闽西地区猪流行性腹泻病毒FJLY1703 株基因特征及遗传进化分析
2020-07-02傅云扉戴爱玲李晓华杨小燕
董 波,傅云扉,戴爱玲,李晓华,杨小燕*
(1. 龙岩学院 生命科学学院,福建 龙岩 364012;2. 福建省家畜疫病防治与生物技术重点实验室,福建 龙岩 364012)
猪流行性腹泻病毒(Porcine epidemic diarrhea virus, PEDV)是有囊膜的单股正链RNA 病毒,属于冠状病毒科的α冠状病毒(Alphacoronavirus)属,易感染哺乳期仔猪。临床症状主要有吸收不良、腹泻、脱水、呕吐和消瘦等[1]。当前,PEDV已在全球范围内广泛流行,特别在欧洲、亚洲和美国等国家和地区。2013年,PED疫情席卷整个美国,给其养猪业造成了重大的经济损失,引起了全球关注[2]。PEDV于1984年在我国首次发现,由于其高致病性和高致死率,为中国养猪业带来了持续挑战[3]。2010 年10 月,我国多个省份爆发PED 疫病,大量哺乳仔猪均出现了水样腹泻,脱水和呕吐等典型症状[4-6]。随后,此次疫情迅速蔓延到世界各国,日本、加拿大、墨西哥和哥伦比亚也都经历了持续的PED 疫情暴发[7-8]。据报道,此次PED 的流行与其病原PEDV 的毒力变化有关,即新型PEDV病毒株的出现,威胁着世界养猪业[7]。
PEDV 编码2 个复制酶多聚蛋白ppla 和pplab,1个非结构蛋白ORF3 和4 个结构蛋白:纤突蛋白(S)、膜蛋白(M)、核壳蛋白(N)和小膜蛋白(E)[9]。自20 世纪90 年代以来,我国养猪场已实施定期接种疫苗策略,以控制PED 的流行。然而,采用了PED CV777疫苗株免疫仍然未能阻挡2010 年PED 的大流行,说明当前流行株的毒力增强,传统疫苗未能提供有效保护。在对PEDV 的遗传进化分析中,仅研究其单个结构基因,无法完全确定病毒株的遗传进化特征。基于此,本研究从福建省龙岩市某规模化养猪场的一例确诊为PEDV 感染仔猪小肠样品中经PCR扩增了PEDV 的E、M、N 和S 基因,并对其序列进行了遗传进化分析,同时,对其S 基因进行了重组和相似性分析。为了解闽西地区PEDV 流行株的基因特征和遗传进化规律提供参考依据。
1 材料与方法
1.1 样品来源 2017 年12 月福建省龙岩市某规模化养猪场送检1 例疑似PEDV 感染仔猪,临床表现为水样腹泻、脱水等症状。剖检后,取出其小肠及肠内容物,样品经常规处理提取总RNA 后,经RT-PCR 检测确诊其为PEDV 感染。将鉴定的该病毒命名为闽西FJLY1703 株。
1.2 主要试剂 AxyPrep质粒DNA小量试剂盒、Axy⁃Prep凝胶回收试剂盒,购自康宁生命科学(吴江)有限公司;RNAiso Plus、dNTP、反转录酶M-MLV、5×M-MLV Buffer、pMD18-T 载体,购自宝生物工程(大连)有限公司;DL2000 DNA Marker、10×Loading Buf⁃fer,购自广州瑞真生物技术有限公司。
1.3 引物设计 参考GenBank已登录的PEDV病毒株(AF353511.1)cDNA 序列,设计了7 对引物(表1)用于扩增其E、M、N 和S 基因。引物序列见表1。引物由上海生工生物工程技术服务有限公司(上海)合成。

表1 扩增E、M、N 和S 基因的引物Table 1 Amplification primers for the E,M,N and S genes
1.4 样品中病毒基因的的PCR扩增与其序列分析 从1.1 处理的样品中提取病毒总RNA。反转录为cDNA,以其为模板,利用1.2中的特异性引物PCR扩增4个结构基因,其中S基因采用了4段PCR扩增。PCR 产物经回收纯化后分别连接至pMD19-T 载体;构建的重组质粒均经PCR 鉴定。鉴定阳性的重组质粒由生工生物工程技术服务有限公司(上海)测序鉴定。
1.5 PEDV 结构基因的同源性及系统发育分析 从GenBank下载PEDV的参考株基因序列,将PCR扩增并经测序正确的FJLY1703 株的4 段S 基因片段经拼接完整后与E、M 和N 基因序列分别利用DNAMAN 软件与PEDV 参考株相应基因序列进行同源性分析,并采用MEGA 5.2 软件分别构建这几个基因的系统发育树[10]。
1.6 PEDV S 基因的重组分析 分析FJLY1703 株与本研究中下载的其它PEDV 病毒株S 基因核苷酸序列的相似性和重组事件。利用遗传算法重组检测(GARD)对重组位点进行筛选[11]。使用SimPlot 3.5 对潜在重组事件进行相似性映射和自助扫描分析[12]。
2 结 果
2.1 结构基因的PCR扩增结果 利用特异性引物,以1.4 中 获 得 的PEDV cDNA 为 模 板,PCR 扩 增 其E、M、N 和S 基因的4 个片段。结果显示,获得231 bp、681 bp、1 326 bp、1 031 bp、1 146 bp、1 154 bp 和1 355 bp的目的条带,均与预期相符(图1)。测序结果显示,所获得的基因序列均为PEDV 相应目的基因大小一致(图1)。表明,获得PEDV FJLY1703 株4 个结构基因目的片段。
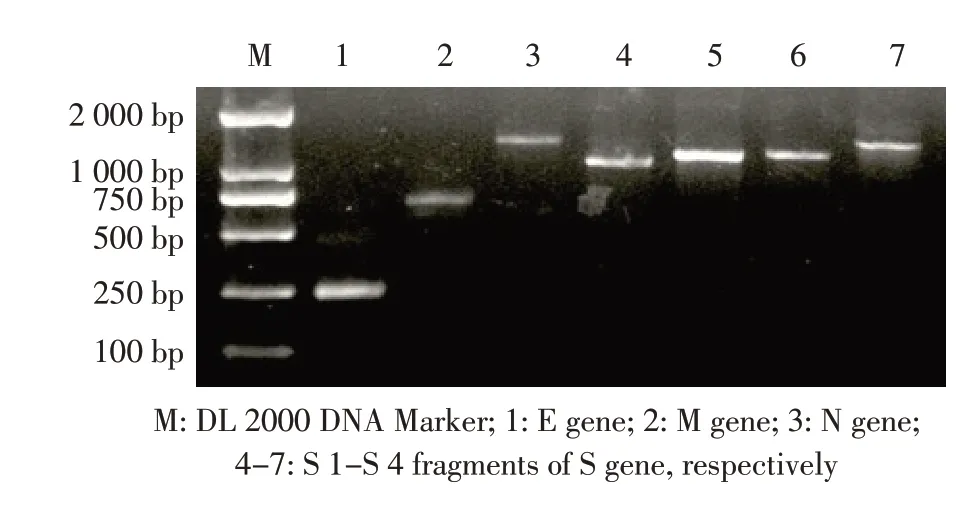
图1 目的基因的PCR 扩增结果Fig.1 Amplification of the target genes by PCR
2.2 PEDV 结构基因的同源性分析 将测序获得的FJLY1703 株的N、E、M 和S 基因分别与GenBank 中登录的疫苗株CV777 等参考株的相应基因序列进行比较,分析其同源性。结果显示,FJLY1703 株N基因核苷酸序列与2010 年后国内PEDV 流行株相应基因序列同源性较高(94.8%~97.8%),然而,其与早期国内外分离株以及疫苗株CV777 相应基因序列的同源性较低(93.7%~94.8%);E 基因序列同源性分析结果显示,FJLY1703 株与2010 年后分离的PEDV 中国株呈现出高度的同源性(95.7%~99.6%),与早期疫苗株CV777 的同源性仅为95.2%;M 基因同源性分析结果显示,FJLY1703 株M 基因序列与2010 年后国内PEDV 流行株相应基因序列同源性为97.1%~99.5%,与疫苗株CV777 等2010 年之前国内流行株相应基因序列同源性较低(96.7%~97.9%);S 基因核苷酸同源性分析结果显示,FJLY1703 株与2010 年之前国内病毒株同源性较低(93.5%~95.0%),与CV777 疫苗株的同源性仅93.9,但与2010 年后国内流行株的同源性为96.0%~97.3%。结果表明,FJLY1703 株主要结构基因均与当前国内流行株同源性较高,而与CV777 疫苗株同源性较低。
2.3 PEDV 结构基因的系统发育分析 N 基因遗传进化分析结果显示,FJLY1703 株属于G2 群G2b 亚群,该群包括多数国内流行株和一株早期韩国株virulent DR13,而CV777 疫苗株属于G1 群。E、M、S 基因的遗传进化分析均显示,FJLY1703 株属于G2群的G2b 亚群,该亚群还包括大多数国内流行变异株,而CV777 疫苗株属于G1 群的G1a 亚群。S 基因遗传进化分析见图2。综上结果表明,FJLY1703 株与当前国内外流行株具有极近的遗传进化关系,而与CV777 等早期经典疫苗株进化关系较远。
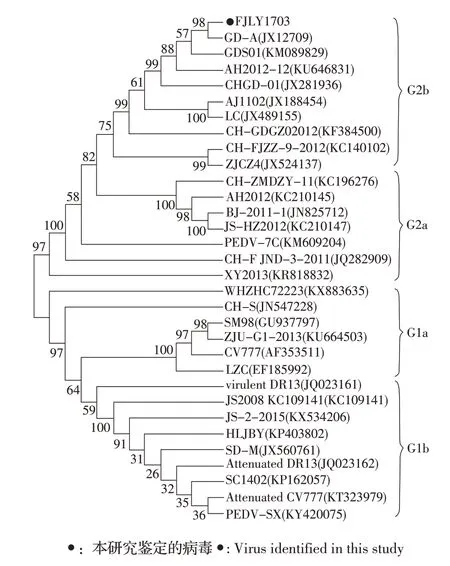
图2 PEDV S 基因系统发育分析Fig.2 Phylogenetic analysis based on S genes of PEDV
2.4 PEDV 结构基因编码的氨基酸序列分析 氨基酸序列分析结果显示,N 基因全长1 326 bp,编码226 个氨基酸,包括一个完整的开放阅读框(ORF)。FJLY1703 株与疫苗株CV777 比较,其N 基因编码的氨基酸序列中存在12 个突变位点(G84A、K122N、 A142T、 N204K、 R241K、 H242L、 K252R、 N255S、L380P、L395Q、Q397L、E400A),这些突变位点与2010年后分离的中国株比较无差异;M 基因编码氨基酸序列与CV777 株比对结果显示,FJLY1703 株存在3个突变位点(E12Q、A42V、A213S),这些突变位点与中国流行株突变位点相似;E 基因编码的氨基酸序列与CV777 株对比结果显示,FJLY1703 株仅存在一处突变(R65Q),这一突变位点与其它2010 年后中国分离变异株的突变位点相似。FJLY1703 株与CV777株在N、E 和M 基因编码氨基酸均存在差异,且这些差异均与国内流行株相同。结果表明,该病毒是一株闽西地区的流行变异株。
PEDV S 基因在病毒侵染细胞中发挥着重要的生物学作用,并且S 基因也是诱导中和抗体产生的结构基因,含有4 个可诱导中和抗体产生的结构域:COE、SS2、SS6 和2C10[13]。FJLY1703 株与经典株CV777 比较,其S 基因编码的氨基酸序列中COE 存在8 个突变位点(I521H、S523G、V527I、T550S、G594S、A605E、L612F、I635V);其SS6 中存在1 个突变位点(Y766S),均与当前国内流行株突变位点相同。上述结果进一步表明,FJLY1703为一株流行变异株。
2.5 FJLY1703 株S 基因的重组分析结果 利用GARD 对FJLY1703 株S 基因进行系统发育不一致和重组事件的比对分析与初步筛选。结果显示,其突变点主要位于S基因5'端1 000 bp~1 500 bp 之间(图3)。通过SimPlotv.3.5.1 评 价S 基 因 核 苷 酸 序 列 的 相 似 性,结果显示FJLY1703 株在S 基因核苷酸序列5'端1 bp~1 200 bp 区域与G1 群病毒株相似性较低,而与G2b亚群以及G2a 亚群病毒株相似性较高,推测这一区域可能来自于G2 群病毒株,即来自PEDV 流行变异株。同时,FJLY1703 株S 基因1 200 bp~4 101 bp 区域与G1 群病毒株相似性高,推测这一区域可能来自于G1 群病毒株,即PEDV 早期经典株(图4)。上述结果表明,FJLY1703 株为一株经典株与变异株重组的闽西地区流行变异株。
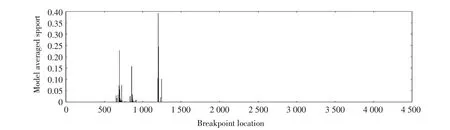
图3 S 基因重组位点的GARD 分析Fig.3 GARD analysis of S gene recombination sites
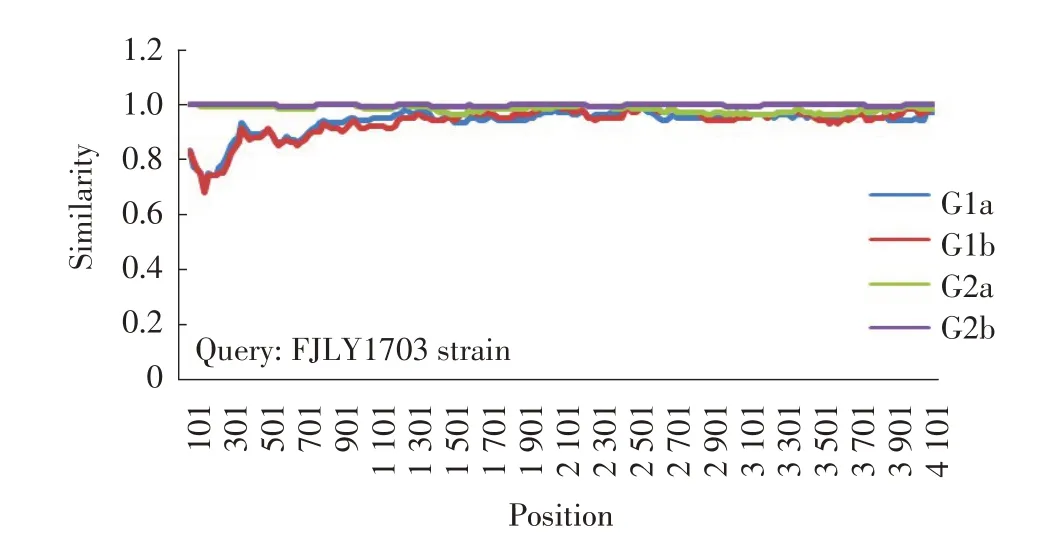
图4 S 基因重组事件分析Fig.4 Recombination analysis of S gene
3 讨 论
本研究结果显示,闽西株FJLY1703 的N、E 和M 基因遗传进化关系与2010 年后大部分中国分离株更为接近,属于当前国内流行变异株。同时,对N基因的系统发育分析显示,早期韩国疫苗株virulent DR13 与FJLY1703 株同属于G2b 亚群,前者是由Vero细胞培养的弱毒株,早期被用于制备疫苗,而FJLY1703 株N 基因与virulent DR13 株遗传进化关系相近,表明FJLY1703 株N 基因可能来自于早期韩国株,这一结果与2010 年以后中国分离株可能起源于早期韩国株的报道相符[13-14]。
与其它冠状病毒相同,PEDV S 基因在病毒侵入和病毒变异中起重要作用,被认为是研究PEDV 遗传特征最有价值的基因[15]。S 基因遗传进化分析显示,FJLY1703 株与其它PEDV 流行变异株均处于G2群,表明这些病毒株同源性较高。该基因序列分析显示,FJLY1703 株S 基因5'区域内存在部分突变,这些变化与其它流行变异株相似。而与CV777 等早期经典株差异较大。N、E、M、S 基因的遗传进化分析均显示,FJLY1703 株与早期疫苗株(CV777)亲缘关系较远,提示以早期经典株CV777 为免疫原所制备的疫苗可能对当前猪群的保护效果不理想,需要选择当前流行株为免疫原研制更为有效的疫苗。
有报道称,PEDV 的重组事件主要发生在经典株和变异株的S 基因,即当前国内流行株基本源于用作疫苗抗原的减毒疫苗株(CV777 或DR13)和高致病性的变异流行株的重组[16-17]。本研究将FJLY1703株S 基因和其它PEDV 病毒株S 基因进行了重组分析。GARD 分析显示,FJLY1703 株S 基因的5'端区域存在突变位点。核酸相似性分析显示,FJLY1703株S 基因5'端1 bp~1 200 bp 区域与2010 年后国 内流行株具有较高的序列相似性,而1 200 bp 以后区域则与CV777 等早期PEDV 经典株相似性高,表明FJLY1703 株S 基因存在基因重组事件,进一步证明FJLY1703 株为一株PEDV 流行株,来自于早期经典株和流行变异株的重组。
氨基酸序列分析显示,FJLY1703株N基因编码的氨基酸序列中存在12个突变位点,其M基因和E基因编码的氨基酸序列中分别存在3处和1处突变,这些突变位点均与2010年后国内流行变异株相似。与CV777株相比,FJLY1703 株S 蛋白的中和表位COE 内存在8个氨基酸突变位点,这些抗原区域的变化,可能会致使其具有更强的致病性。上述结果均表明FJLY1703株为当前闽西地区流行变异株。本研究为了解闽西地区PEDV 地方流行株的基因特征和遗传进化规律提供了基础研究资料。
