自噬在缺氧条件下视网膜色素上皮细胞表达趋化因子IL-8、MCP-1 中的作用
2020-06-19李蓉姚国敏姚杨
李蓉,姚国敏,姚杨
自噬(autophagy,AP)是一种普遍存在于真核生物中的、由溶酶体介导的降解细胞内受损的蛋白质或细胞器的正常代谢过程。作为维持细胞内环境自稳的一种自我保护机制,自噬已成为近年来生物学及生命科学领域的研究热点[1-2]。据报道,细胞自噬对炎症反应有重要的调控作用,而炎症反应也能影响细胞自噬[3]。视网膜色素上皮(retinal pigment epithelium cells,RPE) 细胞是一种具有多种生理生化功能的细胞,病理条件下可分泌包括炎性因子在内的多种细胞因子参与眼部疾病的发生[4]。炎性因子白细胞介素8(interlukin-8,IL-8)和 单核细胞趋化蛋白-1(monocyte chemokine protein 1,MCP-1)属于趋化因子家族,与炎症、机体免疫、肿瘤血管生成和转移等密切相关。目前对于自噬与RPE 细胞功能的相互关系已有报道,但对自噬在RPE 细胞表达趋化因子IL-8 和MCP-1 中的作用尚未见报道。本研究观察细胞自噬在缺氧条件下RPE 细胞表达IL-8 和MCP-1 中发挥的作用,为进一步研究自噬与眼部疾病的关系奠定基础。
1 材料与方法
1.1 材料
ARPE-19 细胞系(武汉普诺赛生命科技有限公司);DMEM/F12 培养基及0.25%胰酶 (美国Gibco公司);3-MA(美国Selleck 公司);CQ(美国Sigma 公司);兔多抗微管相关蛋白1 轻链3 B(microtubule associated protein 1 light chain 3B,LC3B)、兔 多 抗Beclin-1 及兔多抗p62 (美国CST 公司);兔多抗GAPDH(杭州贤至生物有限公司);HRP 标记羊抗兔二抗(武汉博士德生物工程有限公司);人IL-8 和MCP-1 ELISA 试剂盒(欣博盛生物科技有限公司)。三气培养箱 (上海力申科学仪器有限公司,型号HF-100);倒置相差显微镜(日本Olympus 公司,型号IX51);微型高速离心机(美国Labnet 公司,型号C2500-R-230V);全自动酶标仪(美国Thermo scientific 公司,型号Multiskan MK3)。
1.2 方法
1.2.1 细胞处理及分组 用不含EDTA 的0.25%胰酶消化ARPE-19 细胞,用DMEM/F12 培养基 (含10%胎牛血清+1%青霉素和链霉素)终止消化,轻轻吹打混匀成单细胞悬液,传代至培养皿中,在37℃、5%CO2及饱和湿度条件下培养。取处于对数生长期的ARPE-19 细胞(5~10 代)按照每孔5×105个接种于6 孔板内,待细胞贴壁后将细胞分为对照组、缺氧组、3-甲基腺嘌呤(3-methyladenine,3-MA)+缺氧组和缺氧组(简称:3-MA组)和氯喹(chloroquine,CQ)+缺氧组(简称:CQ组),其中对照组在常氧条件下培养;缺氧组置于O2:CO2:N2体积比为1∶5∶94 的培养箱中培养;3-MA组先用10 mM 3-MA 预处理细胞2 h,然后处理方法同缺氧组;CQ组先用50 μM预处理细胞2 h,然后处理同缺氧组。
1.2.2 RPE 细胞IL-8 和MCP-1 的表达 细胞培养及分组方法如前,培养24 h 后将处理好的细胞加入ELISA 试剂盒的96 孔板中,每孔加入100 μL,盖上封纸,置于37℃孵育90 min。孵育结束后,移出液体,不清洗,每孔加100 μL 稀释后的检测试剂A,封口,置于37℃孵育1 h,孵育结束后,移出液体,用350 μL 的洗脱液将96 孔板洗3 次,每次清洗液在孔中停留1~2 min,吸干液体,于每孔加100 μL 稀释后的检测试剂B,封口,37℃孵育30 min,结束后,移出液体,用350 μL 洗脱液将96 孔板洗4 次,洗脱方法如前,吸干液体,于每孔中加100 μL 底物溶液,盖上封纸,孵育15 min,结束后,于每孔加入100 μL 终止液终止反应,酶标仪在450 nm 波长处测量各孔的光密度(OD)值。以IL-8 和MCP-1 标准品浓度为纵坐标,OD 值为横坐标,做直线相关回归,据待测样本的OD 值在标准曲线上查出该样本的IL-8 和MCP-1 浓度对应值。
1.2.3 RPE 细胞自噬相关蛋白的表达 收集各组细胞,用预冷PBS 洗涤3 次,加入含有蛋白酶抑制剂的RIPA 裂解液400 μL,混匀,4℃、1300 r/min 离心15 min,取上清,BCA 法测定蛋白浓度,取50 μg 等量蛋白上样。SDS-PAGE 凝胶电泳,转膜,用含5%脱脂奶粉的TBST 溶液室温封闭2 h,TBST 洗涤3 次,每次10 min。加入一抗LC3B (1:1000)、Beclin-1(1:1000)、p62(1:1000)及GADPH(1:1000,4℃摇 床 过夜,TBST 洗涤3 次,每次10 min,加入辣根过氧化酶标记的二抗(1:50000),室温孵育2 h,洗膜后显色,凝胶成像分析系统扫描分析。以GAPDH 为内参,以目的蛋白/GAPDH 条带灰度值作为其相对表达量。实验进行3 次生物学重复。
1.3 统计学处理
采用SPSS19.0 统计软件对数据进行处理及分析,计量资料以均数±标准差(±s)表示,多组间比较采用方差分析,组间两两比较采用LSD-t 检验,以P<0.05 为差异有统计学意义。
2 结果
2.1 ARPE-19 细胞形态
培养24 h,ARPE-19 细胞呈现扁圆形、梭形、多角形或者不规则形的细胞形态,贴壁良好,胞浆内未见明显黑色素颗粒。不同处理组之间的细胞形态未见明显差异,但3-MA组和CQ组凋亡细胞数量增多(图1)。
图1 4组ARPE-19 细胞在倒置显微镜下的形态(×100)。1A 对照组;1B 缺氧组;1C 3-MA组;1D CQ组
2.2 RPE 细胞中IL-8 和MCP-1 蛋白的表达
ELISA 检测各组ARPE-19 细胞上清液中IL-8、MCP-1 水平,对照组、缺氧组、3-MA组、CQ组的IL-8 浓度(pg/mL)分别为28.98±3.66、129.95±2.29、100.47±4.03 和54.94±3.27;4组细胞的MCP-1 浓度 (pg/mL) 分别为170.33±9.31、526.65±52.43、390.87±44.18 和301.73±4.13。结果显示培养的细胞在正常的情况下仅仅产生少量的IL-8 和MCP-1,在缺氧条件下,细胞表达的IL-8 和MCP-1 迅速增加,而3-MA 或CQ 预处理可以明显抑制IL-8、MCP-1蛋白的分泌(图3)。4组比较,IL-8:F=539.27,P=0.000;MCP-1:F=56.16,P=0.000,差异均有统计学意义。组间两两比较发现,缺氧组的IL-8、MCP-1 浓度明显高于对照组、3-MA组和CQ组,差异均有统计学意义(IL-8:均P=0.000;MCP-1:缺氧组vs.对照组,P=0.000;缺氧组vs.3-MA组,P=0.001;缺氧组vs.CQ组,P=0.000)(图2)。

图2 ELISA 检测四组细胞表达IL-8 和MCP-1 蛋白浓度的比较。2A IL-8 蛋白浓度比较;2B MCP-1 蛋白浓度比较。IL-8:白细胞介素-8;MCP-1:单核细胞趋化蛋白-1
2.3 各组RPE 细胞自噬标记蛋白LC3B-II/I、Beclin-1 和p62 的表达
Westernblot 检测结果显示,对照组、缺氧组、3-MA组和CQ组细胞中的LC3B-II/I、Beclin-1 及p62 的相对蛋白表达量分别为 (0.24±0.02、0.50±0.05、0.34±0.01 和0.67±0.05)、(0.32±0.03、0.97±0.05、0.78±0.06 和0.54±0.01)和(1.03±0.02、0.32±0.10、0.78±0.08 和0.96±0.02)。与对照组比较,缺氧组细胞自噬水平升高,表现为LC3B-II/I 比值、Beclin-1这两种蛋白表达增加和p62 蛋白表达下降。与缺氧组比较,采用自噬抑制剂3-MA 预处理细胞时,LC3B-II/I 和Beclin-1 的表达明显下降,p62 的表达明显增加;采用自噬抑制剂CQ 预处理细胞,Beclin-1 的表达明显下降,LC3B-II/I 和p62 的表达明显增加(图3)。
4组间比较差异,均有统计学意义(LC3B-II/I:F =80.65,P =0.000;Beclin-1:F =139.29,P=0.000;p62:F=70.64,P=0.000)。组间两两比较发现,缺氧组中LC3B-II/I 的相对蛋白表达量明显高于对照组和3-MA组(缺氧组vs.对照组,P=0.000;缺氧组vs.3-MA组,P=0.001);缺氧组中LC3B-II/I 的相对蛋白表达量低于CQ组,差异有统计学意义(P=0.000)。缺氧组细胞中Beclin-1 的相对蛋白表达量均明显高于其它3组(缺氧组vs.对照组,P=0.000;缺氧组vs.3-MA组,P=0.001;缺氧组vs.CQ组,P=0.000)。缺氧组中p62 相对蛋白表达量明显低于其它3组,差异均有统计学意义(均P=0.000)(图4)。
3 讨论
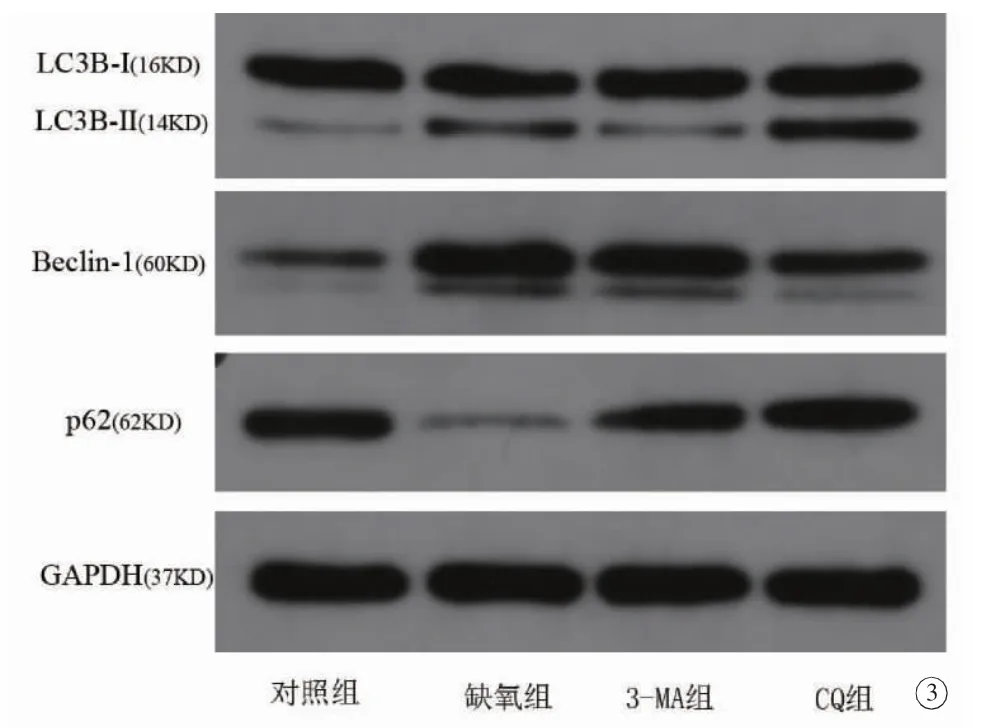
图3 Westernblot 检测四组细胞LC3B、Beclin-1 和p62 的蛋白条带图

图4 四组细胞LC3B-II/ I、Beclin-1 和p62 的相对蛋白表达量比较。4A LC3B-II/I 相对蛋白表达量;4B Beclin-1 相对蛋白表达量;4C p62 的相对蛋白表达量
RPE 是眼底一层重要的细胞结构,其分泌的许多细胞因子对维持眼内微环境具有重要作用。在病理状态下,RPE 细胞可通过分泌IL-4、IL-6、IL-8、IL-10 和MCP-1 等多种炎性细胞因子参与增生性玻璃体视网膜病变 (proliferative vitreoretinopathy,PVR)、年龄相关性黄斑变性(age-related macular degeneration,AMD)及糖尿病视网膜病变(diabetic retinopathy,DR)等的发生和发展[5]。细胞自噬与炎症关系密切,其机制涵盖多个方面[6-7],并且近年来研究表明,细胞自噬在眼部疾病的发生、发展中起着至关重要的作用[3,8-9]。因此,本研究以ARPE-19 细胞分泌的趋化因子IL-8 和MCP-1 为检测指标,探讨自噬是否通过影响RPE 细胞分泌炎症因子而参与眼部疾病的发生。
趋化因子(chemokines)是一类由组织细胞或炎性细胞衍生的、对各种白细胞有趋化活性和激活功能的细胞因子,能选择性地引导白细胞亚类的定向游走和组织内聚集。趋化因子与相应受体结合后主要通过G 蛋白偶联的磷脂酰肌醇途径而发挥免疫炎性和血管生成等多种生物活性。自发现以来,趋化因子与肿瘤的关系一直是研究的热点。趋化因子超家族与相应受体结合可影响多种恶性肿瘤的进展、生物学行为和疾病预后。IL-8、MCP-1 均为具有重要生理作用的趋化因子,在肿瘤的发生、发展、浸润及转移中起着复杂的作用。一般而言,阳性表达者进展快,炎性细胞浸润量高,血管生成密度高,易发生转移,侵袭能力强及预后差[10-11]。IL-8 是Yoshimur 在1987年发现的人类首个趋化因子,目前它是研究最多、促进血管生成作用最强的一种趋化因子[12]。在许多肿瘤组织中均能检测到IL-8 的高表达和分泌,而在相应的正常组织中IL-8 不表达或低表达。研究发现,IL-8 具有诱导肿瘤血管增生,调节血管增殖及渗透性,与多种肿瘤的生长浸润,转移和扩散有关[12-13]。MCP-1 是主要的吸引单核/巨噬细胞到炎症、肿瘤部位的趋化因子,一方面能介导局部的免疫炎性反应,即趋化和激活单核/巨噬细胞,使之产生炎性细胞因子并放大这一过程;另一方面能够诱导和刺激病理性血管生成[14-15]。本研究发现,在正常的情况下,RPE细胞仅产生少量的IL-8 和MCP-1。在缺氧条件下,细胞表达的IL-8 和MCP-1 迅速增加,与文献报道的结果相似[5]。随着近年来对肿瘤血管生成的广泛研究,人们发现各种多种生长因子都具有血管生成的功能。除内皮细胞生长因子VEGF 家族和血管生成素家族之外,血管生成相关性趋化因子和细胞因子被认为其有很强的促血管生成活性。因此,缺氧促进RPE 细胞表达IL-8 和MCP-1,可能进一步参与眼部新生血管的形成。
作为普遍存在于真核生物细胞内的一种机制,细胞自噬能对内环境稳定进行自我维持,且在胚胎发育、细胞自我保护和生存等过程中发挥关键作用。当细胞处于应激状态时,细胞的自噬水平会在短时间内迅速升高。自噬的发生需要多种自噬相关蛋白参与,任何一种自噬基因的缺失或突变都会影响自噬相关进程,从而造成各种眼病的发生[16],例如白内障,AMD,青光眼,葡萄膜炎,视网膜疾病和眼肿瘤等。作为自噬标记蛋白,LC3,Beclin-1 和p62 等被广泛用于自噬的相关研究中,在研究中常结合起来综合判断自噬活性的强弱[17-19]。已有研究发现自噬相关蛋白在视网膜色素上皮层、视网膜神经节细胞层、内核层、外核层均有强烈表达[20],当自噬调节处于失调状态时会导致相关的视网膜疾病的发生。本研究发现,在不同培养条件下,RPE 细胞中LC3、Beclin-1及p62 蛋白都有不同程度的表达,与文献报道相符。另外,鉴于不同自噬抑制剂的作用机制存在差异,本研究采用了两种自噬抑制剂对RPE 细胞进行了干预。其中,3-MA 是Class III PI3K 的抑制剂,它作用于自噬诱导阶段,可特异性阻断自噬体的形成[21]。CQ是一种溶酶体自噬抑制剂,可阻止自噬体与溶酶体的结合,抑制自噬的降解,使得自噬体在细胞内增加,作用后使LC3-Ⅰ向LC3-Ⅱ的转换增加更明显[22]。本研究发现在缺氧条件下,RPE 细胞的LC3B-II/I(LC3B 是LC3 的主要亚型)、Beclin-1 蛋白表达增加及p62 蛋白表达下降,而3-MA 或CQ 处理后这3种自噬关键蛋白的表达明显发生改变,证实缺氧诱导了RPE 细胞的自噬激活。
缺氧刺激可促进RPE 细胞IL-8 和MCP-1 表达,3-MA 或CQ 处理后减少了二者的表达。3-MA和CQ 预处理得到的结果一致,提示抑制自噬能减少RPE 细胞表达IL-8 和MCP-1。结合以往的研究结果,即缺氧条件下自噬激活可促进视网膜血管生成过程[23-24],提示自噬激活促进RPE 细胞分泌趋化因子IL-8 和MCP-1 可能是自噬参与视网膜和脉络膜新生血管形成的另一重要途径。目前,通过调控自噬进程治疗眼部疾病已成为一种崭新的思路,故更深入了解自噬对RPE 细胞分泌IL-8 和MCP-1 的具体调控机制很有可能为治疗视网膜和脉络膜新生血管等眼病提供新方法。
