长江上游小麦新品种(系)品质分析
2020-07-31董亚超戴常军李式昭朱华忠
伍 玲,董亚超,戴常军,李式昭,朱华忠
(1.四川省农业科学院作物研究所,四川成都 610066;2.中国农业科学院作物科学研究所,北京 100081;3.农业农村部谷物及制品质量监督检验测试中心(哈尔滨),黑龙江哈尔滨 150086)
长江上游冬麦区属于我国的中筋和弱筋小麦生产区域,小麦品质改良经历了不重视质量—重视蛋白质含量—重视强筋—重视中筋和弱筋小麦品质全面提升的过程。20世纪50-80年代,四川省的小麦育种以提高产量为第一目标,几乎没有对加工品质进行选择;20世纪90年代后,该区域的小麦品质改良注重了蛋白质含量、湿面筋含量和高分子量麦谷蛋白亚基的改良和选择[1];而同时期云南、贵州等省与四川的情况类似。到2010年前后,四川省小麦新品系的平均蛋白质含量和湿面筋含量已与北方麦区基本相当,含优质高分子量麦谷蛋白亚基的品种大大增加,5+10、7+8、17+18等亚基在新品种中并不鲜见。但是该区域收获期降雨较多,穗发芽严重,降落值较低,严重影响小麦品质。且该区域小麦品种的面团稳定时间一直较短[2]。晏本菊等[3]分析了四川省“九五”和“十五”期间育成的小麦品种的蛋白质组分,认为对该区域的小麦品质改良,在提高总蛋白质含量的同时,需要提高谷蛋白含量。2012年以后,却鲜见有关该区小麦品质的报道。
随着小麦基因组学的发展,很多重要性状的基因已被克隆,但育种家对这些已克隆基因在育成品种中的基因型了解并不多。目前这些基因的SNP差异,可用KASP(竞争性等位基因特异性PCR,Kompetitive allele-specific PCR)标记进行检测。KASP标记是对等位基因的差异位点(SNP和InDel)进行竞争性特异扩增,需要三条引物;其中两条正向竞争性引物的5′端有与荧光基团HEX和FAM互补配对的碱基序列,而这两条正向引物的其余碱基序列仅在3′端的SNP和InDel处有差异;一条反向共同保守区引物。PCR反应体系中含有荧光基团和猝灭基团修饰的通用序列(Master Mix由LGC公司提供);因此,正向引物可以特异性地结合基因型与其匹配的DNA模板,两条正向引物可以发出两种颜色不同的光,若模板链中该位点为纯合,则发出单一的、与其匹配的荧光,若为杂合,则可同时发出两种荧光。PCR产物可用酶标仪FAM VIC ROM光束扫描,并使用Kluster Caller软件进行数据分析;或直接使用实时荧光定量PCR仪检测。目前KASP技术已被广泛应用于小麦、水稻和玉米等作物的分子标记检测[4-6],Rasheed等[7]开发了小麦已克隆基因的KASP标记,并用于小麦育种材料的基因型检测,但未见对长江上游品种KASP标记检测的报道。
2012-2018长江上游区域试验包括109个小麦品种(系),川麦42为长江上游麦区国家级品种试验的对照。本研究拟分析这7年区试的品质数据,以明确长江上游小麦品种的品质现状,同时用KASP标记分析这些品种相关位点的基因型,为明确下一步品质育种目标提供思路。
1 材料方法
1.1 供试材料及品质数据来源与品质分类方法
供试材料为国家小麦区域试验长江上游组在2012-2018参试的品种(系)109个,其中,参试一年的品种(系)94个,参试二年的品种(系)15个;以川麦42为对照。
本文所用品质数据均来源于2012-2018国家小麦区域试验长江上游组的试验总结。2017年仅一组试验,其余年份均设置A、B两组试验,共有13组试验。109个品种(系)及其系谱等见附表1。成都、绵阳、内江、永川、昆明、贵阳、勉县和十堰(2012和2013年为襄阳)8试点为区试品质分析取样点;2016年品质分析取样点为7个(十堰试点未送样),其余年份取样点均为8个。
区试分别采用NY/T1094.2-2006《小麦实验室制粉》、GB5498-1985《粮食、油料检验容重测定法》、GB10361-2008《小麦及其面粉降落数值的测定》、NY/T3-1982《粗蛋白质测定法(半微量凯氏法)》、GB/T5506.2-2008《小麦粉面筋含量仪器法测定湿面筋》、GB/14614-2006《小麦粉面团的物理特性吸水量和流变学特性的测定粉质仪法》等方法进行制粉和容重、粗蛋白质含量、湿面筋含量、面团稳定时间和吸水量等测定。
8个品质取样点的样品首先测定降落值,降落值低于200 s的样品被视为芽麦而被剔除;相同品种(系)的样品混合后用于测定粗蛋白质含量、湿面筋含量、面团稳定时间和吸水量。混合点率=样品混合点数/全部取样点数。
根据小麦分级标准[8]将参试材料分为一到五级及等外六个级别;根据品种审定标准[9]将参试材料分为糯麦和非糯麦,非糯麦又分为强筋、中强筋、中筋和弱筋四类。
1.2 KASP标记分析
采用2个高分子量麦谷蛋白亚基(high-molecular-weight glutenin subunit,HMW-GS)基因、3个低分子量麦谷蛋白亚基(low-molecular-weight glutenin subunit,LMW-GS)基因、2个穗发芽抗性基因、9个粒重及籽粒大小基因、3个籽粒硬度基因和1个小麦-黑麦1BL·1RS易位的KASP标记对109个品种(系)进行基因型检测。所用标记见表1,各标记的引物序列等见表2。

表2 KASP标记的引物详情
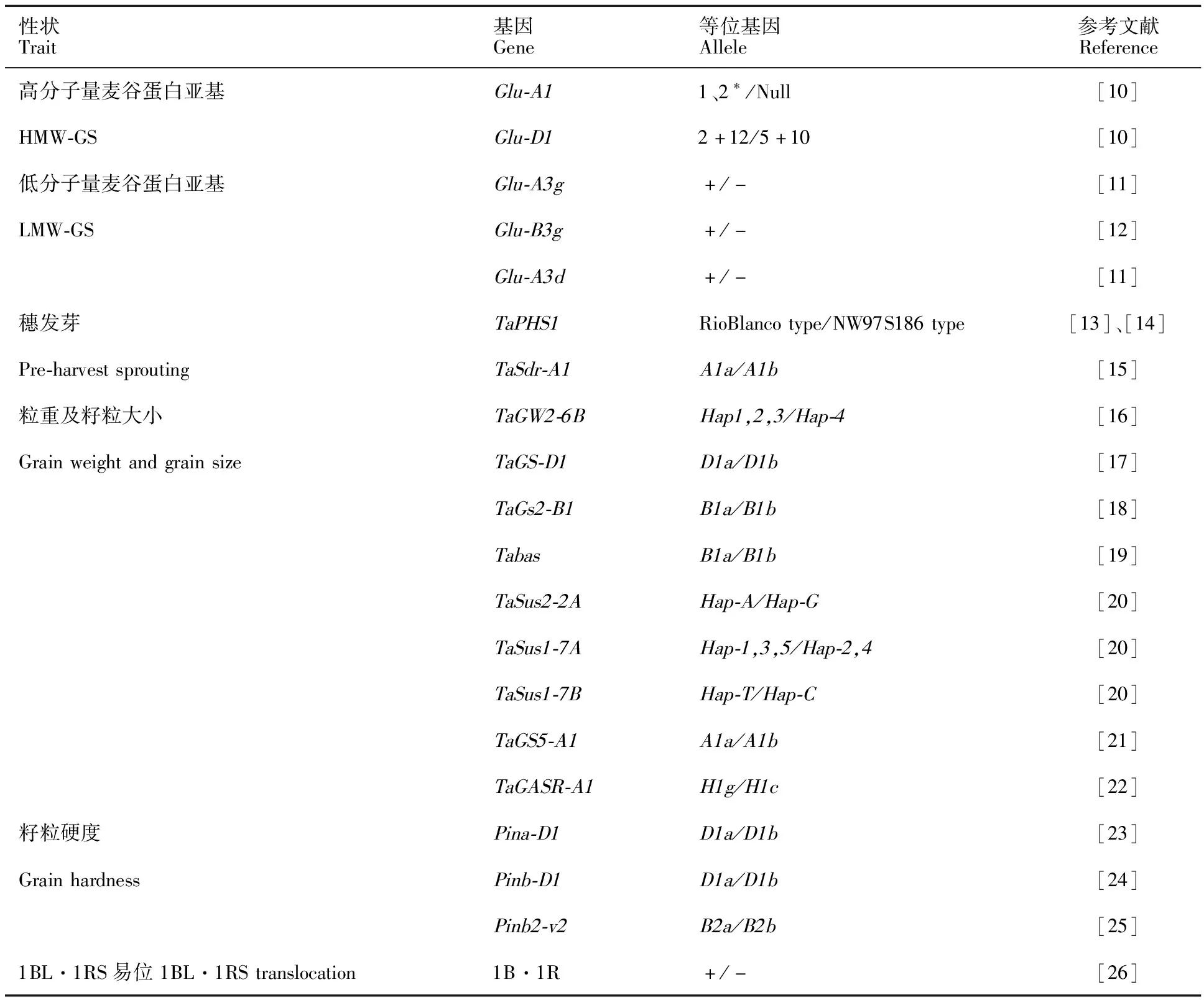
表1 用于分析的KASP标记
每个材料取3~4粒种子,发芽后取2~3 cm的新鲜叶片,采用PVP-40法提取样品DNA,得到DNA浓度为100 ng·μL-1的模板溶液。KASP标记PCR扩增体系和扩增程序同Rasheed等[7]方法,Master Mix和MgCl2购买自LGC公司,Primer Mix含12%HEX引物、12%FAM引物、30%的共同反向引物、46%的ddH2O,由华大基因公司合成。用酶标仪(BiOTek,SYNERGY/H1 microplate reader)的FAM VIC ROM光束扫描PCR扩增产物,用Kluster Caller分型软件对扫描数据进行分型检测。
2 结果与分析
2.1 试验品种(系)的混合点率及两个穗发芽抗性位点的基因型分析
在供试材料中,试验一年的品种(系)94个,得到混合样94个,试验两年的品种(系)15个,得到混合样30个,对照品种1个,得到混合样13个,合计137个样品。137个样品的混合点率不尽相同(表3),其平均混合点率为0.662,多数样品来自4~7个取样点,说明长江上游麦区品种(系)较易发生穗发芽。137个样品中,混合点率为1的仅有9个,分别是2016年的川麦82、川育27、科成麦4号,2015年的R802、川14品16、渝1318、川13015、滇麦2号,2014年的13A8,说明高抗穗发芽的品种(系)较少。

表3 2012-2018长江上游小麦区试品种(系)品质混合样品数统计
用KASP标记检测两个穗发芽抗性位点的结果表明,在TaPHS1位点646SNP位置上,96个品种(系)和对照为抗穗发芽基因型;3个品种(系)为感穗发芽基因型;其余10个品种(系)在该位点的基因型不能判定。含有该抗性等位基因的品种(系)的平均混合点率为0.672,而含感穗发芽基因型的川麦605、绵阳1302和德102的混合点率分别为0.625、0.43、0.625。在TaSdr-A1位点,对照和106个品种(系)是感穗发芽基因型,2个品种(系)为抗穗发芽基因型,1个品种为杂合类型,含抗穗发芽基因型的品种(系)绵麦1302和W18的混合点率是0.430和0.75。两个位点均含抗穗发芽基因的品系是W18,两个位点均含感穗发芽基因的品种(系)是川麦605和德102,它们之间样品的混合点率差异不大。综合混合点率和两个位点的基因型分析结果,这两个位点不能很好区分长江上游区试品种(系)的穗发芽抗性。
2.2 试验品种(系)的籽粒容重分级及粒重、籽粒大小和硬度位点的基因型分析
根据容重将109个品种(系)分成六个等级(两年试验的取高等级),一等和二等共有77个,四等以下的仅18个。13次对照样品中,9次达到了二等以上,3次四等以下,对照等级变差可能与2016和2017发生严重冻害和条锈病有关,推测部分品种(系)的籽粒容重也受到类似影响。
用KASP标记检测7个粒重位点、2个籽粒大小位点和3个籽粒硬度位点发现,109个品种(系)中,多数品种(系)在TaGW2-6B、TaGS-D1、Tabas、TaSus2-2A和TaSus1-7B位点分别为Hap-1,2,3、D1a、B1a、Hap-A和Hap-T,是高粒重基因型;在TaGs2-B1和TaSus1-7A位点分别为B1a和Hap-1,3,5,是低粒重基因型;川麦42在TaGW2-6B位点是Hap-4,其余6个位点与多数品种(系)的基因型相同。在TaGS5-A1和TaGASR-A1两个籽粒大小位点,多数品种(系)是A1b大籽粒基因型和H1g小籽粒基因型。在Pina-D1和Pinb-D1位点,多数品种(系)分别是D1a和D1a,为软质基因型;在Pinb2-v2位点有75个是B2b,为相对硬质类型。不同基因型在相同容重等级的品种(系)数间的分布频率差异不明显,说明容重与这些基因的相关性不大(表4)。

表4 2012-2018长江上游小麦品种试验中占多数的基因型在不同容重级别品种(系)中的分布
2.3 试验品种(系)的品质情况及HMW、LMW位点和1BL·1RS基因型分布
由表5可知,对比品种审定标准[9],供试材料各被测指标的平均值基本符合中筋品种标准。各指标表现为:粗蛋白质含量10.06%~ 14.7%,湿面筋含量15.5%~30.7%,面团稳定时间在 0.6~15.5 min之间,吸水率48.4%~ 70.7%。说明这4个指标在品种(系)间的差别较大。参试两年品种(系)各指标的平均值均低于参试一年的品种(系),对照的粗蛋白质含量和湿面筋含量平均最低。说明年份对各指标的测定值有一定影响,多年的数据更能客观反映材料的品质。

表5 2012-2018长江上游小麦品种试验品种(系)品质指标统计
109个品种(系)中,含糯小麦2个,弱筋小麦6个,中筋小麦101个。在101个中筋品种(系)中,4指标全部符合中筋标准的仅20个,其余81个不完全符合中筋标准。在81个不完全符合中筋标准的品种(系)中,59个粗蛋白质含量达中筋及以上标准(表6),47个湿面筋含量达中筋及以上标准,43个吸水率达中筋及以上标准,36个稳定时间达中筋及以上标准。有38个粗蛋白质含量符合中筋及以上标准而稳定时间为弱筋,14个粗蛋白质含量为弱筋而稳定时间为中筋,这两个类型在81个品种(系)中占多数。

表6 2012-2018长江上游国家品种试验品种(系)品质指标分类及部分高低分子量麦谷蛋白亚基的基因型
高、低谷蛋白亚基位点和1BL·1RS易位的KASP标记分析表明:在109个品种(系)中,Glu-A1位点的1/2*基因型、Glu-D1位点的2+12基因型、非Glu-A3g基因型、非Glu-B3g基因型、非Glu-A3d和非1BL·1RS易位是主要基因型(表6)。云麦49的KASP为Glu-B3g基因型,是 1BL·1RS易位,用功能标记验证得到相同的结果。已知Glu-B3g在1BS上,而云麦49为1BL·1RS易位但含有Glu-B3g,值得进一步的研究。
6个弱筋小麦在Glu-A1位点、Glu-A3g和Glu-A3d的基因型是一致的,为2+12、非Glu-A3g和Glu-A3d基因型。有9个完全达到中筋标准的品种在这3个位点的基因型与6个弱筋小麦完全相同。中筋品系川14品16与弱筋品系11P2-4、中筋品系09J76与弱筋品系BL227、中筋品系R801与弱筋品系11P2-4在5个被测位点的基因型完全相同;还有较多品种(系)在5个被测位点基因型相同但品质不同,说明仅这5个位点的基因型不能完全解释小麦品种间品质的差异。
Glu-D1位点的5+10基因型,在109个品种(系)中占39.4%;而在79个粗蛋白质含量达到中筋及以上标准的品种(系)中有34个,占43%;在67个湿面筋含量达到中筋及以上标准的品种(系)中有28个,占41.8%;在56个稳定时间达到中筋及以上标准的品种(系)中有26个,占 46.4%;在63个吸水率符合中筋及以上标准的品种(系)中有28个,占44.4%。综上,5+10亚基在4个指标分别达到中筋标准中的比例均高于在109个品种(系)中的比例,推测在长江上游区小麦品种中,5+10亚基对品质提升有一定的贡献。
2.4 区试对照川麦42和试验二年品种(系)年度间的品质变化
2012-2018川麦42有13个样品,其中仅2018 B组、2017组、2016 A组和2012 B组共4个样品达弱筋品种标准;2013 B组样品达中筋品种标准;其余8个样品不完全符合品质分类指标,划为中筋类型(表7)。由表7可看出,在7年里,川麦42在弱筋和中筋之间徘徊,更多的是偏弱筋小麦;以稳定时间的变化最大。

表7 川麦42和试验二年品种(系)在2012-2018长江上游小麦品种试验中品质表现
15个参试二年的品种(系)中,仅川14品16在两年完全达到中筋标准。各品质指标在年度间均有变化,容重、湿面筋含量和稳定时间年度间变化较大,粗蛋白质含量和吸水量年度间的变化较小。综合两年的品质数据,川麦604、川农32和N08-51在其中一年的指标完全达到中筋品种标准,可望在合适的条件下生产出符合中筋品种标准的中筋小麦,而绵麦312和N08-51在其中一年的指标达到弱筋品种标准,可望在合适的条件下生产出符合弱筋品种标准的弱筋小麦。
3 讨 论
与专门研究区域品种品质变化的试验不同[1,2,3,29],区试的品质数据是跨年度多品种数据,很难得到规律性的结果;但因长江上游区域参加国家级区试的品种是长江上游七省市通过省级审定或完成省级区试2年的品种(系),及时分析区试品质数据,可为育种者提供该区小麦品种品质的基本状况,以发现影响因素,找到改良途径。
3.1 影响长江上游小麦品质的因素及改良策略
3.1.1 穗发芽严重影响长江上游小麦品质
发生穗发芽的小麦,其降落值会大幅降低。张 艳等[27]研究认为,小麦籽粒发芽会严重影响降落值等,对蛋白质质量的影响大于对蛋白质含量的影响,对面粉品质的影响在品种间存在差异。晏本菊等[2]也认为,降落值是影响四川省小麦加工品质的主要因素之一,是受年份天气情况影响最大的品质性状。国家级品种试验的品质分析仅采用降落值高于200 s的样品,把降落值低于200 s的样品视为芽麦,本研究根据测试样品的混合点数推测在长江上游麦区小麦穗发芽普遍存在,且品种间穗发芽抗性有较大差异,具高穗发芽抗性的品种较少。如果将参试品种的全部样品混合用于品质分析,那么符合中筋标准的品种数量会更少。对长江上游麦区的品质育种,提高穗发芽抗性是改良的关键。
本研究检测的TaPHS1是位于3AS上的主效抗穗发芽基因[13],该基因启动子区的序列突变是穗发芽抗性的主要原因[14],但我们试了多次,其分型结果不理想,本研究结果仅为该基因第646碱基G/A SNP的KASP标记结果。TaSdr-A1位于2A染色体上,一个SNP的差异与发芽指数高度相关,具有抗性的TaSdr-A1a在中国地方品种中频率很高[15]。在本研究中,这两个位点不能很好区分长江上游区试品种(系)的穗发芽抗性,对TaPHS1位点基因型还需要进一步检测其启动子区及其他位置的SNP才能得到判断,也需要进一步开发其他控制穗发芽的位点。
3.1.2 长江上游小麦品种蛋白质数量和质量指标的不均衡严重影响该区小麦品质
粗蛋白质含量是蛋白质数量指标,稳定时间是蛋白质质量指标,而湿面筋含量与蛋白质的数量和质量均相关。胡学旭等[28-29]认为,我国小麦从加工品质指标来看,品质类型间互相交错,强筋不强,弱筋不弱;弱筋小麦蛋白质和湿面筋含量逐年上升,使弱筋特性逐渐下降。强筋小麦的沉降值逐年下降,而弱筋小麦呈上升趋势,导致强、弱筋小麦品质下降。从本研究结果看,长江上游麦区小麦品质现状是弱筋品种少,大部分中筋品种的蛋白质数量和质量不均衡,导致适合优质面条、馒头加工的品种较少,而传统的适合酿酒用的小麦品种因蛋白质含量的提高等,也逐渐不能满足酿酒企业的需求。
适宜的栽培措施可影响蛋白质数量。本研究中,对照品种川麦42和15个试验二年品种的品质结果表明,部分品种在适宜的地点和栽培条件下,可望生产完全达标的中筋小麦或弱筋小麦。本研究中的81个不完全达标的中筋品种(系)中,有38个粗蛋白质含量达到中筋及以上,而稳定时间为弱筋的品种(系),为高数量低质量型品种;有14个粗蛋白质含量为弱筋,而稳定时间为中筋的品种,为低数量高质量型(表6)。针对蛋白质数量和质量不均衡的品种,可调整栽培方式,将38个高数量、低质量型品种的粗蛋白质含量降到12%以下,14个低数量、高质量型品种的粗蛋白质含量提高到12%以上,从而提高长江上游麦区中筋小麦品质。而在遗传改良上,将提高穗发芽和提高稳定时间(前38个品种)或降低稳定时间(后14个品种)设为品质改良目标,也能提高该区小麦品质。
3.2 KASP标记检测基因型在育种中的意义
明确育种群体和育成品种中关键性状的基因型,是育种家进行品种改良和分子育种的基础,KASP标记提供了一种高通量检测基因型的方法。Rasheed等[6]开发了70个小麦已克隆基因的KASP标记用于小麦育种材料的基因型检测,涉及光周期等多个性状;中国农科院作科所夏先春老师实验室在继续开发更多已克隆基因的KASP标记(董亚超私人通信)。
目前,有些KASP标记的有效性还需要在育种群体中进一步验证,主要有以下两个原因。首先,一个基因的SNP较多,对基因单倍型的清楚认识是KASP标记应用于育种的基础。KASP标记是根据基因内某个SNP设计的,单个SNP不能完全代表基因型,育种家需要寻找适合自己育种群体的单倍型用于基因型选择。如本研究中检测的TaPHS1位点,在其启动子区-222,编码区646和666位置的SNP对穗发芽抗性都很重要[14];本研究使用的Glu-A1位点的KASP标记是依据该位点第1883碱基处的InDel开发的标记,而没有依据该位点1594碱基处G/A单碱基的差异,故不能区分1和2*亚基[10]。其次,某些位点等位基因的复杂性使分子标记的准确率有待进一步验证。如本研究检测的小麦低分子量麦谷蛋白亚基等位基因Glu-A3g、Glu-B3g和Glu-A3d三个分子标记的结果还需验证。
小麦品质是复杂性状,不仅受遗传控制,也受环境影响。本研究的5个品质指标中,面团稳定时间与遗传关系较为紧密,也受环境影响;而容重和粗蛋白质含量、湿面筋含量和吸水率4个性状受环境影响更大。控制品质性状(如穗发芽抗性)的基因很多,本研究用的20个标记是目前与小麦籽粒性状和品质性状较紧密的KASP标记,还有很多基因的KASP标记有待开发(如已克隆的高低分子量麦谷蛋白亚基的多个等位基因)。本研究仅仅针对部分位点进行了检测,因而不能解释品质的差异。期待加强基础研究,克隆更多基因,更好地认识基因与性状和环境的关系;更期望进一步将已有的功能标记转化为KASP或STARP(semi-thermal asymmetric reverse PCR)[30]标记,开发更多好用的分子标记,使基础研究的成果更好地应用于育种实践。
4 结 论
对2012-2018共7年109个试验品种和对照品种在长江上游麦区国家级品种试验里的品质数据表明,(1)所有品种的测试样的平均混合点率为0.66;(2)多数品种籽粒的商品分级在四级以上;(3)按照品种审定标准,在109个品种中,101个是中筋品种,其中有81个品种没完全达到中筋分类指标;(4)穗发芽和蛋白质数量和质量指标不均衡是长江上游麦区的突出问题,品质改良应以增强穗发芽抗性和改良稳定时间为主要目标。
对109个品种20个位点的KASP标记检测,发现以下基因型是占多数的基因型:TaPHS1位点的RioBlanco type基因型,TaSdr-A1位点的A1b基因型,TaGW2-6B位点的Hap-1,2,3基因型,TaGS-D1位点的D1a基因型,TaGs2-B1位点的B1a基因型,Tabas位点的B1a基因型,TaSus2-2A位点的Hap-A基因型,TaSus1-7A位点的Hap-1,3,5基因型,TaSus1-7B位点的Hap-T基因型,TaGS5-A1位点的A1b基因型,TaGASR-A1位点的H1g基因型,Pina-D1位点的D1a基因型,Pinb-D1位点的D1a基因型,Pinb2-v2位点的B2a基因型,Glu-A1位点的1/2*基因型,Glu-D1位点2+12基因型,非Glu-A3g基因型,非Glu-B3g基因型,非Glu-A3d基因型,非1BL·1RS易位。
