氨基丁酸(γ-aminobutyric acid,GABA),又称为4-氨基丁酸,是一种具有生物活性的重要功能性四碳非蛋白质氨基酸,于1949 年在马铃薯的块茎中被首次发现,又于1950 年在哺乳动物的中枢系统中被发现,其广泛存在于动植物体及微生物体内[5-6]。作为重要抑制性神经递质的一种,大量研究证明其在调控机体生长、食欲和抗应激等方面有独到的生理功能,在肉仔鸡的抗应激能力[7]、崇仁麻鸡的生长性能[8]、断奶仔猪的抗氧化能力[9]、三元杂交猪的饲料利用能力[10]、鲶(Clarias sp.)的性腺功能[11]、鳜(Siniperca chuatsi)的摄食能力[12]、高体鳑鲏(Rhodeus ocellatus)的抗缺氧能力[13]和南美白对虾(Litopenaeus vannamei)的生长性能[14]方面都有相关报道,但目前尚未有研究报道GA⁃BA 对乌鳢的影响作用。因此,本试验通过外源添加GABA,以乌鳢作为研究对象,探讨GABA 对乌鳢生长、肠道消化酶和抗氧化酶活性的影响,旨在为GA⁃BA在乌鳢的健康养殖中的应用提供参考依据。
1 材料与方法
1.1 试验饲料
试验饲料以鱼粉、玉米蛋白粉为蛋白源,在基础饲料中分别添加0、30、60、90、120 mg/kg 的GABA,配制等氮等能(42% CP、18 MJ/kg)的饲料。将饲料原料粉碎后过孔径0.246 mm(60 目)的筛,按饲料配方称取各原料,加工成直径为2.0 mm 的颗粒,通过晾干后,再置于-20 ℃冰柜中进行保存备用。根据GB/T 6435—2006、GB/T 6432—1994、GB/T 6438—2007 和GB/T 6433—2006 法分别测定全鱼中的水分、粗蛋白质、粗灰分和粗脂肪的含量。试验用基础饲料配方及营养水平见表1。

表1 基础饲料配方及营养水平(干物质基础)
1.2 试验设计
选取健康、体型及规格一致的乌鳢([10.13 ±0.51)g/尾],在吉林农业大学控温养殖系统中暂养15 d,饱食投喂基础饲料,每日投喂2次(9:00、16:00),投喂1 h 后对水族箱内残饵的情况进行检查,有沉积的残饵采用虹吸法将其吸出,并根据残饵量来对投喂量进行相应调整,暂养期间控制水温在23~25 ℃,溶解氧大于5.0 mg/l,pH 值7.1±0.1。试验前禁食24 h,将450 尾乌鳢随机分成5 组,分别饲喂上述5 组饲料,每组3个重复,每个重复30尾鱼,饲养试验为期56 d,期间日常管理及水质条件同暂养期间保持一致。
1.3 样品的采集
56 d饲养试验进行完毕后准备取样,取样前停食24 h,称量每个桶内的乌鳢总质量,清点并记录每桶鱼的尾数。然后,每桶随机取10 尾鱼,采用MS-222(Sigma,10 mg/l)麻醉后,在吸干鱼体表的水分后,称质量,冰上解剖取肠道组织于-80 ℃冰箱保存待测。
1.4 指标的测定
1.4.1 生长指标检测
根据乌鳢初始体质量、终末体质量以及摄食量计算平均增重率(AWGR)、特定生长率(SGR)、饲料效率(FER)、蛋白质效率(PER)。
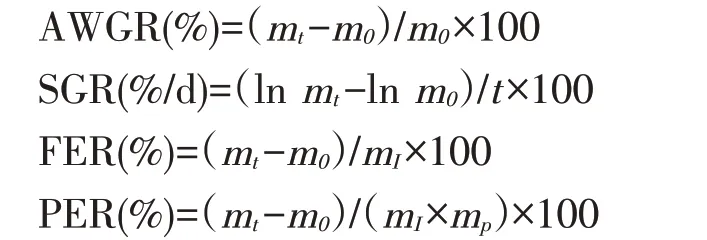
式中:m0、mt——分别表示鱼初始体质量和终末体质量(g/尾);
t——试验时间(d);
mI——摄入干饲料质量(g/尾);
mp——饲料粗蛋白质含量(%)。
1.4.2 肠道消化酶活性的测定
乌鳢的肠道胰蛋白酶和脂肪酶活性采用购自南京建成生物工程研究所的试剂盒进行相应检测。
1.4.3 肠道抗氧化酶活性的测定
乌鳢的肠道总超氧化物歧化酶(T-SOD)、过氧化氢酶(CAT)和谷胱甘肽过氧化物酶(GSH-Px)活性采用购自南京建成生物工程研究所的试剂盒进行相应检测。
1.5 数据统计分析
采用SPSS(20.0)软件对乌鳢的生长性能、肠道消化酶和抗氧化酶相应指标数据进行方差分析,如果方差分析显著,则进一步进行Duncan's法多重比较分析组间差异显著性,显著水平设定为P<0.05,试验数据均采用“平均值±标准差”表示。
2 结果
2.1 GABA添加水平对乌鳢生长性能的影响(见表2、图1~图2)

表2 GABA对乌鳢生长性能和饲料利用的影响
由表2可知,随着GABA添加量的增加,乌鳢生长水平逐渐上升,在GABA 添加量为30~120 mg/kg 时,AWGR、SGR 均显著高于对照组(P<0.05),但添加GABA 的各组间差异不显著(P>0.05);FER 在添加量为60~120 mg/kg 时显著高于其他各组(P<0.05),且在添加量为90 mg/kg 时达到最高;添加GABA 组的PER 显著高于对照组(P<0.05),其中添加量为90、120 mg/kg 组显著高于30 mg/kg 组(P<0.05),且在添加量为120 mg/kg 时达到最高。由图1 可知,饲料GABA 添加量(x)与平均增重率(y)的回归方程为y=-0.009 1x2+1.611 5x+302.42(R2=0.946 2),当ymax时,x=88.54。在本试验条件下,将抛物线回归分析结果与方差分析结果结合起来可知,乌鳢最大平均增重率时的最适GABA 添加水平为88.54~90 mg/kg。由图2可知,饲料GABA 添加量(x)与饲料效率(y)的回归方程为y=-0.001 3x2+0.267 1x+60.621(R2=0.989 8),当ymax时,x=102.73。在本试验条件下,将抛物线回归分析结果与方差分析结果结合起来可知,乌鳢最大饲料效率时的最适GABA添加水平为90~102.73 mg/kg。

图1 饲料GABA添加量(x)与平均增重率(y)的关系
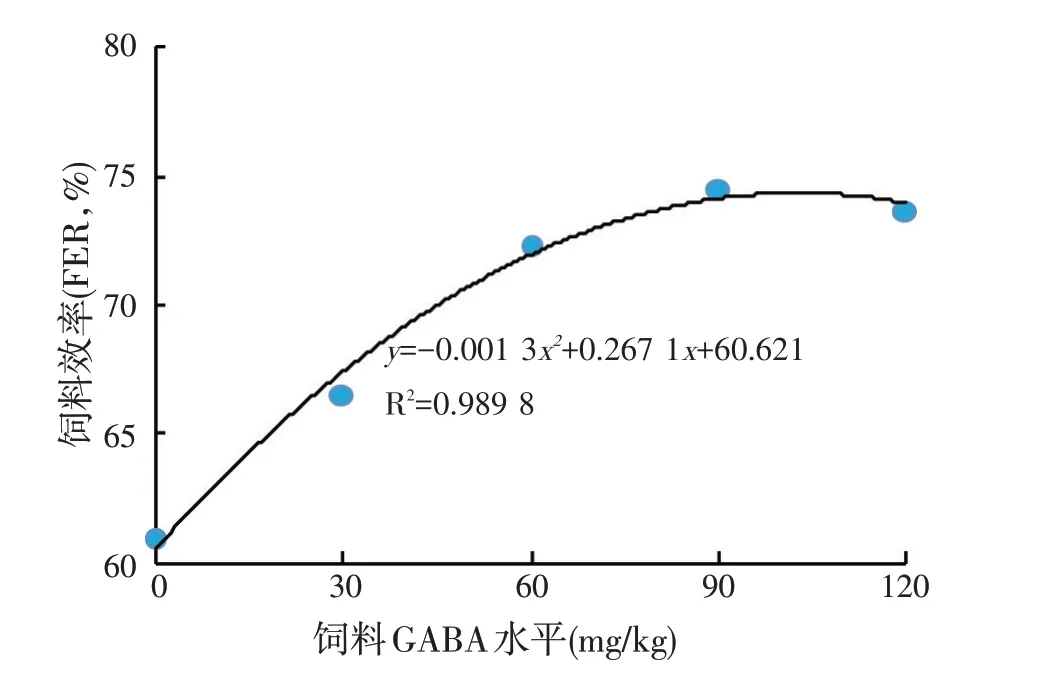
图2 饲料GABA添加量(x)与饲料效率(y)的关系
2.2 GABA对乌鳢肠道消化酶活性的影响(见表3)
由表3可知,随着GABA添加量的增加,乌鳢消化酶活性逐渐上升,在GABA添加量为60~120 mg/kg时胰蛋白酶活性显著高于其他各组(P<0.05),并在添加量为90 mg/kg时胰蛋白酶活性达到最高;GABA添加量为60~120 mg/kg 时脂肪酶活性显著高于对照组(P<0.05),且在添加量为90 mg/kg时脂肪酶活性达到最高。

表3 GABA对乌鳢肠道消化酶活性的影响
2.3 GABA对乌鳢肠道抗氧化酶活性的影响(见表4)
由表4可知,随着GABA添加量的增加,乌鳢抗氧化酶活性逐渐上升,在GABA 添加量为60~120 mg/kg时,T-SOD活性显著高于对照组(P<0.05),且在GABA添加量为90 mg/kg时达到最高值;在GABA添加量为60~120 mg/kg 时,CAT 活 性 显 著 高 于 对 照 组(P<0.05),同样在添加量为90 mg/kg 时达到最高;在GABA 添加量为60~120 mg/kg 时,GSH-Px 活性显著高于其他各组(P<0.05),并在添加量为90 mg/kg时达到最高。

表4 GABA对乌鳢肠道抗氧化酶活性的影响
3 讨论
3.1 GABA对乌鳢生长性能的影响
作为饲料添加剂,GABA 对水生动物具有明显的促生长作用。Kim 等[15]研究表明,当GABA 添加量在1%时可以有效促进日本比目鱼(Paralichthys oliva⁃ceus)的食欲,同时增强机体生长。徐贺等[16]研究表明,在饲料中添加54 mg/kg 的GABA 时可提高建鲤(Cyprinus carpio var.Jian)的饲料效率和蛋白质效率,说明添加GABA 可以提高建鲤对饲料和蛋白质的利用率,进而促进其机体生长。Fan 等[17]研究表明,当GABA添加量为87.5 mg/kg时饲料效率和特定生长率显著增加,说明GABA 可调节食欲,促进草鱼(Cteno⁃pharyngodon idellus)摄食使其加速生长。Xie 等[14]研究表明,饲料中添加150 mg/kg 的GABA 时可增加南美白对虾的采食量和饲料效率,说明GABA对其生长具有促进作用。同样在对锦鲤(Cyprinus carpio)[18]和翘嘴鳜[19]等鱼类以及肉仔鸡[7]和三元杂交猪[10]等哺乳动物上的研究结果也证明,GABA 在机体营养和健康方面有积极的调控作用。陈秀梅等[20]研究发现,在饲料中添加90 mg/kg 的GABA 一定程度上促进了建鲤的生长,但当添加量在60 mg/kg 时对建鲤生长无影响,不能显著提高建鲤的生长率。可能原因是由于饲料中添加GABA 的量不足而导致促生长作用未能表现。GABA 对不同鱼类生长效果的影响具有一定的差异性,可能与鱼的自身种类、机体体质和其饲料组成以及其生存环境等各种方面都具有一定关系[21];鱼的体质以及其生存环境不同会导致鱼体对需求GA⁃BA 的量产生一定差异[20];不同饲料原料配比对内源GABA 的合成会产生一定影响,导致鱼对饲料中GA⁃BA 的需求量也发生相应改变[22]。这暗示着GABA 的促生长作用可能存在剂量依赖性,具体调控机制需深入研究。
3.2 GABA对乌鳢肠道消化酶活性的影响
肠道作为一种重要的消化器官,在消化食物、吸收营养方面发挥重要的功能[23],同时这一功能的发挥与分泌的胃酸(有胃鱼)、黏液、黏蛋白、胆汁、糖蛋白、黏多糖、各种消化酶等化学物质密切相关,尤其是消化酶活性[24]。消化酶活性是决定某种食物在促进生长、摄食和利用等方面的有效参数,在一定程度上鱼类的消化吸收能力与摄食饲料中营养组分的能力是由其消化酶活性的强弱来体现的,从而对鱼类机体的生长发育产生影响[25-26]。胰蛋白酶作为一种重要的消化酶,是一种动物来源的蛋白水解酶,使蛋白质生成小分子的氨基酸[27]。脂肪酶是一组特异性较低的脂肪水解酶类,能水解多种含长链脂肪酸的甘油酯[28]。本试验条件下,在GABA添加量为60~120 mg/kg时胰蛋白酶和脂肪酶活性显著高于对照组。这表明饲料中外源添加GABA 能一定程度上改善乌鳢肠道的消化能力。但目前尚未有关于GABA 调控鱼类消化能力的其他报道。
3.3 GABA对乌鳢肠道抗氧化酶活性的影响
鱼类生活在水中,因水中环境的复杂多变而易受到氧化应激。肠道作为机体与外部环境进行交流的场所之一,因肠黏膜与肠腔内大量的细菌、病毒、生物毒素及化学毒素广泛接触,极易成为机体遭受威胁的部位[29]。T-SOD、CAT及GSH-Px作为机体抗氧化酶类,其活性的高低可以直接反映出机体的抗氧化水平[30]。SOD作用于超氧自由基,将体内的超氧自由基经催化歧化作用转化为氧与过氧化氢[31],CAT 则负责催化组织器官中的H2O2[32],GSH-Px为含硒半胱氨酸,至少有4 种同工酶,催化还原H2O2和有机氢过氧化物[33]。本试验条件下,在GABA 添加量为60~120 mg/kg 时TSOD、CAT 及GSH-Px 活性显著高于对照组。这表明GABA 可以对乌鳢肠道的抗氧化能力具有一定的改善作用。类似的报道有很多,Fan 等[17]对草鱼的研究结果表明GABA 可显著提高其机体内SOD 活性。王斌等[34]对栉孔扇贝(Chlamys Farreri)上的研究也证明GABA 能显著提高其机体内CAT 的活性。而且还有研究报道GABA 对鱼类抗缺氧[13,35-36]、运输应激[37]、急性低温应激[38]等能力也有一定的改善作用。以上研究均表明GABA 对提高鱼类的抗应激能力有重要的作用,但其具体机制尚未深入研究。
4 结论
综上所述,当饲料中添加88.54~102.73 mg/kg 的GABA 时在一定程度上能够有效提高乌鳢生长、肠道消化酶和抗氧化酶活性。
