小麦-卵穗山羊草1Mg异附加系蛋白质品质形成解析
2020-07-30丁梦云李慧敏李少鹏仝靖洋王中华
羊 阳,丁梦云,郭 雷,李慧敏,李少鹏,仝靖洋,王中华,高 欣
(西北农林科技大学农学院,陕西杨凌 712100)
普通小麦(TriticumaestivumL.)作为世界上分布最广泛、最重要的粮食作物之一,因其独特的面筋结构而具有较强的可加工性,被广泛应用于食品生产。小麦麦谷蛋白是面筋的骨架成分,根据其分子量的大小被分为高分子量麦谷蛋白亚基(HMW-GS)和低分子量麦谷蛋白亚基(LMW-GS),其中HMW-GS仅占普通小麦总麦谷蛋白的7%~15%,但解释了面筋、面团和小麦加工品质特性变异的45%~70%[1],是影响小麦面制品品质的关键因素。HMW-GS的组成和含量对小麦面团的加工品质有显著影响[2]。在亚基组成方面,一般认为5+10、17+18等为优质亚基组合,而2+10、5+12、2+12为劣质亚基组合;在含量方面,HMW-GS含量较高的小麦面团表现出更好的加工品质和烘培品质。我国当前小麦育种工作中,小麦主产区都存在着由于优质亲本源单一导致新品种遗传相似性高、育成突破性品种难度加大的问题,尤其在品质育种方面[3]。挖掘新型优质的HMW-GS资源,对小麦品质改良具有重要意义,也是当前小麦育种研究的热点。
小麦的近缘属种中存在着许多对小麦品种改良有着重要作用的基因,利用远缘杂交和分子生物学技术创制小麦近缘属种与小麦异附加系已经成为小麦品种改良的重要方法[4-5]。有研究利用小麦-黑麦异附加系发现并定位了来自于黑麦染色体的抗旱基因[6]。Olson等[7]利用远缘杂交获得了抗条锈病的小麦-粗山羊草异附加系。目前,利用小麦近缘属种的优质基因已经创建了包括山羊草、水草、类麦、偃麦草、赖草、鹅观草、大麦、黑麦、冰草等34种属小麦近缘植物200多个异附加系,极大丰富了小麦育种种质资源[8]。
卵穗山羊草(Aegilopsgeniculata,2n=4x=28,UgUgMgMg)是普通小麦的一个近缘种属,其在抗虫、抗病、抗逆等方面均表现出优异的特性。已有研究证明,卵穗山羊草对菲利普孢囊线虫存在抗性,并具有一定的抗旱性;其所携带的抗白粉病基因具有可遗传性并在后代中能良好表达[9-11]。
小麦近缘属种中存在优质HMW-GS的编码基因,对改善小麦面团的品质有显著作用[12-14]。但利用卵穗山羊草进行品质改良的研究较少。本研究拟以中国春(CS)为对照,解析CS-卵穗山羊草1Mg异附加系(CS-1Mg)面团的品质特性,为丰富小麦种质资源、进一步促进小麦品质改良提供科学依据。
1 材料与方法
1.1 试验材料
试验材料为中国春(CS)以及以中国春为背景的小麦-卵穗山羊草1Mg异附加系(CS-1Mg,染色体组成2n=44)。材料于2017-2018年种植于中国陕西省杨陵区西北农林科技大学作物教学标本区(108°4′E,34°16′N)。每个小区种植6行,行长为2 m,行距为0.25 m,株距为0.05 m。设置三个生物学重复。
1.2 试验方法
1.2.1 麦谷蛋白的分离
参考Gao等[15]的方法,利用4%的上层浓缩胶和10%的下层分离胶建立连续的SDS-PAGE分离体系。参考Li等[16]方法,使用高效液相色谱仪(1260,Agilent,USA)对供试材料的HMW-GS进行RP-HPLC精细分离,并利用Agilent对液相图谱的峰面积进行手动积分,计算各HMW-GS的表达量。
1.2.2 籽粒发育过程中谷蛋白聚合体积累的动态检测
利用排阻-高效液相色谱测定开花后籽粒发育不同时期(花后4 d、7 d、13 d、16 d、21 d、26 d、31 d和成熟)SDS不溶性谷蛋白聚合体(UPP)和SDS可溶性谷蛋白聚合体(EPP)的含量。UPP与EPP的提取参考Zhao等[17]的方法。UPP占比(%)=UPP的峰面积/(UPP+EPP)峰面积×100%,以不同时期的UPP占比表示籽粒发育过程中谷蛋白聚合体的动态变化。
1.2.3 面筋样品的制备
供试材料的面筋样品参考Gao等[18]的方法通过手洗法得到。得到的面筋样品被分为两份,一份直接用于微观结构的观察,另一份在超低温冷冻干燥机(GWSP,Jiwei,Shanghai,China)下冻干48 h后研磨成粉,用于二级结构的测定。
1.2.4 面筋样品二级结构的测定
利用傅里叶红外变换光谱仪(Vetex70,Bruker Optics,Germany)测定供试材料面筋蛋白的二级结构。测定其在1 600~1 700 nm波段的吸光度,测得的光谱图像用Peakfit(PeakFit v4.12,SeaSolve Software Inc.USA)多峰拟合处理后得到二级结构分布。
1.2.5 面筋样品微观结构的观察及定量分析
参考Li等[19]的方法,利用场发射扫描电子显微镜(Scientz-750F,SCIENTZ,Ningbo,China)对面筋样品进行形态学观察。扫描电镜的电压为15 kV,放大倍数为2 000倍。
激光共聚焦显微镜(CLSM)的观察方法参考Bernklau等[20]的方法,并稍作修改。利用Olympus IX83 反向电子显微镜对样品进行观测,其自身带有FLUOVIEW(FV1200)生物激光共聚焦扫描系统并配有LD559半导体激光发生器和物镜。分辨率为800×800像素,大小为317 μm×317 μm。
根据Gao等[15]的方法利用激光共聚焦显微镜(Olympus,Tokyo,Japan)结合AngioTool软件(National Cancer Institute,National Institute of Health,Maryland,USA)对面筋的微观网络结构进行分析,通过蛋白质面积、蛋白质面积比率、蛋白节点数、节点密度、蛋白长度、蛋白端点数、孔隙度、端点率和分支率共9个参数定量分析两份供试材料的面筋蛋白结构。
1.2.6 部分籽粒品质指标测定
利用近红外谷物分析仪(Diode Array 7250,Perten,Sweden)测定供试材料籽粒的含水量、蛋白质含量、湿面筋含量、淀粉含量、沉降值。
1.2.7 面团混揉特性的测定
参考Rosell等[21]的方法,利用肖邦混揉仪(Chopin,Tripette and Renaud,France)测定供试样品的吸水率、面团形成时间与稳定时间。
1.3 数据处理
利用Excel 2016进行数据处理,利用SPSS 20.0进行方差分析和多重比较。
2 结果与分析
2.1 谷蛋白组成分析
2.1.1 谷蛋白亚基的分离和鉴定
从CS与CS-1Mg麦谷蛋白的分离结果(图1A)看,两者的LMW-GS组成未出现明显差异。在HMW-GS组成方面,与CS相比,CS-1Mg的HMW-GS组成有四个亚基,表现为Dx2亚基缺失,但出现一个未知新亚基(UK),其余三个HMW-GS亚基与CS一致,推测由于其携带了卵穗山羊草1Mg染色体。小麦的HMW-GS虽然只占小麦籽粒蛋白质含量的7%~15%,但是它对小麦面团的强度和弹性有着重要作用,新型未知亚基的出现可能影响CS-1Mg的品质[22]。利用RP-HPLC对CS与CS-1Mg的HMW-GS进行了精细分离(图1B),可以看出,CS-1Mg与CS样品均含有四个峰,表明两个样品分别含有四个亚基,其中三个亚基的出峰时间相同,推测其为Bx7、By8和Dy12亚基。CS-1Mg含有一个不同于Dx2亚基的一个新亚基,这与SDS-PAGE的结果相吻合。

图1 供试材料麦谷蛋白组成的SDS-PAGE和RP-HPLC鉴定
2.1.2 HMW-GS定量分析
HMW-GS组分分布结果(表1)表明,与CS比较,CS-1Mg中Bx7亚基含量显著提高,而By8亚基含量略微下降,Dy12亚基含量无明显变化。CS-1Mg中新型未知亚基含量明显高于原CS中的Dx2亚基,推测会导致这两份材料的面团特性与功能存在差异。

表1 供试材料HMW-GS的组分分布
2.2 籽粒发育过程中谷蛋白聚合体动态积累过程
由图3可知,CS-1Mg的UPP占比在整个籽粒发育过程中均高于CS,由于UPP占比与面团的强度有明显的正相关关系[23],推测CS-1Mg的面筋品质优于CS。在籽粒发育的初期(4~7 d)UPP迅速积累,CS与CS-1Mg变化趋势基本相同。在籽粒发育的中前期(7~10 d),CS的UPP占比出现明显下降的趋势,而CS-1Mg则持续增加。到籽粒发育的中期时(10~16 d),CS与CS-1Mg的UPP占比均呈增加趋势,但在CS-1Mg中的增加速率明显高于CS。在籽粒发育中后期(16~21 d),CS与CS-1Mg均出现一段UPP占比的下降期,但CS-1Mg的下降程度较小。接近成熟期时,两个材料均有一段UPP快速积累期,但CS-1Mg的快速积累期较短。
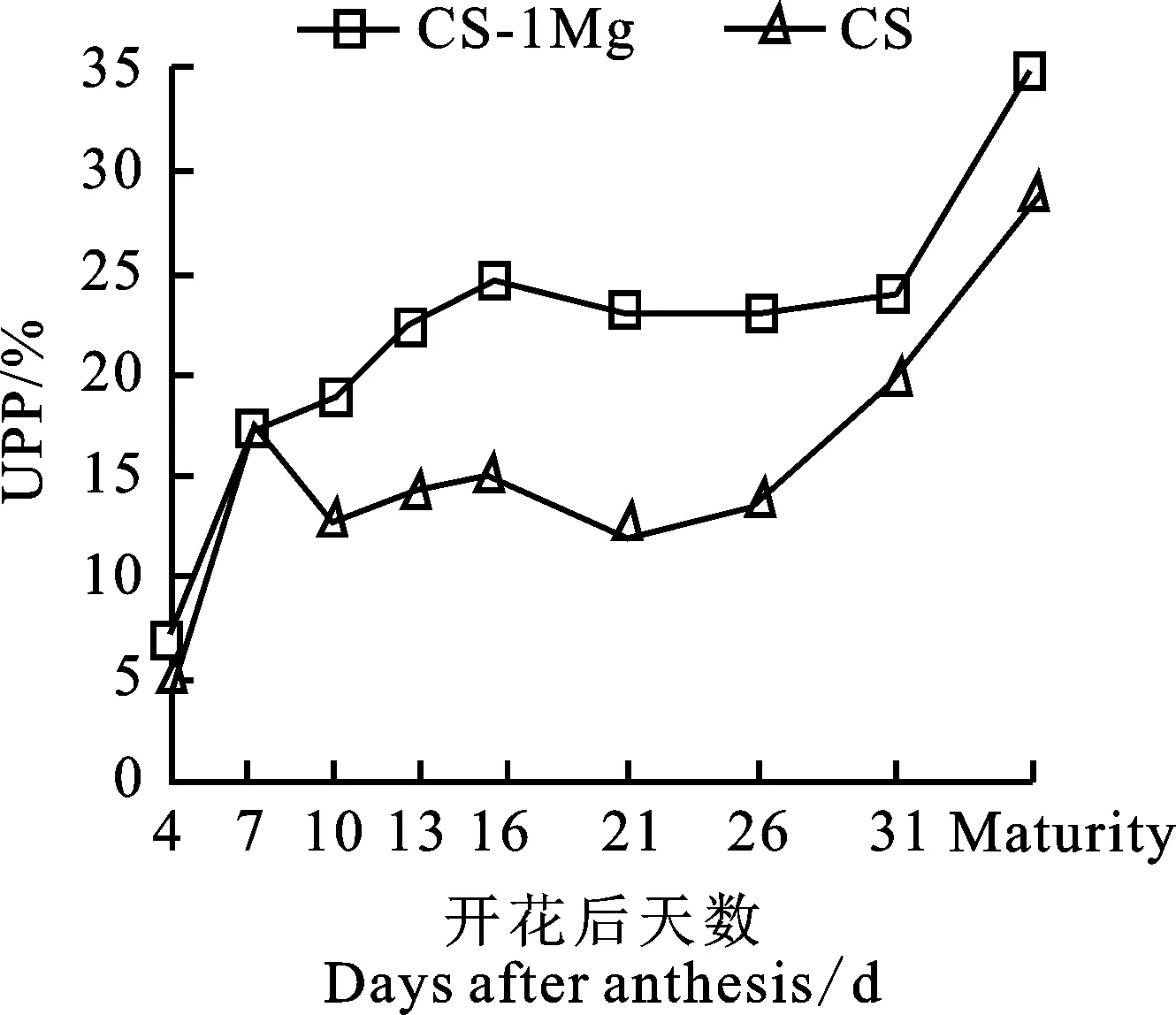
图2 小麦花后不同时期籽粒的UPP%变化
2.3 面筋蛋白二级结构的分析
与CS相比,CS-1Mg的面筋蛋白中 α螺旋含量较低,水合β折叠含量较高,其β转角含量显著提高(P<0.05),分子间β折叠含量显著下降(P<0.05)(表2)。面团的流变学特性与水合β折叠、β转角之间存在着显著的正相关关系,而与α螺旋、分子间β折叠存在负相关关系[24]。β折叠和β转角相较于α螺旋有着更好的亲水性,能够更好的结合面团中的水分,形成分子间氢键,增加面筋蛋白的稳定性和强度,进一步提高其流变学特性[25]。因此,推测CS-1Mg中因为含量更高的水合β折叠、β转角使其与CS相比具有更好的水结合力,同时含量较低的分子间β折叠使其能更好的破坏蛋白间相互作用,进而促进面筋蛋白网状的形成[26]。
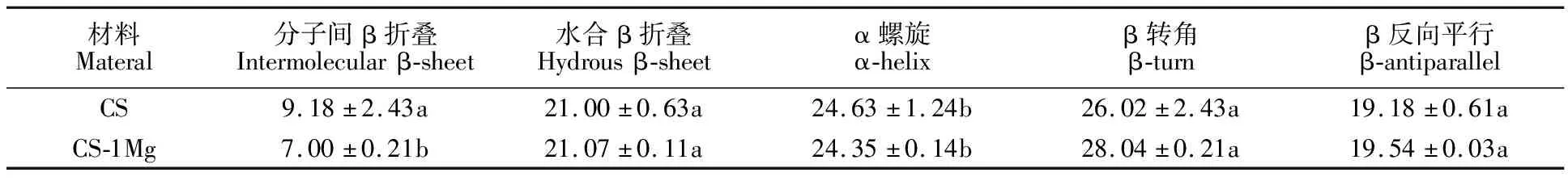
表2 两份材料的面筋蛋白二级结构
2.4 面筋蛋白的微观结构分析
2.4.1 扫描电镜结果分析
相较于CS,CS-1Mg的面筋蛋白网络分布更均匀且孔径较小(图3),说明其面筋蛋白具有更紧密的网络结构,面筋蛋白分子之间连接的更紧凑,使CS-1Mg在接受外界压力后能更好的保持其原有的结构,宏观上表现为流变学特性的提高。

A:CS面筋在2 000×的扫描图像;B:CS-1Mg面筋在在2 000×的扫描图像;标尺代表10 μm。
2.4.2 激光共聚焦图像定量分析
与扫描电镜结果相似的是,激光共聚焦图像(图4)中,CS-1Mg同样表现出相较于CS更均匀紧密的网络结构。结合AngioTool定量分析结果(表3)发现,CS-1Mg与CS相比,蛋白质面积略微增加且蛋白节点数和节点密度显著增加(P<0.05),表明CS-1Mg蛋白含量增加;CS-1Mg的蛋白端点数和端点率显著提高(P<0.05),说明其面筋连接更紧密[21];CS-1Mg的孔隙度较于CS明显下降(P<0.05),这表明了CS-1Mg面筋网络分布更均匀。
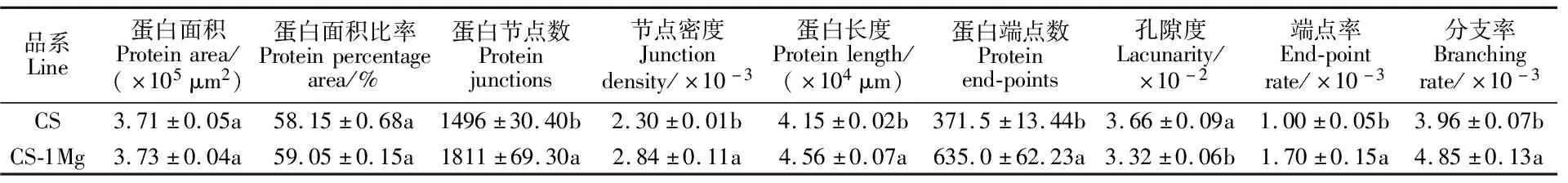
表3 两份小麦品系面筋蛋白微观结构定量分析
CS-1Mg在面筋网络结构的形态学特点与数量指标上均优于CS对照,可归因于其更优质的亚基组成和更好的二级结构分布,从而形成了更紧密的面筋网络结构,推测这些变化会直接影响面团的混揉特性。
2.5 面团混揉特性的分析
利用近红外谷物分析仪分析CS与CS-1Mg籽粒品质特性,结果表明,与CS对照相比,CS-1Mg的淀粉含量变化不显著,蛋白质、湿面筋含量和沉降值显著提高(P<0.05),说明CS-1Mg的籽粒品质优于CS,结合上述结果,可以认为其籽粒品质的提升是由HMW-GS组成变异导致的。表4表明,与CS对照相比,CS-1Mg面团的形成时间与稳定时间均显著提高(P<0.05),形成时间和稳定时间与面筋强度存在着显著正相关关系[27],推测CS-1Mg的面团筋力更高,具有更好的加工品质。
3 讨 论
小麦的HMW-GS只占到小麦籽粒蛋白质含量的7%~15%,但对小麦面团的强度和弹性有着重要作用。育种工作者对有关小麦近源种属植物中的HMW-GS已有一定研究,如Zahra等[12]发现,伊朗山羊草中存在丰富的HMW-GS变异;颜泽洪等[13]在粗山羊草中发现了两种新型HMW-GS。但大部分研究往往只停留在对新型亚基的筛选上,没有对新型亚基对于品质可能出现的提升进行系统的分析和研究。本研究发现,在亚基组成方面,CS-1Mg中出现的新型HMW-GS替换了CS中的Dx2型亚基,打破了2+12的劣质亚基组成,使得其亚基组成优于CS。这可能是由于材料创制过程中卵穗山羊草1Mg染色体上基因的表达或发生了DNA甲基化等表观遗传学修饰使得Dx2基因沉默导致的[28]。在小麦籽粒的发育过程中,HMW-GS和LMW-GS会通过二硫键结合形成谷蛋白聚合体。新型HMW-GS促进了CS-1Mg籽粒发育过程中UPP的积累速率,CS只在籽粒发育的开始和接近成熟期快速的积累UPP,而CS-1Mg在籽粒的发育的全过程中几乎一直保持着一个高于CS的积累速率,呈现出一种稳步上升的趋势。在面筋蛋白二级结构与微观结构上,新型HMW-GS使得一部分由于蛋白聚合作用形成的β折叠转化为亲水的β转角结构,表现出更好的二级结构组成,加强了面团的与水的结合能力,使得CS-1Mg面筋网络结构更紧密,从而提高了面筋强度。
总的来说,新型HMW-GS使得CS-1Mg表现出了更好的蛋白质组分,进一步影响了其二级结构与微观结构,在宏观上表现出流变学特性的提高以及具有更好的加工品质。
在小麦近缘属种中已经发现了许多优秀的抗病、抗虫基因和异源贮藏蛋白基因,且普通小麦具有庞大且复杂的基因组,具备接受与融合外源染色体片段的能力,通过染色体工程转移一条或者多条染色体至小麦中,将近缘属中的优质基因导入到小麦中可创制出小麦异附加系,对扩大小麦育种的种质资源有重大意义。由此已选育出优异的小麦品种(小偃系列、陕麦系列)。本试验所使用的小麦-卵穗山羊草1Mg异附加系克服了卵穗山羊草加工特性差的缺点,能够直接作为试验材料对其籽粒贮藏蛋白与面粉和面团品质进行一系列研究。本研究中未对CS-1Mg的农艺性状、抗逆性、环境适应性等进行测定和评价,无法像Du等[29]筛选出的GN05异附加系一样作为一个潜在的优质品种直接应用于生产实践中。但CS-1Ug中卵穗山羊草1Mg染色体上的异源贮藏蛋白基因能正常表达,所携带的新型HMW-GS能够显著的提高面团的强度,改善面团品质,可以作为中间材料和定向提供优质贮藏蛋白基因源,用于选育优良加工品质的小麦品种。
