青藤碱抑制CaN/NFAT通路信号抑制CIA骨破坏的研究①
2020-07-13张浩然吉金雨侯晓强梅志刚冯知涛
张浩然 吉金雨 侯晓强 颜 岚 梅志刚 冯知涛
(三峡大学医学院国家中医药管理局中药药理科研三级实验室,宜昌 443002)
类风湿关节炎(rheumatoid arthritis,RA)是一种常见的慢性自身免疫性疾病,骨破坏是其最主要的特征之一[1]。骨破坏对关节的进行性损伤不仅会导致后期的功能障碍,还会带来巨大的经济负担[2]。大量破骨细胞(osteoclast,OC)的形成是骨破坏的主要原因,目前的骨保护措施是抑制其分化成熟[3]。
青藤碱(sinomenine,SIN)是在中国传统中药青风藤中提取的一种生物碱单体,是青风藤中的主要有效成分,具有镇痛、抗炎、免疫抑制等多种药理作用,自1921年,Ishiwari首次提取出该成分,用于RA的治疗已有近百年的历史[4-6]。临床研究表明,SIN可有效延缓RA骨破坏的进程,改善关节肿胀状态[7-9];在基础研究中,SIN亦可通过抑制OC分化相关因子和信号通路降低胶原诱导性关节炎(collagen-induced arthritis,CIA)骨破坏[10-12]。活化T细胞核转录因子c1(nuclear factor of activated T cells,cytoplasmic 1,NFATc1)作为OC分化下游的重要转录因子,其可被钙调磷酸酶(calcineurin,CaN)活化进而促进OC分化成熟[13,14]。正常骨处于由成骨细胞(osteoblast,OB)介导的骨形成和OC介导的骨吸收组成的动态平衡。然而,在RA中大量OC的分化成熟导致该动态平衡失衡,进而导致了骨破坏[15]。有研究表明,CaN活化后可增加NFATc1蛋白表达水平加剧骨破坏[16]。此外,CaN基因敲除后可降低NFATc1蛋白表达水平从而延缓骨破坏[17]。提示CaN/NFAT信号通路在RA骨破坏中至关重要。然而,SIN虽可有效抑制RA骨破坏,但与CaN/NFAT的关系尚未阐明。为进一步探讨SIN是否可通过CaN/NFAT信号通路降低RA骨破坏,本研究建立了CIA模型,以期为临床运用SIN治疗RA提供实验依据。
1 材料与方法
1.1材料
1.1.1实验动物 SPF级雄性C57BL/6小鼠60只,体质量(20±1)g,购自湖北省疾病预防控制中心,动物合格证号:SYXK(鄂)2017-0061。实验期间饲养于三峡大学实验动物中心,保持环境恒温恒湿,自由饮水,饲料固定。本研究方案经三峡大学实验动物伦理委员会批准。
1.1.2试剂与仪器 主要试剂:完全弗氏佐剂(complete freund′s adjuvant,CFA)、不完全弗氏佐剂(incomplete freund′s adjuvant,IFA)、牛二型胶原(collagen Ⅱ,CⅡ)购自美国Chondrex公司;SIN购自美国Selleckchem公司;DMSO购自美国Sigma公司;小鼠单抗NFATc1购自英国Abcam公司;兔多抗CaN购自proteintech公司;HRP标记的山羊抗兔IgG、HRP标记的山羊抗小鼠IgG购自谷歌生物有限公司。主要仪器:酶标仪购自奥地利TECAN公司;磁力搅拌器购自上海沪西分析仪器厂;摇床、掌型离心机购自中国其林贝尔公司;Western blot垂直电泳槽、Western blot电转仪购自北京六一仪器厂;化学发光呈像系统购自上海欧翔仪器有限公司;台式高速冷冻离心机购自中国格瑞恩科技有限公司;电子天平购自中国梅特勒-托利多仪器有限公司;脱水机(TP1020)、超薄切片机、倒置荧光显微镜购自德国Leica公司;光学显微镜购自日本Olympus公司。
1.2方法
1.2.1CIA模型的制备 雄性C57BL小鼠60只,小鼠适应性喂养1周后,随机选取6只为正常组,其余小鼠为造模组(排除造模失败小鼠)。根据文献[18],取等体积CⅡ和CFA混合置于5 ml玻璃注射器,反复抽吸30 min(冰上操作),直至混合物完全、充分乳化,以乳化物滴入水中不松散为度,制成抗原。造模小鼠尾根部注射抗原0.1 ml。3周后加强免疫,取CⅡ与等体积的IFA混合注射,造模小鼠尾根部再次注射抗原0.1 ml;正常组不予以任何处理。加强免疫后,剔除发病不明显小鼠。将造模成功的小鼠随机分为模型组(M组)、青藤碱低剂量组(SL组)、青藤碱高剂量组(SH组)各9只。
1.2.2给药方法 初次免疫后第31天起,SL和SH组小鼠给予SIN 30 mg/kg、300 mg/kg灌胃,总共给药12 d,1次/d。C组、M组给予等体积的生理盐水,给药时间与SL组、SH组相同。
1.2.3关节炎指数评分方法 每日观察并记录全身关节病变程度。全身等级按5级评分评价,计算出关节炎指数(arthritis index,AI) 。AI评分标准如下:0分:无红肿;1分:足小趾关节红肿:2分:小趾和足跖关节红肿;3分:踝关节以下的足爪均红肿;4分:包括踝关节在内的整个足爪全部红肿或者出现畸形。四肢评分之和(最高16分)为AI。AI越高,表明关节炎程度越严重[7]。
1.2.4小鼠关节病理切片 小鼠处死后,取双后肢踝关节,于4%多聚甲醛中固定、常规脱钙、脱水、石蜡包埋,切片后,光学显微镜观察病理组织学变化:①炎症细胞:关节有无炎症细胞浸润或纤维化;②滑膜组织:关节滑膜细胞有无变性,滑膜血管数量有无增多;③骨破坏:关节腔有无消失、纤维化,软骨有无破坏或纤维化。HE染色病理切片指标评分标准如下:0分正常;1分轻度;2分中度;3分重度,每只小鼠足爪单个指标得分≤3分[19]。
1.2.5免疫组化 选取小鼠滑膜组织切片,60℃烤箱烘烤1~2 h,常规脱蜡;0.01 mol/L、pH 6.0柠檬酸微波修复后取出玻片置于湿盒中PBS冲洗3次;干燥玻片周围组织,配置3%H2O2室温孵育,PBS冲洗3次(10 min/次);正常山羊血清室温封闭30 min;弃去封闭液,滴加兔多抗CaN(1∶200)、小鼠单抗NFATc1(1∶200),4℃过夜;取出玻片,PBS冲洗3次(10 min/次),滴加辣根过氧化物酶标记的山羊抗兔抗体(1∶200)、山羊抗小鼠抗体(1∶200),室温孵育1 h,PBS冲洗3次(10 min/次);配置DAB显色液,现配现用,镜下调节显色适宜后流水冲洗,双蒸水润洗;苏木精复染细胞核2~3 min;常规脱水、透明、干燥、封片。每组随机抽取3个样本,每个样本选取5个不同部位,于光学显微镜下观察并采集图像,Image-Pro Plus图像分析软件计算阳性率。
1.2.6Western blot检测CIA小鼠踝关节滑膜组织中CaNAα、NFAT的蛋白表达 选取小鼠四肢关节组织20~40 mg制备样品,用BCA蛋白试剂盒进行蛋白定量;10%SDS-PAGE电泳分离,将分离的蛋白转移至PVDF膜,5 %脱脂奶粉封闭1 h,加入兔多抗CaN(1∶1 500)、小鼠单抗NFATc1(1∶250),4℃孵育过夜,TBST充分洗膜3次(10 min/次)。将PVDF膜分别封入山羊抗兔抗体(1∶1 500)、兔抗小鼠抗体(1∶3 000),室温摇床孵育1 h,TBST洗膜3次(10 min/次)。将ECL显影液按照A液:B液=1∶1的比例配置发光液,将其均匀滴在PVDF膜上,放入凝胶成像仪器内显影。以β-actin作为内参,实验重复3次,Image J软件计算各条带灰度值,结果以目的蛋白与内参蛋白之比表示。

2 结果
2.1关节炎评分与关节炎指数变化趋势 加强免疫后每天进行AI评分,关节炎评分标准如图1A所示,0∶0分;1∶1分;2∶2分;3∶3分;4∶4分。统计结果如图1B所示,造模后第37天,与C组相比,M组AI评分明显升高(P<0.01);与M组相比,SL组、SH组AI评分明显降低,且呈剂量依赖性,差异具有统计学意义(P<0.05,P<0.01)。造模后第39天,与C组相比,M组AI评分明显升高(P<0.01);与M组相比,SL组、SH组AI评分明显降低,且呈剂量依赖性,差异具有统计学意义(P<0.05,P<0.01)。造模后第41天,与C组相比,M组AI评分明显升高(P<0.01);与M组相比,SL组、SH组AI评分明显降低,且呈剂量依赖性,差异具有统计学意义(P<0.05,P<0.01)。
2.2HE染色及病理评分 如图2A所示,C组踝关节组织结构清楚完整,关节软骨表面光滑,滑膜细胞无增生、充血、水肿,无炎症细胞浸润,软骨细胞呈椭圆形,排列整齐。M组提示关节软骨及骨组织破坏明显,关节滑膜细胞大量增生,纤维组织增生充血、水肿,大量炎症细胞浸润,关节邻近骨组织也有明显破坏吸收,可见增生的纤维组织向骨髓腔内侵入生长;与M组比,SL组小鼠踝关节炎症细胞浸润减少,软骨及骨组织破坏有所改善,SH组小鼠踝关节炎症细胞浸润明显降低,骨破坏明显减轻,表面光滑。病理评分统计结果如图2B,与C组相比,M组炎症细胞浸润、软骨及骨破坏评分均显著增加,差异具有统计学意义(P均<0.01);与M组相比,SL、SH组炎症细胞浸润、软骨及骨破坏评分均有所降低,差异具有统计学意义(P<0.05,P<0.01)。
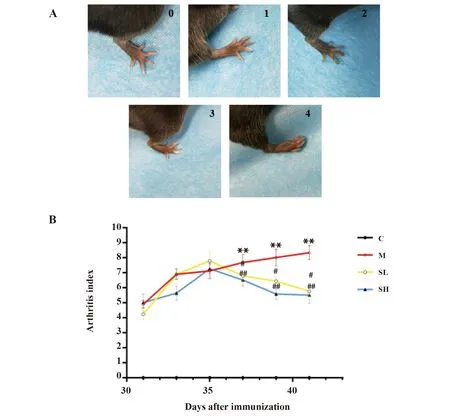
图1 关节炎评分及其变化趋势Fig.1 Arthritis index and its trends of each group


图2 各组病理染色结果及评分(×400)


图3 免疫组化检测CaNAα、NFAT蛋白表达情况(×400)

图4 Western blot检测CaNAα、NFAT蛋白表达情况Fig.4 Protein expressions of CaNAα and NFAT by Western blot
2.3免疫组化检测CaNAα、NFAT蛋白表达 各组结果如图3A所示,阳性表达呈棕黄色点状;与C组相比,M组CaNAα、NFAT表达显著上调(图3B,P<0.01);给予SIN治疗后,与M组相比,SL、SH组CaNAα、NFAT表达显著下调,且呈剂量依赖性(图3B,P<0.01)。
2.4Western blot检测CaNAα、NFAT蛋白表达 与C组相比,M组CaNAα、NFAT蛋白表达显著上调(P<0.01);与M组相比,SL、SH组CaN、NFAT蛋白表达显著下调,差异有统计学意义(图4,P<0.01)。
3 讨论
RA是一种临床常见的自身免疫性疾病,其基本病理特征包括慢性滑膜炎、滑膜增生、血管翳形成及骨和软骨的破坏,导致关节功能障碍等社会负担[20]。2012年流行病学调查指出,我国RA患病率约占总人口的1%,女性略高于男性,并存在区域差异[21]。延缓骨及骨质的破坏,迅速达到低疾病活动状态是目前的主要治疗目标。近期研究表明,OC是导致骨破坏的主要原因,大量细胞因子和信号通路影响其分化成熟[12]。其中,CAN/NFAT信号通路在OC分化过程中发挥重要作用。
CaN是一种Ca2+依赖型丝氨酸/苏氨酸蛋白磷酸酶,其是一种异源二聚体,主要由与CaNB、钙调蛋白(calmodulin,CaM)相结合的催化亚基CaNA和与Ca2+相结合的调节亚基CaNB组成[22]。CaNA由CaNAα、CaNAβ、CaNAγ三个基因编码,但与Ca2+信号相关的是CaNAα基因[23]。细胞外的Ca2+通过其释放活化的钙通道(calcium-release activated calcium,CRAC)流入细胞质,与CaM、CaNB相结合导致CaNA的构象发生变化,彻底激活CaN[24]。未活化的NFATc1一般存在于细胞质中,处于高度磷酸化的状态。被激活的CaN会活化其下游蛋白NFATc1进而入核表达,促进OC分化成熟[25],最终导致大量OC形成,对骨关节产生破坏。Sun等[26]通过应用CaNAα抑制剂及敲除CaNAα基因的方式,发现RAW-C3细胞由于NFATc1表达的降低,分化为OC的能力显著下降[27]。此外,缺乏NFATc1信号激活的破骨细胞前体在RANKL存在的情况下无法完成正常分化。提示NFATc1可能是CaN/NFAT信号通路整合RANKL信号、调控OC终末分化的最重要转录因子。有研究表明,黄芩苷、大麻素受体2激动剂、圣草酚均可通过抑制NFAT的活性降低骨破坏[28-30],提示NFAT与骨破坏密切相关。此外,Liu等[31]研究发现,光色素可通过降低CaN、NFAT的表达减少CIA小鼠骨破坏。Guo等[32]发现,环孢素A亦可通过抑制CaN/NFATc1信号通路活性降低CIA骨破坏。以上研究表明,抑制CaN/NFAT信号通路,从而抑制OC分化,进而阻止关节炎骨破坏至关重要。
SIN是在中国传统中药青风藤提取的一种生物碱单体,是青风藤的主要有效成分,具有良好的抗炎、镇痛、免疫抑制作用及较低的毒副作用[32]。既往研究表明,SIN联合MTX、针刺、乌头原碱均可有效缓解RA患者骨破坏进程,改善关节肿胀[7-9]。此外,SIN可通过抑制OC分化相关信号通路核转录因子κB(nuclear factor kappa B,NF-κB)、丝裂原活化蛋白激酶(mitogen activated protein kinase,MAPK)、janus激酶/信号转导和转录激活因子(janus kinase/signel transducers and activators of transcription,JAK/STAT)降低CIA小鼠骨破坏[33-35]。然而,SIN虽可有效延缓RA骨破坏,但其与CaN、NFAT的调控是否相关尚不清楚。因此,本实验通过建立CIA模型,探讨SIN是否可通过调控CaN/NFAT信号通路来降低CIA骨破坏。
本研究表明SL组和SH组小鼠关节炎症状与模型组相比CIA 显著减轻,SIN在治疗效果上呈现出药物剂量依赖性。通过HE染色和病理评分可以观察到模型组关节组织出现高度异常的病理变化,如血管翳形成和关节软骨破坏等。相反,SL组、SH组表现为滑膜细胞浸润、血管翳形成减少、骨和软骨破坏降低,且呈剂量依赖性。以上结果进一步证实SIN抑制CIA炎症进展和骨破坏。由免疫组化、WB结果可知,CaN、NFAT蛋白表达水平在正常组表达较低,在CIA模型组表达明显,SL组、SH组均可降低CaN、NFAT表达,尤其SH组较为明显。因此,SIN可能是通过抑制CaN/NFAT信号通路,进而抑制OC分化,以此达到治疗关节炎骨破坏的目的。综上所述,CaN/NFAT是OC分化的重要信号通路,SIN可通过抑制CaN/NFAT信号通路活性,减少了OC分化,进而抑制RA骨破坏。此外,在SIN给药期间,小鼠未出现明显不良反应,表明SIN安全性良好。但本实验未使用阳性对照组,还需要进一步实验证实。
