3个纤维颜色不同的棉花近等基因系主茎功能叶生理特性变化与产量因素的相关性分析
2020-07-09张晋龙冯加加渠云芳黄晋玲
张 丽,赵 岩,张晋龙,陈 义,冯加加,渠云芳,黄晋玲
(山西农业大学农学院, 太谷 030801)
天然彩色棉是指棉纤维成熟后具有棕、绿、黄、粉红、蓝等颜色的棉花。研究表明,棉纤维细胞发育大致经历细胞的分化和突起、纤维伸长、次生壁加厚和脱水成熟等4个阶段[1]。彩色棉的形成是棉纤维发育过程中色素物质在中腔细胞中积累所致。利用彩色棉,既能满足人们对不同棉纤维颜色的需求,同时也降低了环境污染的风险。但是,由于天然彩色棉产量较低、品质较差,故市场上商业化种植的仅有棕色棉和绿色棉[2]。华水金等[3]对棕色棉和绿色棉的产量性状与白色棉进行了比较,结果发现,二者的单株铃数、单铃重及衣分较白色棉低。
光合作用是植物生长、发育的基础,作为光合作用的功能器官,叶片产生的有机物是棉株各器官发育的基础,同时也是产量形成的物质基础。棉花叶片和纤维中碳水化合物含量的下降会导致纤维品质降低[4]。叶绿素是植物光合作用中一类重要的色素,叶片叶绿素含量的高低与不同时期光合作用的强弱有关。天然彩色棉叶片的光合色素含量比白色棉低,造成彩色棉光合产量比白色棉低,纤维发育所需物质基础减少,进而影响到产量和品质[5,6]。
碳水化合物的代谢受相关酶类活性的影响,是制约纤维发育的重要因素。蔗糖磷酸合成酶能够调节光合产物在淀粉和蔗糖之间的转化,促使蔗糖合成。而蔗糖合成酶在调控纤维的发育中具有重要作用,其催化蔗糖产生的尿苷二磷酸葡萄糖(uridine diphosphoglucose,UDPG)是合成纤维的直接底物。因此,两种酶活性的强弱直接影响到纤维素的合成。纤维素含量的高低又与棉花纤维的产量和品质密切相关[7]。本研究主要以纤维颜色不同的3个棉花近等基因系为材料,探究整个生育期叶片光合生理生化指标的变化特点,旨在揭示其与产量的关系,为棉花高产品种的培育提供理论依据。
1 材料与方法
1.1 试验材料
本研究选用从三元杂种[(亚洲棉×比克氏棉)×陆地棉]中培育的3个纤维颜色不同的棉花近等基因系为材料,即绿色棉、棕色棉和白色棉。所用材料均由山西农业大学棉花育种实验室提供。
1.2 试验方法
参试材料于2018年在山西农业大学农作站种植。试验采用随机区组设计,处理为3个材料,3次重复,两行区,小区长5 m,行距0.7 m,每行15株。2018年4月25日播种,以常规大田管理方式管理。在棉花生长发育时期,分别于5月30日(苗期)、6月24日(蕾期)、7月22日(花铃期)、9月30日(吐絮期)取相同部位完全展开的功能叶(打顶之前取倒四叶,打顶之后取倒二叶)测定其光合速率及光合相关生理指标。每个材料随机取9株,每3株为1次重复,重复3次。
利用CI光合测定系统(CI-340,USA)测定净光合速率(net photosynthetic rate,Pn)、蒸腾速率(transpiration rates,Tr)、气孔导度(stomatal conductance,Gs)。测定时光合有效辐射(photosynthetically active radiation,PAR)范围为1 000~1 800 nm,系统光强、CO2浓度以及叶室温度分别设置为760 μmol/(m2·s)、0.000 37%及28 ℃;叶绿素含量采用丙酮直接浸提法[8]测定;可溶性糖和淀粉含量采用蒽酮法[8]测定,可溶性蛋白含量采用考马斯亮蓝法[8]测定,以牛血清蛋白为标准品绘制标准曲线,参照高俊凤[8]的方法;酶液提取、蔗糖合成酶测定分别参考Dougla等[9]和Ou-Lee等[10]的方法;蔗糖磷酸合成酶活性测定参考於新建[11]和Wardlaw等[12]的方法。
取100粒籽棉,分别称其籽棉与棉籽的质量,计算籽指、衣指和衣分,设置3次重复。吐絮期,每小区收获第5~10果枝第一、二果节吐絮正常的50个棉铃,晒干称重,计算单铃重。对标记的9株材料,计算单株铃数和单株籽棉产量。
1.3 数据统计与分析
试验数据采用Excel等软件进行处理,用SPSS22.0进行各项测定指标的统计分析和相关性分析。
2 结果与分析
2.1 光合作用相关生理指标动态分析
植物通过光合作用产生干物质和能量[13]来影响其生长发育。光合速率是衡量叶片对CO2同化能力的重要指标之一;气孔导度则反应外界环境中的CO2进入到叶绿体羧化部位的限制程度,与净光合速率的密切程度最高[14,15];而蒸腾速率则对气孔导度的变化具有一定的影响[16]。在棉花的生育期,对绿色棉、棕色棉和白色棉的叶片净光合速率、蒸腾速率、气孔导度进行测定(表1)。由表1可以看出,苗期,2种彩色棉主茎功能叶片的净光合速率均低于白色棉,但差异不显著;蒸腾速率均极显著高于白色棉,棕色棉的气孔导度略高于白色棉。蕾期至吐絮期,2种彩色棉主茎功能叶片的净光合速率、蒸腾速率、气孔导度均低于白色棉,且彩色棉与白色棉之间净光合速率差异达到极显著水平。彩色棉主茎功能叶片较低的净光合速率导致了其合成的光合产物较少,不利于其产量的提高。
2.2 叶绿素含量动态分析
叶绿素是植物叶绿体中一类重要的色素,在光吸收过程中发挥核心作用,是影响光合作用强弱的重要因素,也是衡量叶片衰老的重要指标[17,18],叶绿素含量能够反映植物光合速率的强弱[19]。
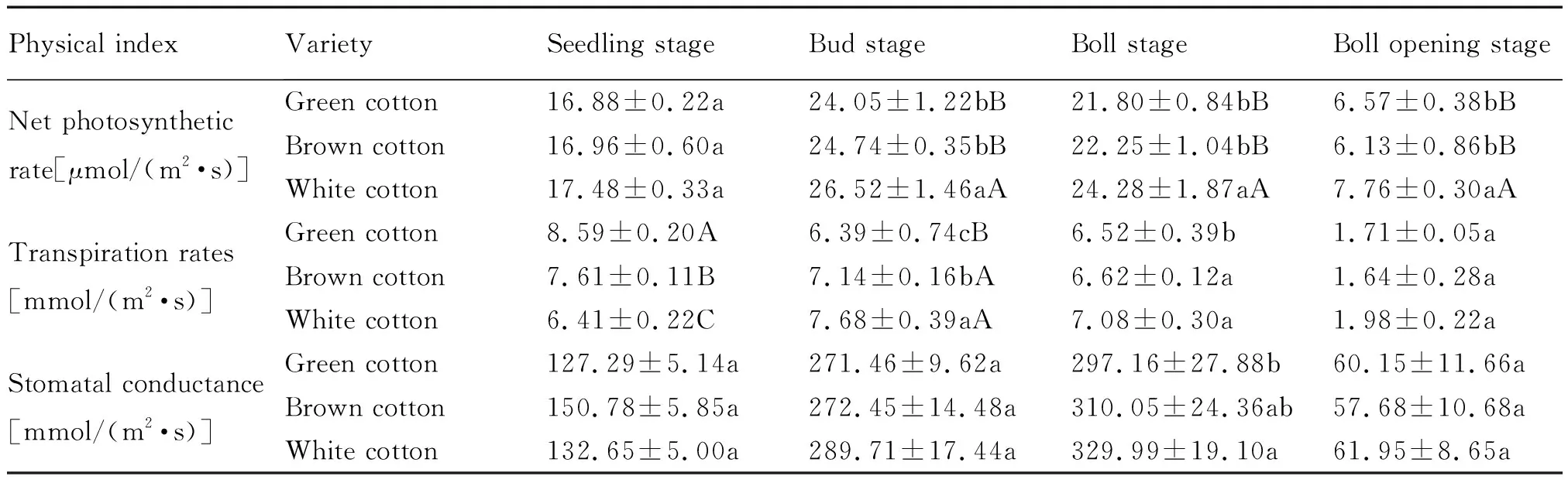
表1 参试材料主茎功能叶片光合生理指标分析Tab.1 Analysis of photosynthetic physiological indexes of functional leaves of main stem of the tested materials
注:小写字母表示5%差异显著水平,大写字母表示1%差异极显著水平。下同
Note:Small letters represent a significant difference of 5% and capital letters represent a extremly significant difference of 1%. The same below
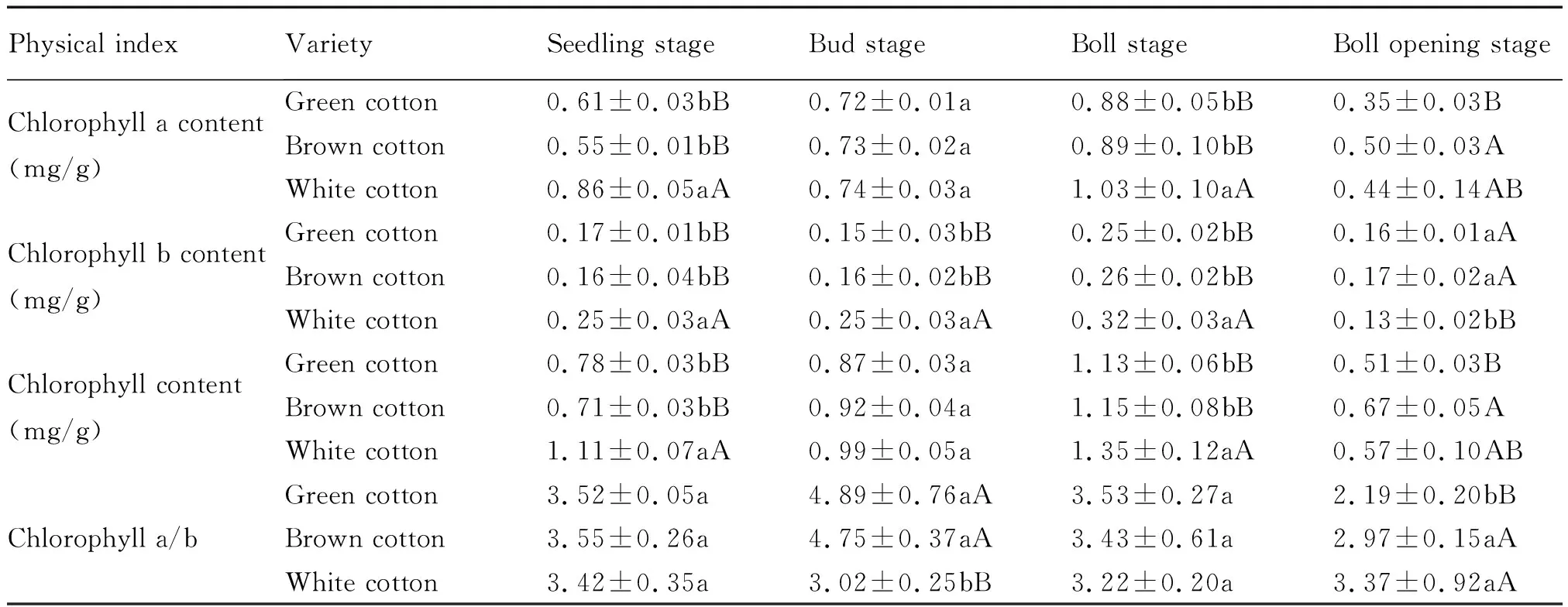
表2 参试材料主茎功能叶片叶绿素含量分析Tab.2 Analysis of chlorophyll content of functional leaves of main stem of the tested materials
由表2可知,除吐絮期外,其他生育期2种彩色棉主茎功能叶片叶绿素a、叶绿素b以及总叶绿素含量均低于白色棉,而叶绿素a/b值均高于白色棉。彩色棉与白色棉之间叶绿素b含量差异在苗期至花铃期达到了极显著水平,叶绿素a与总叶绿素含量差异在苗期与花铃期达到极显著水平,而叶绿素a/b值差异仅在蕾期达到极显著水平。在棉花生长关键时期,彩色棉总叶绿素含量较低,而叶绿素a/b值较高,说明彩色棉主茎功能叶片的光合作用能力较弱,不利于较多光合产物的合成。
2.3 碳水化合物含量动态分析
植物叶片通过光合作用产生的碳水化合物可以通过一系列途径运输到其他部位,供生长发育所需,主要包括糖类、淀粉等。在纤维发育过程中,大量的碳水化合物作为能量物质用来维持各种代谢活动需要[20]。
棉花整个生育期,3种参试材料主茎功能叶片的可溶性糖含量均表现为降低-升高趋势(表3)。除花铃期以外,2种彩色棉主茎功能叶片可溶性糖含量均低于白色棉,其差异分别在苗期与蕾期达到极显著水平。花铃期,棕色棉主茎功能叶片中的可溶性糖含量略高于白色棉,但差异不显著。花铃期为棉花产量形成的关键时期,该时期棕色棉主茎功能叶片中可溶性糖含量较高,推测其向库器官转运量较少,不利于棉铃的发育,最终导致其产量的降低。
3种参试材料主茎功能叶片中淀粉含量变化与可溶性糖含量变化趋势相同(表3)。苗期,棕色棉主茎功能叶片中淀粉含量最高,白色棉居中;其他生育期的2种彩色棉主茎功能叶片淀粉含量均低于白色棉。淀粉是光合碳同化的初级代谢产物,可以为植株的营养生长与生殖生长提供能量物质。在棉花营养生长与生殖生长并进时期(蕾期与花铃期),彩色棉主茎功能叶片淀粉含量较低,说明其最初合成的光合产物较少,不能为彩色棉植株的生长与棉铃的发育提供更多的营养物质,从而影响了其产量的提高。

表3 参试材料主茎功能叶片碳水化合物含量分析Tab.3 Analysis of carbohydrate content of functional leaves of main stem of the tested materials
2.4 蔗糖合成相关酶活性动态分析
蔗糖是光合作用的主要产物,是碳运输的主要形式[21]。蔗糖合成酶(sucrose synthetase,SuSy)与蔗糖磷酸合成酶(sucrose phosphate synthase,SPS)是植物蔗糖代谢过程中极其关键的两个酶,其活性的高低调控着光合产物在蔗糖代谢中的分配比例[22]。
棉花整个生育期,两种彩色棉与白色棉主茎功能叶片的SuSy与SPS酶活性均表现为降低趋势(表4)。除吐絮期外,其他生育期两种彩色棉主茎功能叶片SuSy与SPS酶活性均极显著低于白色棉。在棉花产量形成的关键时期,彩色棉主茎功能叶片中SuSy与SPS酶活性较低,不利于其体内最易运输的碳水化合物-蔗糖的合成,从而不能为棉铃的发育提供足够的养分,最终影响了产量的提高。
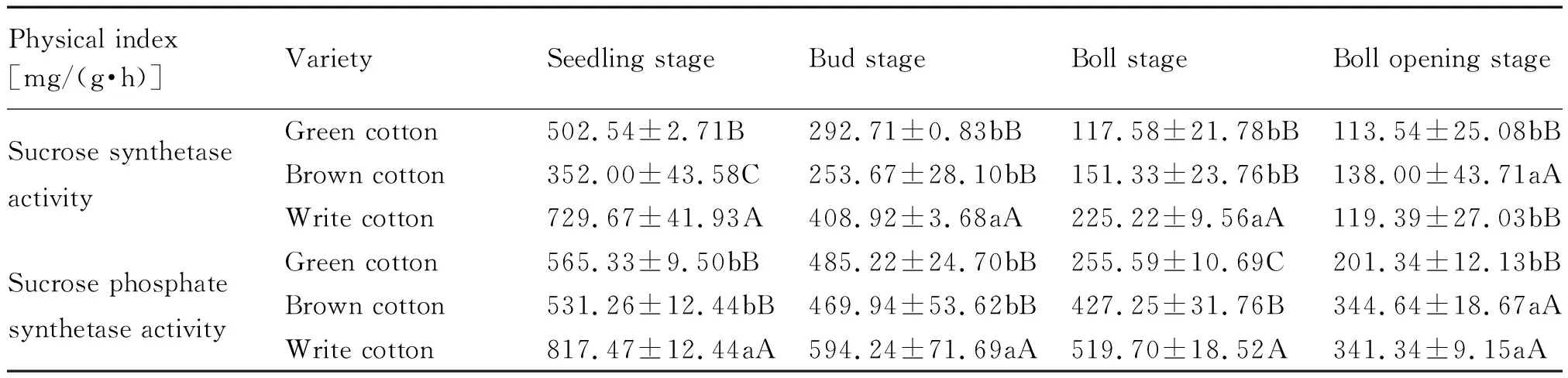
表4 参试材料主茎功能叶片两种蔗糖合成相关酶活性分析Tab.4 Analysis of two sucrose synthetases activity of functional leaves of main stem of the tested materials
2.5 可溶性蛋白含量动态分析
植物叶片的可溶性蛋白中,有很多酶与植物的生长发育密切相关,其含量的多少可以反应叶片中光合产物代谢的强弱[23]。棉花整个生育期,参试材料主茎功能叶片中可溶性蛋白含量均表现为下降趋势(表5)。整个生育期,两种彩色棉主茎功能叶片中可溶性蛋白含量均低于白色棉,且在吐絮期达到显著水平。

表5 参试材料主茎功能叶片可溶性蛋白含量分析Tab.5 Analysis of soluble protein content of functional leaves of main stem of the tested materials
2.6 3种参试材料考种数据分析
从表6中可以看出,2种彩色棉的单株铃数、单铃重、衣指、衣分及单株产量均低于白色棉,籽指均高于白色棉。单株铃数、单铃重、衣分是构成棉花产量的主要因素,2种彩色棉的单株铃数、单铃重与衣分较低,导致了其单株产量也较低。
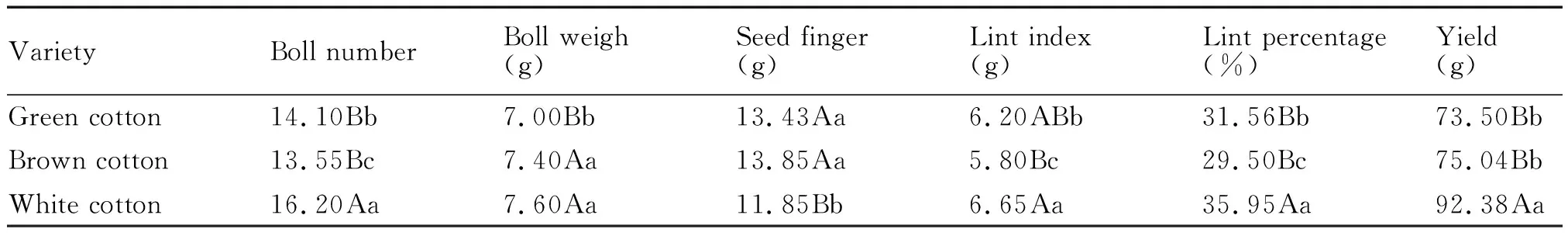
表6 参试材料考种数据分析Tab.6 Data analysis of tested materials
2.7 光合生理指标和产量相关性分析
由于花铃期棉花叶片光合作用最强[24],因此,本研究中以棉株叶片花铃期的光合指标为代表分析了该时期的光合特性与产量的相关性(表7)。结果表明:净光合速率与单株铃数、单铃重、衣指及单株产量显著正相关,与籽指极显著负相关;蒸腾速率与单株铃数、衣分及单株产量显著正相关,与籽指显著负相关;叶绿素a含量与单株产量极显著正相关;叶绿素b含量与单株铃数、衣指及单株产量极显著正相关;总叶绿素含量与单铃重、单株产量极显著正相关;蔗糖磷酸合成酶与单铃重极显著正相关,与单株产量显著正相关;可溶性蛋白与单株铃数、衣分显著正相关,与衣指极显著正相关。由此推测,较高的叶片净光合速率、叶绿素含量及蔗糖磷酸合成酶活性,有利于棉花产量的提高。
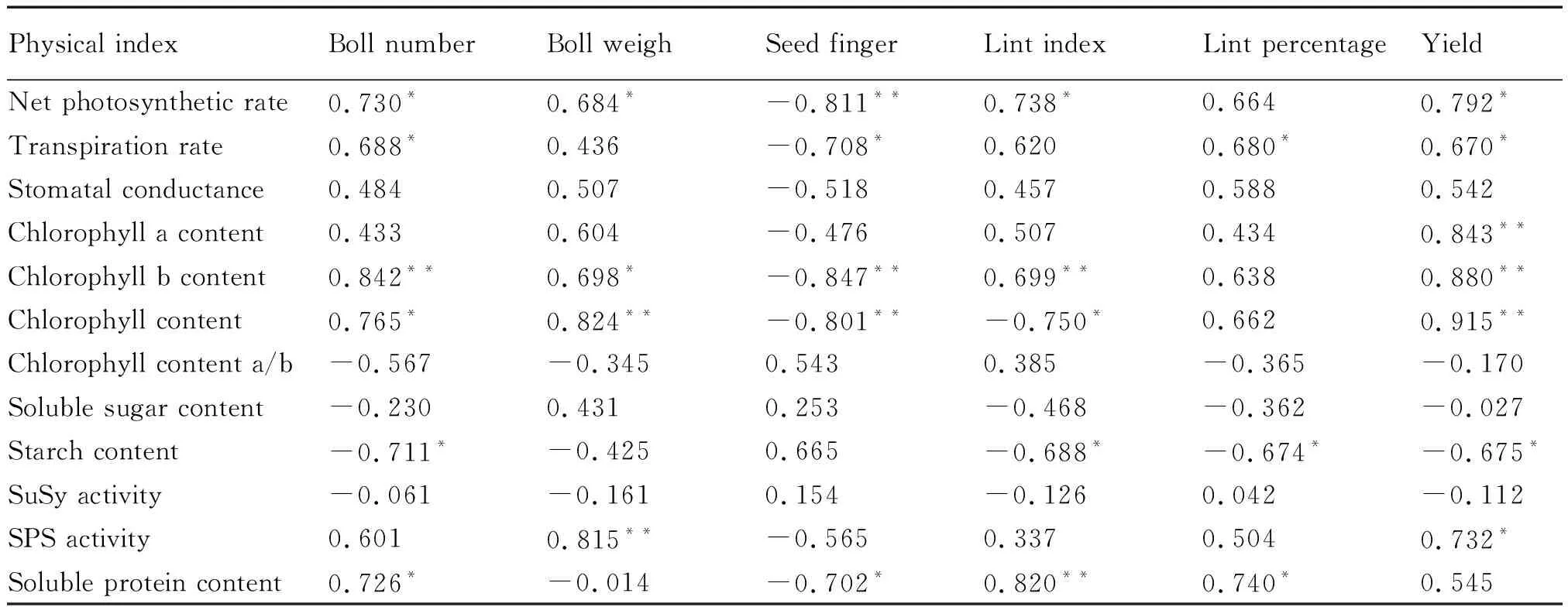
表7 各项指标与产量相关性分析Tab.7 Correlation analysis of various indicators at flowering boll stage with yield indicators
注:*表示在5%水平上差异显著;**表示1%水平上差异极显著
Note:*Indicatesa significant difference at 5% level;** indicates extremely significant difference at 1% level
3 讨论
作物产量与光合速率呈显著正相关[25],较大的净光合速率有利于干物质的积累和最终产量的形成[26],是作物高产的重要生理基础。单株铃数和单铃重是构成棉花产量的主要因素,本研究结果表明,参试材料主茎功能叶片净光合速率与单株铃数、单铃重及单株产量呈显著正相关。在棉花整个生育期,2种彩色棉主茎功能叶片净光合速率均低于白色棉,导致光合作用合成的光合产物比较少,从而影响其单株结铃数和铃重,使得产量较低。由此可以推测,彩色棉主茎功能叶片中较低的净光合速率是导致其产量较低的原因之一。
叶绿素是最重要的光合色素,它的含量直接影响光合作用强度。在本研究中,除吐絮期外,其他生育期2种彩色棉主茎功能叶片的叶绿素a、叶绿素b以及总叶绿素含量均低于白色棉,而叶绿素a/b值均高于白色棉。这表明叶绿素含量大小与净光合速率成正比,而叶绿素a/b值的变化与净光合速率成反比。
棉纤维的形成和积累过程受多种酶的调控,纤维素是棉纤维的主要成分。植物纤维素合成的底物是UDPG,在蔗糖合成酶的作用下,以蔗糖和UDP(uridine diphosphate,UDP)作为原料合成UDPG。SuSy酶在对纤维素合成提供底物UDPG方面起重要的作用,并且有利于增强纤维素合成酶的作用[27]。蔗糖磷酸合成酶是蔗糖进入各种代谢途径的关键酶之一,其主要作用是在光合细胞中促使蔗糖的合成,调节叶片中光合产物在淀粉和蔗糖之间的转化与分配。蔗糖磷酸合成酶活性越高,合成的蔗糖就越多[28],植物中蔗糖磷酸合成酶活性的高低反映了合成蔗糖的能力[29]。Pillonel等[30]发现,UDPG是纤维细胞合成纤维素最好的底物,蔗糖则是棉纤维次生壁合成最好的底物。因此,蔗糖合成酶和蔗糖磷酸合成酶活性与棉纤维的合成与积累密切相关。本研究结果表明:蔗糖磷酸合成酶活性与单铃重呈极显著正相关,与单株产量呈显著正相关,较高的蔗糖磷酸合成酶活性,能促进纤维素等物质的积累,从而促进纤维的生长;除吐絮期外,两种彩色棉叶片中,蔗糖磷酸合成酶和蔗糖合成酶活性均低于白色棉,彩色棉单株铃数与单铃重均比白色棉低。由此推测,棉花生育关键期,彩色棉主茎功能叶片中低活性的蔗糖磷酸合成酶和蔗糖合成酶,阻碍了纤维的形成,是导致其产量较低的原因。
