七星瓢虫触角转录组及嗅觉相关基因分析
2020-08-06杨雪娇江宇航唐启河张棋麟林连兵陈均远
杨雪娇, 张 蔓, 江宇航, 唐启河, 张棋麟,*, 林连兵, 陈均远
(1. 昆明理工大学生命科学与技术学院, 昆明 650500; 2. 云南省高校饲用抗生素替代技术工程研究中心, 昆明 650500; 3. 中国科学院南京地质古生物研究所, 南京 210008; 4. 南京大学生命科学学院, 医药生物技术国家重点实验室, 南京 210023)
嗅觉是昆虫食物选择、求偶、交配及产卵等各种行为的基础,对其种群生存、繁衍有重要作用(Fieldetal., 2001; Sato and Touhara, 2009; Zhangetal., 2015; Gadenneetal., 2016)。昆虫触角上着生许多嗅觉感器且有各种嗅觉蛋白,不同的感器和嗅觉蛋白可识别不同的气味物质进而调控昆虫的行为活动(Brownetal., 2013; 郑海霞等, 2018)。目前,昆虫触角中研究最为广泛的嗅觉相关蛋白主要包括:气味结合蛋白(odorant binding protein, OBP)、化学感受蛋白(chemosensory protein, CSP)、嗅觉/气味受体(olfactory odorant receptor, OR)、离子型受体(ionotropie receptor, IR)、味觉受体(gustatory receptor, GR)和昆虫感觉神经元膜蛋白(sensory neuron membrane protein, SNMP)六大类(Leal, 2013; Zhangetal., 2016; Wang Jetal., 2017)。当前,基于昆虫触角转录组学挖掘嗅觉相关基因已经被广泛开展。Zhang等(2016)从美国白蛾Hyphantriacunea触角转录组中鉴定了124个嗅觉相关基因,主要包括OR, OBP和CSP基因等,并分析了OBP和CSP基因的表达模式。Jia等(2016)采用了相似的研究策略鉴定了桃蛀螟Conogethespunctiferalis雌雄成虫触角中的嗅觉相关基因,并对呈性别差异表达的嗅觉相关基因进行了鉴定。有研究从云南同域生境下云南松毛虫Dendrolimushoui和思茅松毛虫D.kikuchii触角转录组中鉴定了一批嗅觉相关基因,进行了属内和目间比较和基因表达水平分析,探讨了基因功能的潜在相似性(Zhangetal., 2014)。然而,迄今为止未见从嗅觉相关基因的角度探讨其在不同食性昆虫中的差异。昆虫触角嗅觉相关基因是否是其食性分化的主要因素,昆虫嗅觉相关基因数量及不同食性昆虫间基因数量差异对了解昆虫间食性分化能提供什么信息等,都是有待探讨的问题。
瓢虫是鞘翅目(Coleoptera)瓢虫科(Coccinellidae)昆虫的总称,涵盖90个属超过1 000个种(Escalonaetal., 2017),其中七星瓢虫Coccinellaseptempunctata隶属瓢虫亚科(Coccinellinae),是以蚜虫为主食的捕食性昆虫(Van der Werfetal., 2000)。瓢虫科昆虫已经被广泛地用于科学研究,主要集中在食性可塑性的发育、系统发育、种群遗传、鞘翅颜色多样化和捕食性(天敌)瓢虫对其他害虫的生物防治等(Escalonaetal., 2017; Matsubayashietal., 2017; Saitoetal., 2017; Tangetal., 2017)。值得注意的是,科内高度的食性分化是瓢虫科的一大特征(Escalonaetal., 2017),大部分瓢虫都是以捕食蚜虫(如棉蚜Aphisgossypii, 大豆蚜A.glycines, 萝卜蚜Lipaphiserysimi等)、蚧壳虫、鳞翅目幼虫、叶螨类和真菌孢子为主的捕食性瓢虫。另外,还有少部分种类为植食性瓢虫,主要包括草食性的食植瓢虫亚科(Epilachninae),以花粉和蔬菜等为食,多数为农业害虫,如茄二十八星瓢虫Henosepilachnaviginyioctopunctata等(庞虹, 2002; Zhangetal., 2018)。
为了探索昆虫触角嗅觉相关基因在昆虫食性分化中的地位及角色,本研究以捕食性七星瓢虫为研究对象,运用Illumina HiSeq高通量测序技术对其触角转录组进行测序,采用生物信息学方法对其基因功能注释,并联合我们以前测序的植食性茄二十八星瓢虫转录组鉴定并比较各类型嗅觉相关基因占嗅觉相关基因总数的比例。研究结果将加深对瓢虫食性分化分子基础的了解,也为瓢虫科天敌利用和害虫防治提供理论依据。
1 材料与方法
1.1 试虫
七星瓢虫雌雄成虫采自广西壮族自治区北海市(海拔10 m, 109°08′E, 21°25′N)。为了避免局部环境的差异,七星瓢虫分别采集于不同的3个点,每个点相距至少30 km以上。因七星瓢虫成虫特征明显,种类鉴定简单易行,野外鉴定带回实验室后,选取活动能力较强的七星瓢虫成虫200头,切下触角,经液氮速冻,后保存于-80℃超低温冰箱(Thermo Scientific, 美国)待用。
1.2 总RNA提取
采用RNeasy Mini Kit(天根, 北京)试剂盒分别提取供试雌雄七星瓢虫成虫各100头触角总RNA。经NanoDrop® ND1000(Thermo Scientific, 美国)核酸浓度测定仪检测RNA的浓度和纯度,并采用1.5%的琼脂糖凝胶电泳和Agilent 2100(安捷伦科技, 美国)精确检测RNA的完整性。等量混合合格的雌雄七星瓢虫触角总RNA为一个混池,用于后续转录组测序。
1.3 cDNA文库的构建和测序
合格的七星瓢虫触角总RNA使用DNase I(Qiagen, 德国)消化残余基因组DNA后,按照Preparing Samples for Sequencing of mRNA试剂盒说明书(Illumina, 美国)构建测序文库。具体步骤:用带有Oligo(dT)的磁珠富集总RNA池中的mRNA;加入打断试剂后于Thermomixer C(Eppendorf, 德国)中适温将mRNA打断成短片段(200~400 bp),以打断后的mRNA短片段为模板合成cDNA第1链,随后在DNA polymerase I体系下,以dNTPs 为原料合成cDNA第2条链,并使用试剂盒纯化回收、粘性末端修复、cDNA的3′末端加上碱基A并连接接头,然后进行片段大小选择,最后进行PCR扩增;构建好的文库用Agilent 2100 Bioanalyzer(安捷伦科技, 美国)和ABI StepOnePlus Real-Time PCR System(Applied Biosystems, 美国)质检,合格后使用HiSeq 4000高通量测序平台(Illumina, 美国)的PE150 bp双向测序技术进行测序。
1.4 转录组测序数据组装及完整性评估
利用Trimmomatic v0.39(Bolgeretal., 2014)软件对测序得到的原始数据(raw data)进行过滤,去除接头序列以及低质量reads,获得高质量的数据(clean data),具体步骤如下: (1)去除包含接头的reads(接头污染);(2)去除未知碱基N含量大于5%的reads;(3)去除低质量的reads(质量值低于15的碱基占该reads总碱基数的比例大于30%的reads为低质量的reads)。利用Trinity软件对clean data进行序列拼接与组装。然后,去除长度小于150 bp的序列,并通过CD-HIT软件包(http:∥www.bioinformatics.org/cd-hit/)中的CD-HIT-EST子程序对unigenes以95%的相似性进行聚类,并选取最长的序列为每一个聚类簇的代表,以去除unigenes的冗余,最终获得高质量的七星瓢虫触角unigenes集。使用BUSCO(Benchmarking Universal Single Copy Orthologs)v3软件于默认参数下,将最终组装获得的unigenes通过与insecta_odb9数据库(https:∥busco.ezlab.org/)1 658个昆虫进化保守的直系同源基因进行比较,检测每个unigene的完整性,以评估转录组组装质量。
1.5 转录组数据分析及嗅觉相关基因鉴定
将1.4节所得unigenes,使用本地Blastx软件分别与NR, NT, COG, Swiss-Prot和KEGG等数据库进行比对(E值<1×10-5),获得相应的注释信息。根据NR注释信息获得基因功能信息,然后提取每个基因相应的GO注释信息,并基于GO注释把基因划分到生物学过程、细胞组分和分子功能等三亚类。利用COG数据库注释信息对直系同源蛋白家族进行功能分类。KEGG注释信息用以呈现基因参与的代谢通路。茄二十八星瓢虫高质量的unigenes集及相关基因功能注释信息从我们以前的数据集相关研究中获取(Zhangetal., 2018)。最后,采用以往文献使用的方法(Jiaetal., 2016; Zhangetal., 2016; 郑海霞等, 2018),即根据基因注释信息鉴定嗅觉相关基因。
2 结果
2.1 七星瓢虫触角转录组序列分析与组装
七星瓢虫触角转录组测序获得10.73 Gb原始数据。质控后获得70 291 132条共10.54 Gb高质量clean reads数据,Q20和Q30值碱基比例分别为97.57%和93.69%。利用Trinity软件对获得的高质量reads进行组装,共获得46 687条contigs,总长度为41 887 759 bp,平均长度为897 bp,N50的长度为1 871 bp,N70的长度为1 061 bp,N90的长度为311 bp。对contigs进一步进行聚类和去冗余,得到31 775条unigenes,总长度为34 872 097 bp,平均长1 097 bp,N50的长度为2 010 bp;GC含量为38.99%。unigenes长度分布(图1)显示,在长度1 kb以上的unigenes为13 107条,占unigenes总数的41.25%。BUSCO分析结果显示83.22%的unigenes为完整基因,7.13%是部分片段,剩下的unigenes没有比对到任何序列。为了使相关学者使用方便,避免前期数据组装的不便,unigenes集已释放到公共存储数据库Figshare(https:∥doi.org/10.6084/m9.figshare.10259660.v1)供直接下载使用。
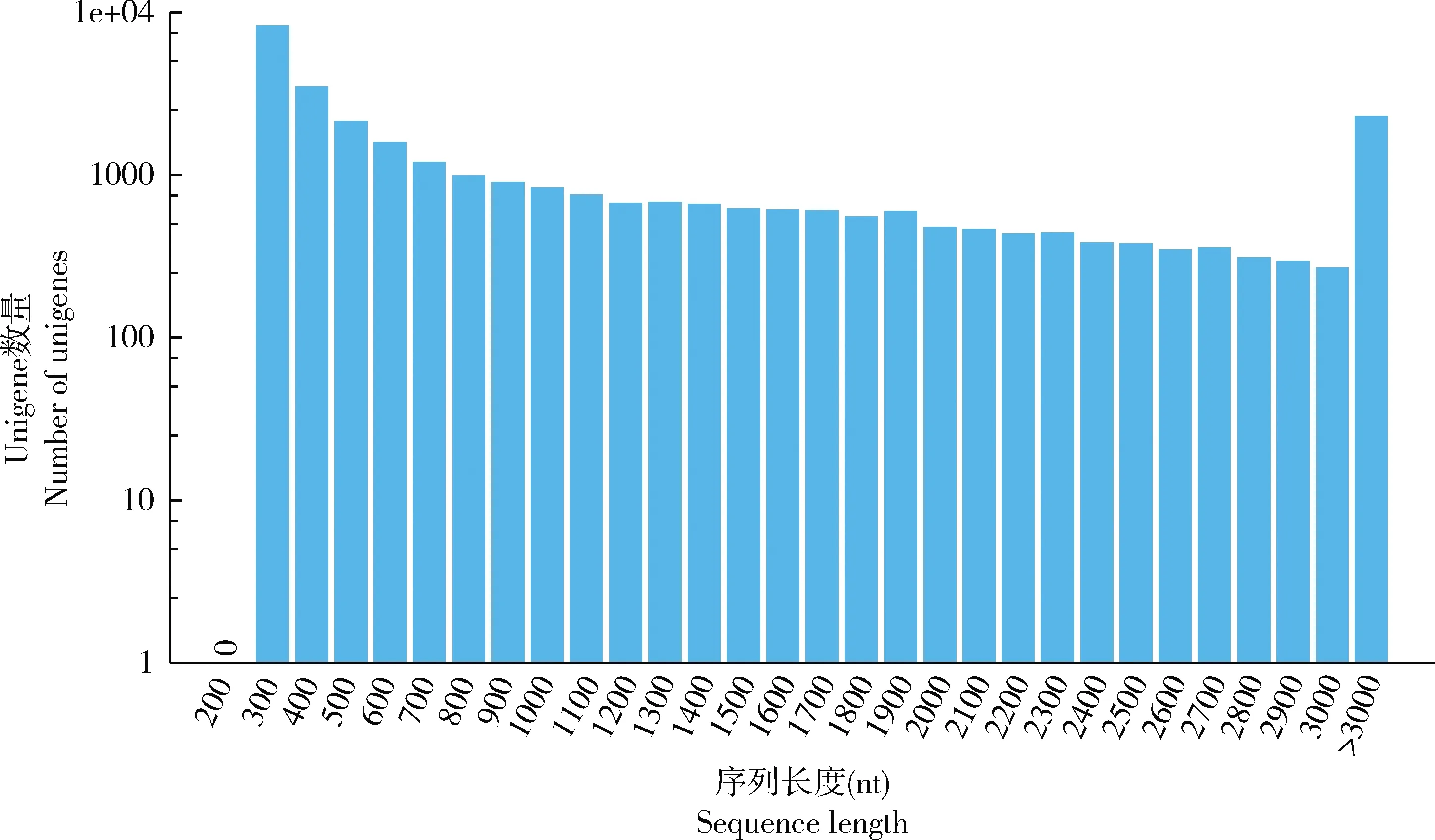
图1 七星瓢虫触角转录组unigenes长度分布Fig. 1 Length distribution of all assembled unigenes in the antennal transcriptome of Coccinella septempunctata
2.2 七星瓢虫触角转录组基因功能注释
七星瓢虫触角转录组中,共有22 149条(69.71%) unigenes获得注释。其中NR数据库注释到的unigenes数目最多,为20 539条(64.64%);GO数据库注释的unigenes最少,为4 863条(15.30%);NT数据库注释到7 576条(23.84%);Swiss-Prot数据库注释到16 862条(53.07%);KEGG数据库注释到16 445条(51.75%);COG数据库注释到9 342条(29.40%)。
基于NR注释信息,七星瓢虫触角转录组中共注释到20 539条unigenes,其中与赤拟谷盗Triboliumcastaneum的相似基因最多,所占比例为62.70%,其后依次为中欧山松大小蠹Dendroctonusponderosae(7.79%)、丽蝇蛹集金小蜂Nasoniavitripennis(1.21%)、豌豆蚜Acyrthosiphonpisum(1.05%),其他物种占27.26%(图2)。基于GO数据库对七星瓢虫触角转录组进行功能分类统计(图3),共得到22 020条功能注释信息,分布于4 863条unigenes,生物学过程(9 841条unigenes)中代谢过程基因最多(2 373条unigenes),其次是细胞进程(2 325条unigenes)和单生物进程(1 695条unigenes),其余的均低于1 000条unigenes;细胞组分(6 406条unigenes)中细胞部分和细胞最多,均为1 454条unigenes,其次是细胞器(931条unigenes)、膜(800条unigenes)、大分子复合物(641条unigenes)、膜部分(529条unigenes),其余均低于500条unigenes;分子功能(5 773条unigenes)中催化活性有2 430条unigenes, 结合有2 350条unigenes,其余均低于500条unigenes。
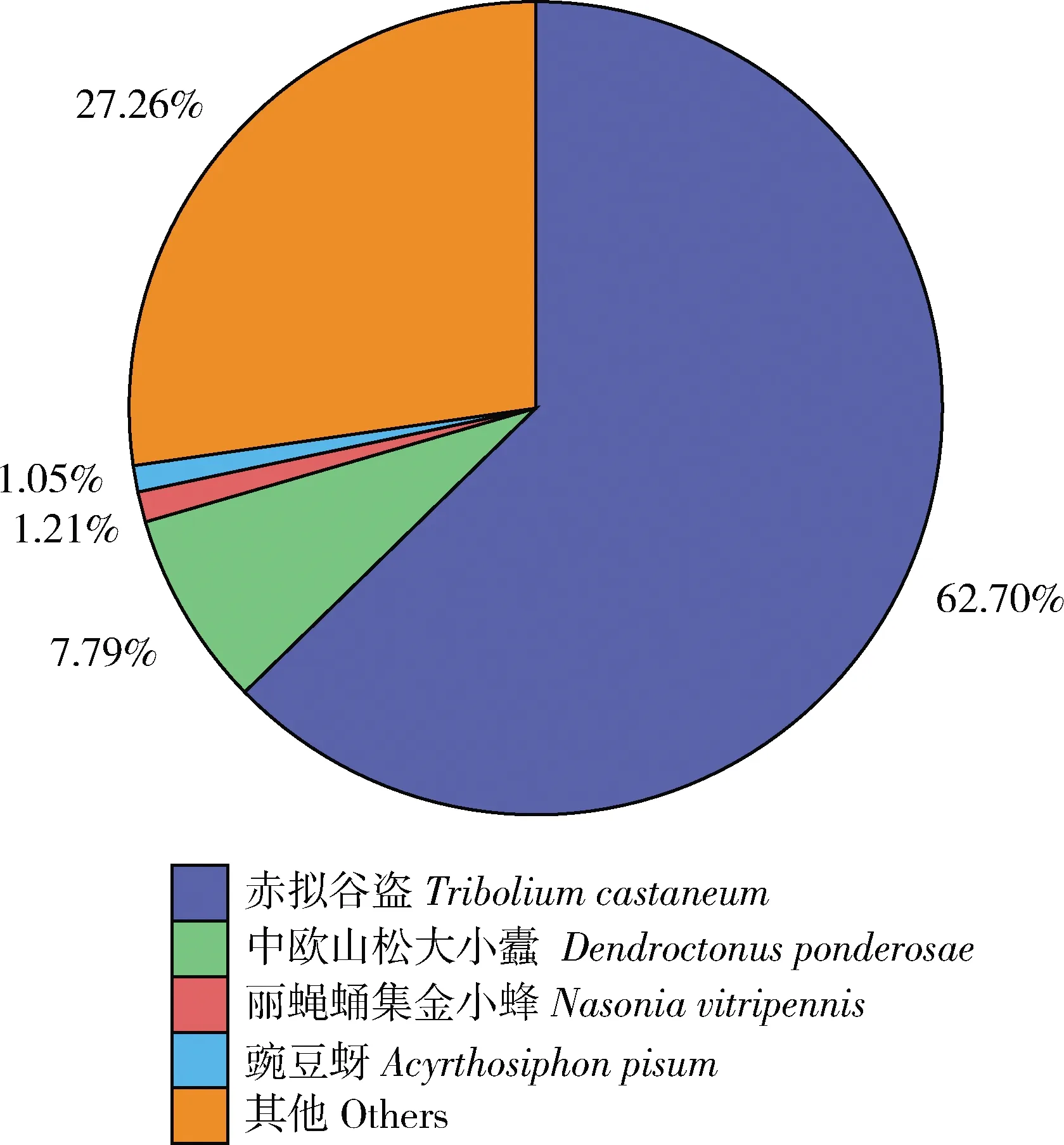
图2 七星瓢虫触角unigenes在NR数据库中的物种分布Fig. 2 Species distribution of antennal unigenes of Coccinella septempunctata in the NR database
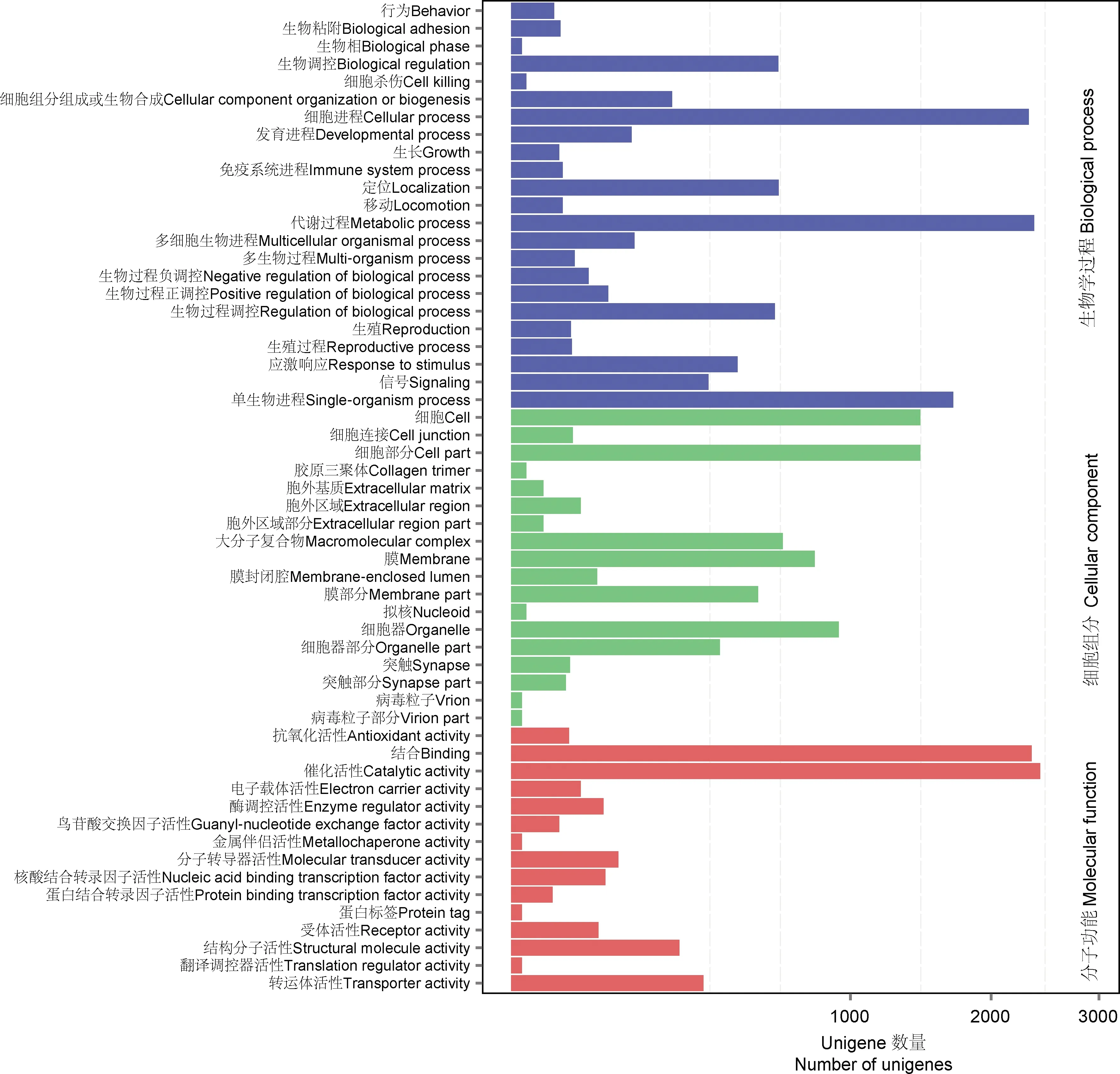
图3 七星瓢虫触角unigenes GO功能注释Fig. 3 GO function annotation of antennal unigenes of Coccinella septempunctata
在COG数据库中,9 342条unigenes被分为25个大类,共包含14 906条注释信息(图4)。其中数量最多的是仅一般功能预测类(2 841条, 19.06%),其余依次为翻译、核糖体结构和生物合成类(1 197条, 8.03%),复制、重组和修复类(1 136条, 7.62%)等。根据KEGG数据库对七星瓢虫触角转录组的注释信息,16 445条unigenes注释到31 645个代谢通路(表1),有9个代谢通路包含300条以上的unigenes:代谢通路(649, 3.95%)、癌症中通路(538, 3.27%)、内吞作用(393, 2.39%)、艾巴氏病毒感染(382, 2.32%)、嘌呤代谢(370, 2.25%)、噬菌体(368, 2.24%)、癌症中的蛋白多糖(362, 2.20%)、RNA转运(328, 1.99%)、肌动蛋白骨架调节(322, 1.96%);包含150~300条unigenes的代谢通路有18个;其余代谢通路所含unigenes数量均低于150条。
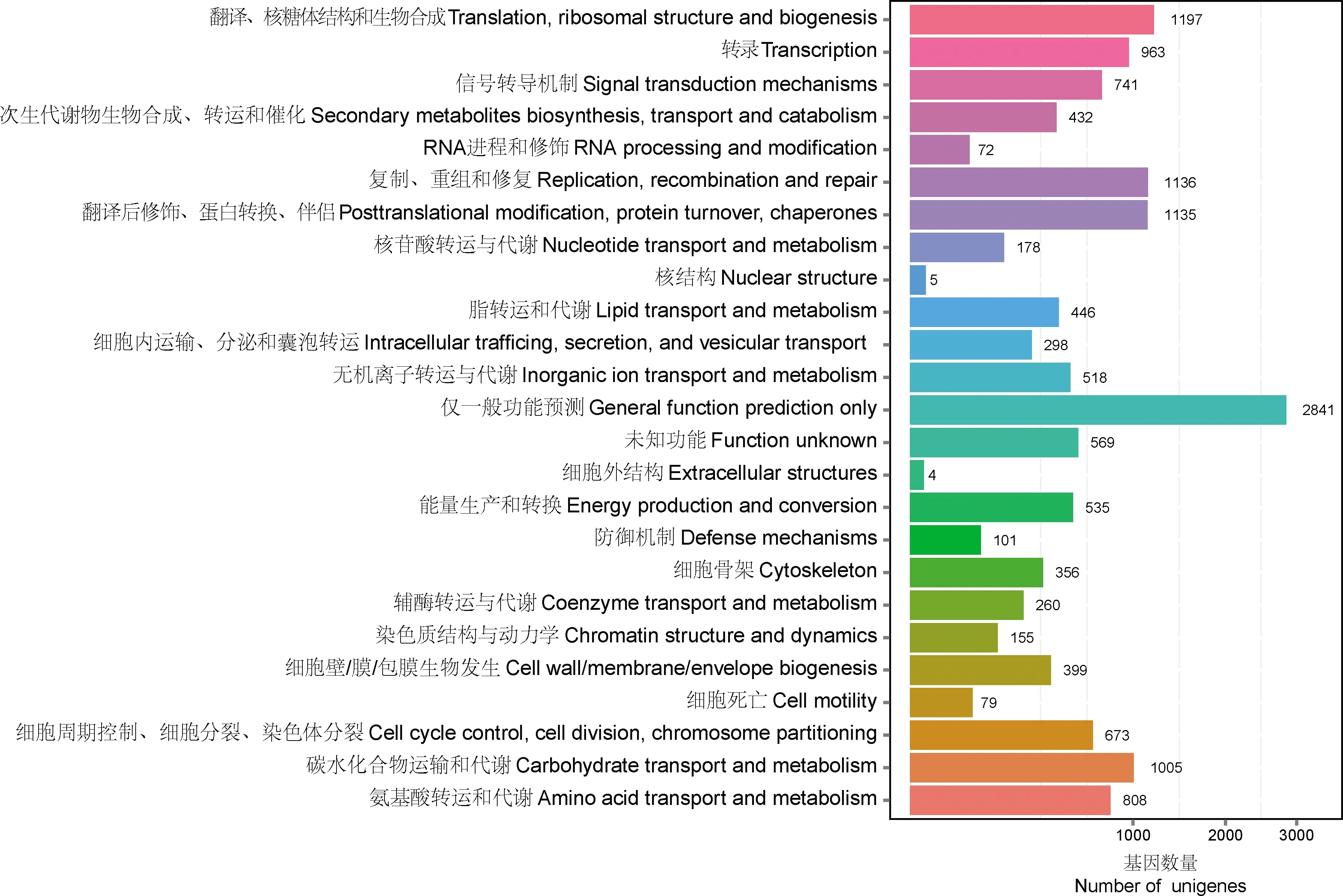
图4 七星瓢虫触角unigenes COG功能分类Fig. 4 COG function classification of antennal unigenes of Coccinella septempunctata

表1 七星瓢虫触角转录组KEGG通路分析Table 1 KEGG pathways in the antennal transcriptome of Coccinella septempunctata

续表1 Table 1 continued
2.3 七星瓢虫触角转录组嗅觉相关基因分析
基于六大公共数据库得到的unigenes注释信息,27个嗅觉相关基因在七星瓢虫触角转录组中被鉴定(表2): 1个气味结合蛋白(OBP)基因(3.70%)、13个化学感受蛋白(CSP)基因(48.15%)、4个气味受体(odorant receptor, OR)基因(14.48%)、7个味觉受体(GR)基因(25.93%)和2个感觉神经元膜蛋白(SNMP)基因(7.41%)。另外,在茄二十八星瓢虫转录组中鉴定到38个嗅觉相关基因,包括5个OBP基因(13.16%), 17个CSP基因(44.74%), 4个OR基因(10.53%), 1个IR基因(2.63%), 5个GR基因(13.16%)和6个SNMP基因 (15.79%)。比较发现,茄二十八星瓢虫转录组各类型嗅觉相关基因中,OBP基因比例(13.16%)高于七星瓢虫触角转录组的(3.70%),而七星瓢虫触角转录组中GR基因的比例(25.93%)高于茄二十八星瓢虫(13.16%)。
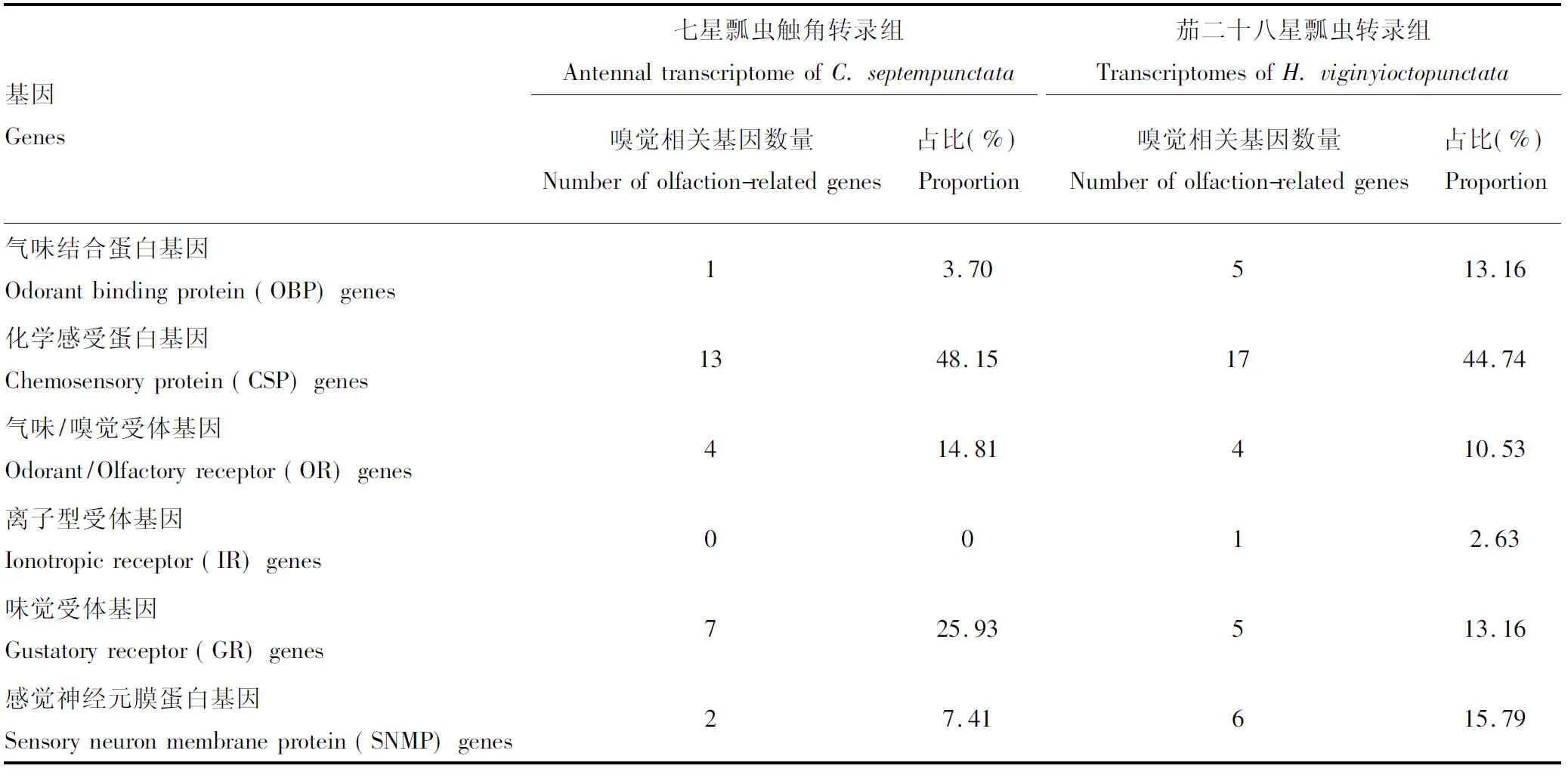
表2 七星瓢虫和茄二十八星瓢虫转录组嗅觉相关基因统计Table 2 Olfaction-related genes in the transcriptomes of Coccinella septempunctata and Henosepilachna viginyioctopunctata
3 讨论
迄今为止,嗅觉相关基因已经在20余种鞘翅目昆虫中被研究,但是在瓢虫中还鲜有报道。此外,通过与其他组装的45种节肢动物转录组BUSCO结果(Theissingeretal., 2016)比较,本研究获得的七星瓢虫触角转录组中,完整的unigenes占比为83.22%,证实了我们得到高质量的七星瓢虫触角转录组测序及组装结果。采用NR蛋白数据库比对获得的unigenes最佳比对物种信息注释分类显示,最高相似性的物种为赤拟谷盗和中欧山松大小蠹(图2),这两种甲虫作为最佳比对物种的unigenes占据了所有注释基因的约70%,这主要归因于其与七星瓢虫一道作为鞘翅目昆虫,具有相对完整的全基因组序列,且具有丰富的基因组注释信息;尤其赤拟谷盗为著名的模式动物,拥有鞘翅目昆虫中最全面的基因组学信息(Tribolium Genome Sequencing Consortium, 2008)。已有大量研究结果表明,注释到的基因均与基因组信息已知且亲缘关系较近的物种相似性序列最多(郑海霞等, 2018; 杜迎刚等, 2019)。GO分类表明,七星瓢虫触角转录组基因划分为生物学过程、细胞组分与分子功能3个亚类,其中参与细胞进程、代谢过程、细胞和细胞部分、结合、催化活性的基因功能注释较多,这与大多数鞘翅目昆虫触角转录组中报道的(Guetal., 2015; 郑海霞等, 2018)较为一致,表明这些基因是维持甲虫触角生物学功能的基础。当然,不同文献报道的不同鞘翅目昆虫触角转录组中基因功能注释的GO数目不同,这可能是由不同的昆虫种类、生存环境、测序手段和分析参数等不同所导致的。此外,包括KEGG注释信息,69.71%的unigenes在所有六大数据库中得以注释,这个结果表明有超过30%的七星瓢虫触角转录组unigenes没有任何注释信息,可见仍有一大批未知基因功能的新基因有待鉴定,其中也可能包含许多潜在的新的嗅觉相关基因。此外,unigenes主要涉及代谢、免疫防御、核糖体和蛋白过程等代谢通路,表明了外源物质防御、免疫响应、蛋白质的合成及代谢在七星瓢虫嗅觉行为活动中的重要性。
本研究首次以两种食性分化程度最高的瓢虫转录组为对象,从七星瓢虫中挖掘到27个嗅觉相关基因(表2),并通过以往获得的茄二十八星瓢虫转录组数据(Zhangetal., 2018),从中鉴定到38个嗅觉相关基因,主要包括OBP, CSP, OR, IR, GR和SNMP基因。此外,已有研究发现捕食性的异色瓢虫Harmoniaaxyridis触角转录组中至少存在57个嗅觉相关基因(包括30个OBP基因和27个CSP基因)(张伟, 2018)。可见,嗅觉相关基因数量差异在捕食-捕食瓢虫间比捕食-植食瓢虫间大。另一方面,不同食性的瓢虫间,嗅觉相关基因数量差异不及瓢虫与其他目昆虫间嗅觉相关基因数量差异大。例如鞘翅目红脂大小蠹Dendroctonusvalens、星天牛Anoplophorachinensis、光肩星天牛Anoplophoraglabripennis和红棕象甲Rhynchophorusferrugineus触角转录组中六大类嗅觉相关基因总数分别为60, 151, 108和157个(Guetal., 2015; Antonyetal., 2016; Huetal., 2016b; Wang Jetal., 2017);鳞翅目美国白蛾、草地螟Loxostegesticticalis和沙棘木蠹蛾Eogystiahippophaecolus触角转录组中分别为124, 131和137个(Huetal., 2016a; Zhangetal., 2016; Weietal., 2017);半翅目柑橘木虱Diaphorinacitri、枯蝉Subpsaltriayangi和黑肩绿盲蝽Cyrtorhinuslividipennis触角转录组中分别为126, 77和57个(Wuetal., 2016; Qietal., 2018; Wangetal., 2018);双翅目南亚实蝇Bactroceratau、黑带食蚜蝇Episyrphusbalteatus和大灰优食蚜蝇Eupeodescorollae、柑橘大实蝇Bactroceraminax触角转录组中分别为104, 154, 134和127个(Wang Betal., 2017; 杜迎刚等, 2019; Wuetal., 2019)。因此,我们提出昆虫触角嗅觉相关基因的绝对数量可能不是昆虫食性分化一个最重要的因素。如两种食性瓢虫、不同食性蝇类间等食性差异很大,但嗅觉相关基因数量差异较小;相反,有的昆虫类群(如柑橘木虱与枯蝉和黑肩绿盲蝽、红脂大小蠹与天牛和象甲等)食性相似,但嗅觉相关基因数量差异较大。另外,上述例子中两种(星天牛和光肩星天牛)同隶属Anoplophora属食性相似的天牛触角转录组中嗅觉相关基因数目也有一定差异(151vs108),而与其他鞘翅目昆虫又较为一致,这两种天牛嗅觉相关基因数量差异,可能是除食性外的其他环境因子引起的。
在七星瓢虫触角和茄二十八星瓢虫转录组中,CSP基因数量均占据了嗅觉相关基因数目的绝大多数(40%以上)(表2)。不同昆虫间CSP基因所占嗅觉相关基因比重不同,如星天牛(10.60%)、红脂大小蠹(10%)和美国白蛾(13.71%)(Guetal., 2015; Zhangetal., 2016; Wang Jetal., 2017)。另外,OBP基因在七星瓢虫触角(3.70%)和茄二十八星瓢虫(13.16%)转录组中所占比例差异较大(表2)。其他嗅觉相关基因类型也是如此,可见不同昆虫类群间,嗅觉相关基因组成比重不尽相同。另外,OBPs是嗅觉发生进程中的一个关键环节,因为它们通过昆虫感觉淋巴将气味从外部环境传送到ORs (Leal, 2004, 2013)。在七星瓢虫触角转录组中,未有IR基因被鉴定到,茄二十八星瓢虫中鉴定到了1个IR基因(表2)。IR基因家族包含两大类:一类是与嗅觉有关的保守的触角IRs;另一类是在包括味觉器官在内的其他组织中表达的具有物种特异性的IRs (Crosetetal., 2010)。本研究中,七星瓢虫触角转录组未发现IR基因,再者茄二十八星瓢虫因为是其不同发育阶段整个虫体的转录组混池组装结果,故这些证据表明瓢虫中鉴定到的IR基因极有可能属于第2种类型。然而,还需要通过与其他昆虫两种类型的IR基因联合构建系统发育树和进行不同组织基因表达分析最终确认。GR基因在昆虫中主要在味觉器官的味觉感受神经元中表达,作为糖受体、热传感器、二氧化碳和苦味受体,参与糖、苦味化合物和其他昆虫信息素的检测 (Nietal., 2013; Guetal., 2015)。本研究中发现七星瓢虫触角所含GR基因的比例(25.93%)是茄二十八星瓢虫中的近2倍(13.16%)(表2),表明了七星瓢虫在捕食过程中触角有更强的代谢能力,对糖及苦味化合物有更灵敏的嗅觉,对其他昆虫(尤其捕食对象)有更快速的响应。SNMPs的功能在昆虫中还鲜有报道,故仍需更多的实验研究。
本研究测序、组装及注释了七星瓢虫触角转录组,并结合茄二十八星瓢虫不同发育阶段测序数据混池组装的转录组,在两种食性的瓢虫中鉴定了六大类昆虫嗅觉相关基因,并基于每类基因所占鉴定出的嗅觉相关基因总数比例,比较了它们在两种食性瓢虫间的差异。值得注意的是,因为测序深度及测序组织的不同,直接比较每种嗅觉相关基因类型在两种瓢虫中的绝对数目是欠妥的,故我们将其转换成统一测序背景下,不同类型嗅觉相关基因在总数中所占的比重/比例进行比较。本研究获得了七星瓢虫高质量的触角转录组及两种瓢虫中的一批嗅觉相关基因,但这些基因的组织表达模式和生理生态功能仍有待进一步研究。今后,可利用实时荧光定量PCR技术,对瓢虫嗅觉相关基因组织特异表达情况确认,以辅助明确其生物学功能;结合不同层次的组学测序结果,利用正反向遗传学手段,过表达或沉默关键嗅觉相关基因,进一步明确嗅觉相关基因在瓢虫食性分化中的作用。
