小麦 TaGS-D1与 TaFlo2-A1等位变异对粒重的影响
2020-07-30严勇亮付婧璇耿洪伟
张 芳 ,任 毅,严勇亮,付婧璇,耿洪伟
(1.新疆农业大学农学院,农业生物技术重点实验室,新疆乌鲁木齐 830052;2.新疆农业科学院,农作物品种资源研究所,新疆乌鲁木齐 830091)
千粒重是小麦产量构成三要素之一,提高千粒重是小麦增产的有效途径和重要育种目标[1]。小麦千粒重是受微效多基因控制的数量性状,利用可靠的分子标记可提高对小麦粒重选择的准确性和有效性,有利于对高千粒重优异基因型在小麦育种中的充分应用[2]。产量三要素中千粒重是最稳定,具有59%~80%的广义遗传率[3]。胡延吉等[4]认为,粒重的增加是提高小麦产量的主导因素。冯素伟等[5]发现,在穗粒数的贡献相对稳定时,提高千粒重则成为小麦增产的关键因素。由此可见,粒重的遗传改良是对小麦高产非常重要[6]。国内外的研究表明,过去的几十年中,小麦穗数和穗粒数的改良相对缓慢,且对产量提升贡献较小,而千粒重的提高对小麦产量的提升贡献较大[7-11]。因此,在小麦品种选育过程中,注重千粒重的选择是提高小麦产量的重要途径,也成为小麦分子标记辅助育种的一个重要内容[12]。
近年来,小麦粒重相关基因定位取得了很大进展[13-14]。Cui等[15]在5B、6A和7B染色体上发现3个与粒重相关的主效QTL,分别解释表型变异的11.28%~16.06%、5.64%~18.69%和 6.76%~21.16%。Jia等[16]利用来自RIL群体南大2419/望水白的230个家系,将粒重主效QTL定位于3A、4B、4D和5A染色体上。Huang等[17]在2D、4B和5A染色体上发现了3个粒重主效QTL。与此同时,作为育种应用的理想标记,功能标记被用于小麦粒重基因鉴定和克隆[18-19]。Zhang等[20]克隆了普通小麦7DS染色体上的籽粒大小基因TaGS-D1的全长编码序列,并基于40 bp 的插入/缺失(Indel),开发了与千粒重显著相关(P<0.01)的共显性功能标记GS7D。该位点的两个等位基因TaGS-D1a和TaGS-D1b分别与高千粒重和低千粒重相关,对应的扩增片段长度分别为562和522 bp。利用GS7D标记对51份新疆冬小麦进行检测发现,该标记能较好地区分TaGS-D1a和TaGS-D1b等位基因[21]。Flo2是广泛存在植物中的基因家族,小麦TaFlo2基因与粒重紧密相关,其表达量对小麦籽粒的淀粉含量和蛋白质合成有影响,从而影响小麦的粒重。Sajjad等[22]克隆了普通小麦2A染色体上的粒重基因TaFlo2-A1的全长编码序列,并根据其等位基因TaFlo2-A1a和TaFlo2-A1b在启动子区存在8 bp的插入/缺失(Indel),开发了一对共显性功能标记TaFlo2-Indel8。TaFlo2-Indel8标记可在具有等位基因TaFlo2-A1a(与低千粒重相关)和TaFlo2-A1b(与高千粒重相关)的材料中分别扩增出153和 145 bp的片段。虽然目前有许多小麦粒重相关基因的分子标记被开发和利用[23-26],但TaGS-D1和TaFlo2-A1为粒重基因研究的主要对象。这些研究多是对单个基因效应进行分析,而对不同粒重基因组合对千粒重影响尚不清楚。基于此,本研究利用粒重功能标记GS7D和TaFlo2-Indel8对298份国内外冬小麦品种(系)进行检测,对各等位基因、基因组合与千粒重的相关性及其在不同小麦类型中分布规律进行分析,以期探究不同粒重基因型对千粒重的作用,为小麦粒重分子聚合育种提供参考依据。
1 材料与方法
1.1 供试材料
供试的298份冬小麦品种(系)来自12个国家,包括65份国外品种(系)及233份国内四个主要冬麦区的历史品种(系)和当前主栽品种(表2)。其中,国外冬小麦品种(系)分别来自法国(21份)、意大利(9份)、美国(7份)、阿根廷(7份)、罗马尼亚(6份)、俄罗斯(5份)、日本(4份)、匈牙利(2份)、澳大利亚(1份)、德国(1份)、英国(1份)、土耳其(1份);国内冬小麦品种(系)分别来自黄淮冬麦区(145份)、北部冬麦区(55份)、西南冬麦区(22份)和长江中下游区冬麦区(11份)。所有小麦品种(系)均于2016-2017年和2017-2018年度在新疆农科院玛纳斯试验站种植。随机区组设计,2个重复,行长2 m,行距25 cm,3行区,每行播量为100粒,田间管理同大田生产,正常成熟后及时收获,人工脱粒。
1.2 千粒重测定
采用杭州万深检测科技有限公司生产的SC-G型千粒重仪测定小麦千粒重,3次重复,计算平均值。
1.3 分子标记检测
1.3.1 基因组DNA提取
为避免单粒种子偶然性错误,每个品种(系)选取3 粒粒型均匀的种子,粉碎后放入2.0 mL离心管中,参照Lagudah等[27]方法提取基因组DNA,并用1%琼脂糖凝胶电泳技术检测DNA。
1.3.2 PCR扩增与检测
利用Zhang等[20]开发的共显性标记GS7D检测小麦7DS染色体上粒重基因TaGS-D1等位基因;利用Sajjad等[22]开发的共显性标记TaFlo2-Indel8检测小麦2AL染色体上粒重基因TaFlo2-A1等位基因。引物(表1)均由上海生物工程技术服务有限公司合成。
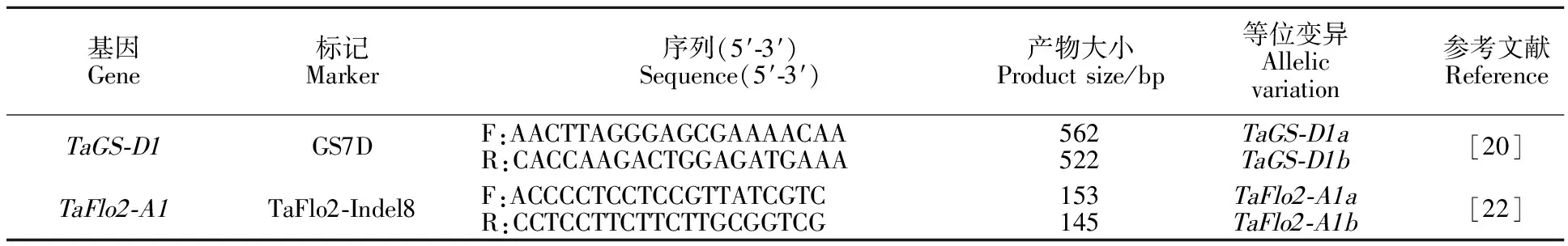
表1 目标基因分子标记信息
PCR体系15 μL,含1 μL的DNA为模板,2×Es Taq MasterMix(Dye)7.5 μL(康为世纪),上、下游引物(10 μmol·L-1)各0.5 μL,用ddH2O补充至15 μL。PCR反应程序为94 ℃ 5 min;94 ℃ 1 min,55~59 ℃ 45 s,72 ℃ 2 min,共35个循环;72 ℃ 10 min。其中,标记GS7D的PCR产物在4.0%琼脂糖凝胶(加入核酸染料)上进行电泳分离,在VILBER LOURMAT凝胶成像系统紫外灯扫描成像并保存到计算机;标记TaFlo2-Indel8的扩增产物用6.0%的变形聚丙酰胺凝胶电泳分离,银染后记录结果。
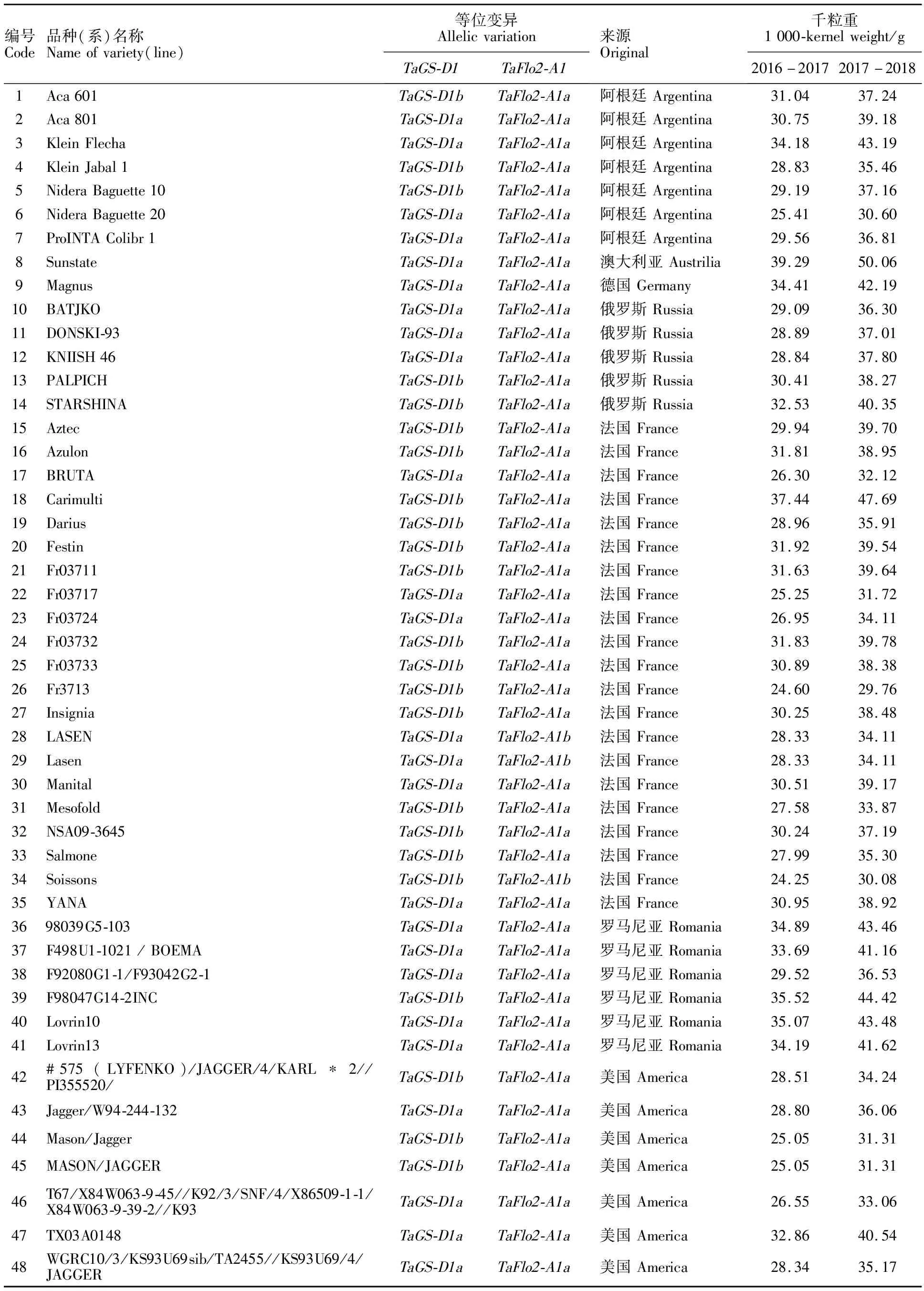
表2 中国和国外小麦品种(系)的千粒重及基因型
1.4 统计分析
采用Excel 2010对测定的千粒重数据进行整理分析,并利用 SPSS 21软件对同一基因不同等位基因品种的千粒重进行t测验和不同基因型品种的千粒重间进行邓肯法多重比较。
2 结果与分析
2.1 小麦千粒重的表型变异
298份小麦品种(系)的两年千粒重平均值为37.65 g,变化范围为24.72~46.82 g,变异系数为12.31%;国外材料的平均千粒重低于国内各冬麦区的材料(表3)。
2.2 TaGS-D1和 TaFlo2-A1位点不同等位基因及其组合频率分布
在298份冬小麦品种(系)中,利用GS7D标记检测出TaGS-D1a和TaGS-D1b2种等位基因,扩增长度分别是562和522 bp(图1),其中具有高千粒重TaGS-D1a等位基因的材料有236份(占79.2%),具有低千粒重TaGS-D1b等位基因的材料有62份(占20.8%)。TaGS-D1a和TaGS-D1b在国外及国内各冬麦区中分布不同(表4)。在所有材料中,整体上TaGS-D1a的分布频率大于TaGS-D1b。从材料来源看,TaGS-D1a的分布频率表现为西南冬麦区(95.5%)>长江中下游冬麦区(90.9%)>北部冬麦区(85.5%)>黄淮冬麦区(78.6%)>国外(55.4%)。
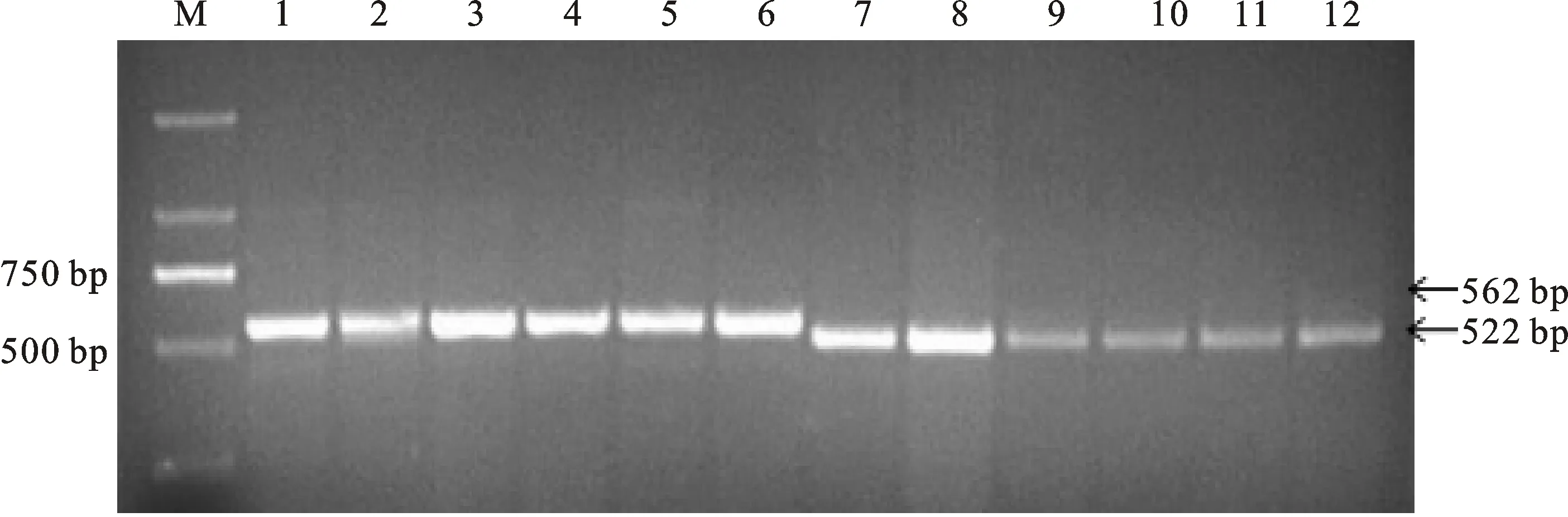
M:DL2000;1:小偃6号;2:豫麦47;3:周麦25;4:鲁麦7号;5:济麦21;6:阿夫;7:碧蚂1号;8:百农3217;9:鲁麦9号;10:周麦31;11:泰农731;12:豫麦21.
在供试材料中,利用TaFlo2-Indel8标记共扩增出145和153 bp两种片段类型(图2),说明存在TaFlo2-A1b和TaFlo2-A1a2种等位基因。在298份冬小麦品种(系)中,56份具有高千粒重TaFlo2-A1b等位基因,242份具有低千粒重TaFlo2-A1a等位基因,说明具有高千粒重TaFlo2-A1b等位基因的材料分布频率(18.8%)低于具有低千粒重TaFlo2-A1a等位基因的材料(81.2%)。从材料来源看,在参试材料中TaFlo2-A1b的分布频率表现为长江中下游冬麦区(36.4%)>北部冬麦区(27.3%)>黄淮冬麦区(18.6%)>西南冬麦区(13.6%)>国外(10.8%)(表4)。
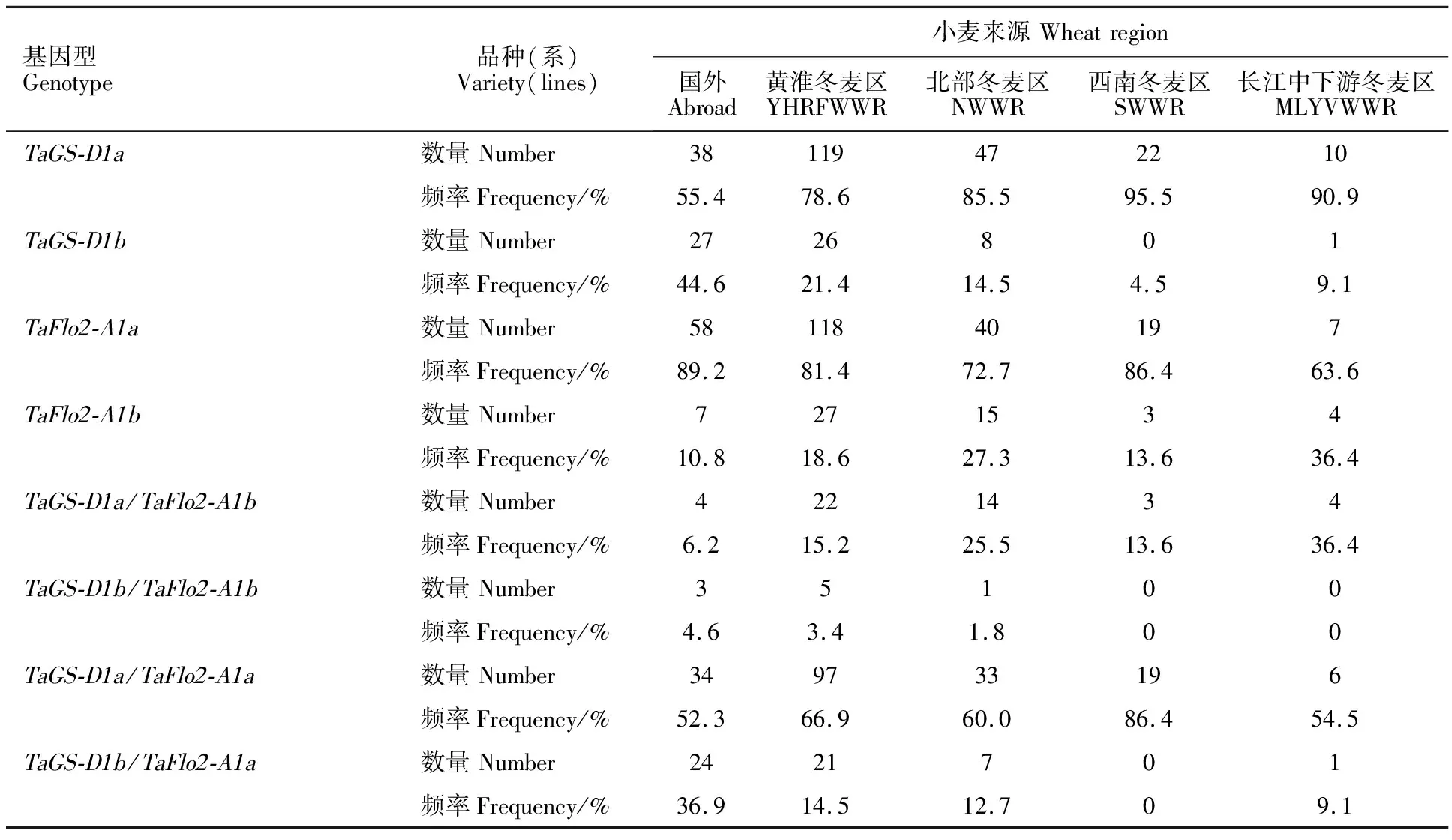
表4 不同类型冬小麦品种(系)等位变异的分布频率
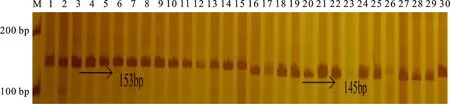
M:DL2000;1:淮麦18;2:淮麦20;3:淮麦21;4:皖麦29;5:皖麦33;6:皖麦38;7:矮抗58;8:武农 148;9:新麦19;10:新麦9号;11:豫麦13;12:豫麦18;13:豫麦34;14:豫麦35;15:豫麦57;16:兰考906;17:豫麦47;18:豫麦49;19:郑9023;20:中麦892;21:中麦871;22:周麦12;23:周麦16;24:周麦18;25:周麦19;26:周麦23;27:周麦30;28:扬麦15;29:鲁麦23;30:烟农18.
在298份参试材料中共检测出4种基因组合(基因型),分别为TaGS-D1a/TaFlo2-A1b、TaGS-D1a/TaFlo2-A1a、TaGS-D1b/TaFlo2-A1b和TaGS-D1b/TaFlo2-A1a,其分布频率分别为15.4%、63.4%、3.0%和18.1%(表5)。在国内各冬麦区和国外品种(系)中,TaGS-D1a/TaFlo2-A1a基因型均占比最大,TaGS-D1b/TaFlo2-A1b基因型均占比最小或没有。高千粒重基因型在不同小麦类型中均占比较小,且国外品种(系)均低于国内各冬麦区材料。不同来源小麦材料中4种基因型的分布频率不同。其中,TaGS-D1a/TaFlo2-A1b基因型在长江中下游冬麦区分布频率较高,达到36.4%;TaGS-D1a/TaFlo2-A1a基因型在西南冬麦区分布频率较高,占比86.4%;TaGS-D1b/TaFlo2-A1b基因型在国外品种(系)中的分布频率较高,为4.6%;TaGS-D1b/TaFlo2-A1a基因型在国外品种(系)中的分布频率较高,达到36.9%。这说明国内小麦资源中高千粒重的TaGS-D1a/TaFlo2-A1b基因型丰富,尤其是在长江中下游冬麦区分布较多,而低千粒重的TaGS-D1b/TaFlo2-A1a基因型在国外品种(系)中分布较多。
2.3 TaGS-D1和 TaFlo2-A1位点不同等位基因及其组合千粒重比较
对TaGS-D1和TaFlo2-A1等位基因及其组合的平均千粒重进行分析(表5)表明,TaGS-D1a和TaFlo2-A1b等位基因的平均千粒重分别为38.18和39.12 g,均极显著高于对应的等位基因TaGS-D1b和TaFlo2-A1a,进一步印证TaGS-D1a和TaFlo2-A1b等位基因与高千粒重相关,属于优异等位变异。在TaGS-D1和TaFlo2-A1位点形成的4种基因型中,TaGS-D1a/TaFlo2-A1b的平均千粒重为39.82 g,极显著高于其他基因型;TaGS-D1b/TaFlo2-A1a的平均千粒重为35.59 g,低于其他3种基因型,说明TaGS-D1a/TaFlo2-A1b与高千粒重相关且为最优组合,对千粒重的增加具有较大作用,且高于单一高千粒重等位基因增效作用。

表5 不同基因间和不同基因型间千粒重分析
3 讨 论
小麦粒重相关分子标记辅助选择对小麦育种效率至关重要[28]。尽管粒重基因在品种间和环境间均存在显著差异,但主要受遗传控制,通过分子标记技术,可有效提高小麦高产育种效率。利用TaGS-D1位点功能标记GS7D和TaFlo2-A1位点功能标记TaFlo2-InDel8可较好地区分出小麦千粒重的大小。Zhang等[20]利用TaGS-D1位点的显性标记GS7D检测 175 份中国小麦材料,结果表明,TaGS-D1a等位基因分布频率较高(80%),TaGS-D1a等位基因是优异及优势等位变异。简大为等也[21]印证了在51份新疆冬小麦资源中TaGS-D1a等位基因(70.6%)分布频率明显高于TaGS-D1b等位基因(29.4%)。Sajjad等[22]利用TaFlo2-A1位点的显性标记TaFlo2-InDel8检测262份中国小麦微核心种质,结果显示TaFlo2-A1b占比仅为16.5%。本研究利用两对功能标记GS7D和TaFlo2-InDel8对298份冬小麦品种(系)进行检测,结果与Zhang等[20]和Sajjad等[22]研究基本一致。其中高千粒重TaGS-D1a分布频率较高,可能与育种家在品种选育过程中优先选择该等位变异有关。而高千粒重TaFlo2-A1b分布频率较低,可能与千粒重受多基因控制有关,其效应被其他粒重基因所掩盖,仅在少数材料中体现出高千粒重TaFlo2-A1b等位基因。虽然TaFlo2-A1b等位基因分布频率远低于TaFlo2-A1a等位基因,但TaFlo2-A1b等位基因材料较TaFlo2-A1a等位基因材料差异极显著(P<0.01)。
不同来源小麦品种(系)中,具有高千粒重的TaGS-D1a和TaFlo2-A1b等位基因及其TaGS-D1a/TaFlo2-A1b基因型的分布频率均存在较大差异(表4),其中长江中下游冬麦区高千粒重分布频率较高,而国外品种(系)分布较低。究其原因一方面可能与多年粒重人工选择有关。长江中下游冬麦区以大穗大粒选择为主,对粒重的重视程度和选择力度较强,而国外品种产量的变化体现在穗粒数和穗数,千粒重变化不大,对高千粒重选择较小,该等位基因未被广泛选择利用[29]。另一方面可能与育种需求及不同地理环境的生态条件有关。高千粒重小麦将更有利于我国小麦粒重遗传改良及筛选高产品种。我国长江中下游冬麦区气候温和,雨水充沛,但易发生病虫害,导致穗数不足,千粒重优势突出[30],而国外人口密度基数较小,对于高千粒重选育需求相对较低,因此高千粒重TaGS-D1a和TaFlo2-A1b等位基因育种选择利用将集中在个别少数国家中。此外,还可能在品种改良过程中,我们仅关注国外材料中的一些个别性,从而造成国外品种(系)中高千粒重TaGS-D1a和TaFlo2-A1b等位基因占比较少,同时反映出不同地域品种在遗传组成上的明显差异[31-33]。
从小麦主栽品种推广应用以及目前一些新品种的高产纪录来看,高千粒重对我国小麦产量的提高至关重要[34,2]。高千粒重TaGS-D1a/TaFlo2-A1b基因型的千粒重显著高于其他组合,且高于单基因高千粒重TaGS-D1a与TaFlo2-A1b等位基因,说明高千粒重基因型将对小麦粒重遗传改良具有较大的提升空间,同时可作为小麦粒重的辅助选择的实用性标记。时佳等[35]也证明了较单一高千粒重等位基因,多个基因型可获得更高千粒重。品种审定中可利用高千粒重基因型将有效准确地获得高千粒重品种材料;育种家在高千粒重选育过程中也可以通过TaGS-D1a/TaFlo2-A1b基因型,对其选配亲本提供研究基础[36],筛选出具有高产潜力的小麦种植资源,从而提升我国小麦千粒重。千粒重除受多基因控制外,还受籽粒大小、灌浆速率等因素影响。在今后的高千粒重小麦品种选育过程中,对基因型的累加聚合和多指标综合分析十分重要,将是一种高产育种的研究方向,可加速高产小麦育种步伐。
