趋化因子受体5 Δ32 突变在中国20个民族群体中的分布*
2020-06-03陶玉芬史磊姚宇峰史荔刘舒媛
陶玉芬, 史磊, 姚宇峰, 史荔, 刘舒媛
(中国医学科学院 & 北京协和医学院 医学生物学研究所, 云南 昆明 650118)
趋化因子受体5(chemokine receptor 5,CCR5)是一种G蛋白偶联受体[1],CCR5主要表达于巨噬细胞、单核细胞、记忆性T细胞和树突状细胞表面,其在病原感染后调节免疫细胞的激活和迁移中起到了重要的作用[2]。研究发现,CCR5是人和猴免疫缺陷病毒(human and simian immunodeficiency virus,HIV and SIV)的主要辅助性受体之一,CCR5基因位于人染色体3q21.3。迄今为止,在CCR5基因上发现了大约96种突变[3],其中以CCR5Δ32突变的研究最为广泛。CCR5Δ32(即rs333)是指CCR5基因编码区第185位氨基酸密码子之后(554nt-585nt)发生了32个碱基的缺失,导致开放读码框(open reading frame, ORF)错位,造成了翻译的提前终止,在细胞膜表面产生截短的、无功能的穿膜蛋白,从而使的HIV-1的包膜蛋白gp-120不能与CCR5有效结合,从而阻止了HIV-1进入细胞[4]。携带CCR5Δ32突变纯合子的个体对HIV-1感染有明显的抵抗性,携带CCR5Δ32杂合子个体也表现出明显的获得性免疫缺陷综合症(acquired immune deficiency syndrome, AIDS)进程减慢[5]。有研究报道,通过移植携带CCR5Δ32突变纯合子的造血干细胞后,先后有2例AIDS患者体内的HIV病毒清除[6],另外也有使用CCR5拮抗剂如Maraviroc对AIDS进行治疗的方案,可见,CCR5已成为治疗AIDS研究的最佳靶点之一。在全世界范围内CCR5Δ32突变的分布具有明显的地理特征,不同群体中的分布频率差异很大:在非洲、美洲印第安人、东亚人及南亚人中频率极低或者缺乏,欧洲人群CCR5Δ32突变的平均频率为10%,呈现出由北到南逐渐降低的趋势[7]。本文旨在观察CCR5Δ32突变在中国20个不同民族群体中的分布情况,了解CCR5Δ32突变在中国人群中的扩散特征。
1 对象与方法
1.1 研究对象
根据“知情同意”的原则,纳入中国20个不同民族群体,共计2 788例健康无关个体作为研究对象(表1),所有研究对象自述3代之内均来自同一民族群体,年龄30~75岁。
1.2 方法
1.2.1DNA提取 采集研究对象外周静脉血2 mL,EDTA抗凝,用AxyPrep血基因DNA组试剂盒(Axygen,中国)提取基因组DNA,并用超微量紫外分光光度计(ND-2000,美国ThermoFisher公司)检测DNA的浓度和纯度。
1.2.2CCR5Δ32等位基因检测 参考文献设计CCR5Δ32等位基因的扩增引物:正向引物为5′-CTCGGATCCACCAGATCTCAAAAAGAAGGTCT-3′,反向引物为5′-CTCGTCGACATGATGTGTAAGATAAGCCTCAC-3′(引物由Invitrogen公司合成)。10 μL PCR扩增反应体系包含2×PrimeSTAR MAX Premix (TaKaRa,大连) 1 μL、20 ng基因组DNA及每条引物10 pmol/L。PCR扩增条件:98 ℃预变性5 min,98 ℃变性10 s、55 ℃退火5 s、72 ℃延伸5 s,35个循环;72 ℃延伸5 min。PCR产物经2.5%琼脂糖凝胶电泳,溴化乙啶染色,紫外凝胶电泳成像系统成像并分析结果。根据PCR产物的大小和NCBI中报道的CCR5基因的序列,检测个体的基因型判定:野生型纯合子(rs333 wt/wt)为一条242 bp的片段、突变型纯合子(rs333 mt/mt)为一条210 bp的片段、杂合子(rs333 wt/mt)包括242 bp和210 bp的2条片段。对研究对象的CCR5Δ32(rs333-mt)等位基因进行计数,并计算各群体CCR5Δ32突变的频率。
1.2.3CCR5Δ32等位基因的测序验证 PCR产物分别使用QIAquick PCR Purification Kit (Qiagen,德国)PCR产物纯化回收试剂盒对CCR5Δ32等位基因的PCR产物进行纯化回收,并以PCR引物(F: 5′-CTCGGATCCACCAGATCTCAAAAAGAAG
GTCT-3′)作为测序引物,使用BigDye V3.1(ABI,美国)测序试剂盒进行测序反应,并在ABI3730测序仪上进行序列测定。测序结果用DNAstar软件的Clustal W模块进行序列比对,参考序列为GRCh38.p12(NC_000003.12)。
2 结果
2.1 CCR5 Δ32等位基因的PCR产物电泳
2.5%琼脂糖凝胶电泳结果显示(见图1),野生型纯合子(rs333 wt/wt)为一条242 bp的片段、杂合子(rs333 wt/mt)为242 bp和210 bp的2条片段,本研究中未检测到突变纯合子(rs333 mt/mt)。
2.2 CCR5 Δ32等位基因的测序验证
分别将等位基因rs333-wt和rs333-mt进行测序验证(见图2),测序结果用DNAstar软件的Clustal W模块进行序列比对,发现rs333-mt等位基因与rs333-wt等位基因相比缺失了32个核苷酸,缺失序列为GTCAGTATCAATTCTGGAAGAATTTCCAGACA。
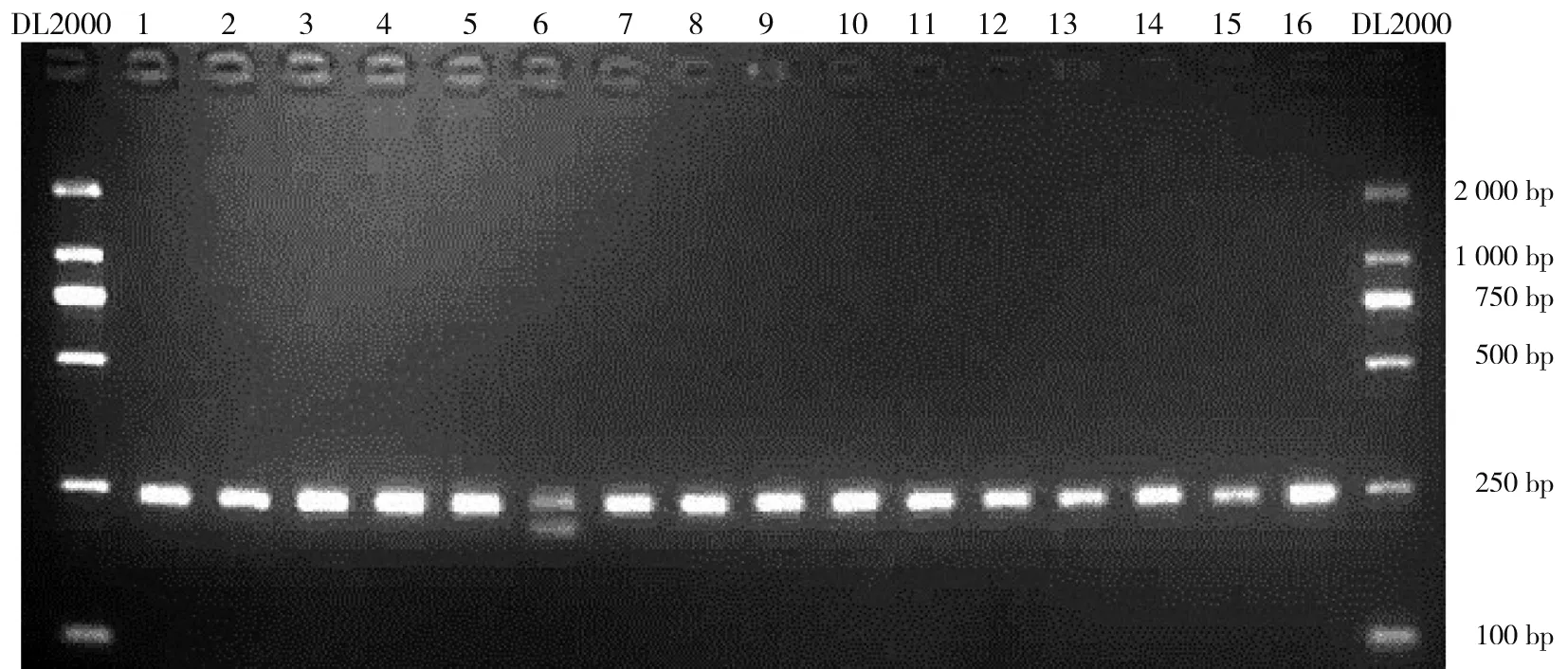
注:DL2000为DNA分子标记,第6泳道为杂合子(rs333 wt/mt),其余泳道为野生型纯合子(rs333 wt/wt)的PCR扩增产物。
图1CCR5Δ32等位基因的PCR产物进行2.5%琼脂糖电泳结果
Fig.1 Results of 2.5% agarose gel electrophoresis on PCR products ofCCR5Δ32 alleles

注:rs333wt为野生型等位基因,rs333 mt为突变等位基因。
图2CCR5Δ32(rs333)等位基因测序结果
Fig.2 Sequencing results ofCCR5Δ32 (rs333) allele
2.3 中国20个不同民族CCR5Δ32基因突变
检测中国20个不同民族共计2 788个健康个体的CCR5Δ32基因突变情况。结果发现,在鄂温克族检测到1例CCR5Δ32基因突变杂合子(rs333 wt/mt),CCR5Δ32突变频率为1.25%;回族(宁夏)检测到1例CCR5Δ32基因突变杂合子(rs333 wt/mt),CCR5Δ32突变频率为0.68%;塔吉克族检测到1例CCR5Δ32基因突变杂合子(rs333 wt/mt),CCR5Δ32突变频率为1.11%;云南汉族人群中检测到2例CCR5Δ32基因突变杂合子(rs333 wt/mt),CCR5Δ32突变频率为0.04%;其余群体中均未检测到CCR5Δ32基因突变。见表1。
3 讨论
CCR5是主要表达于巨噬细胞、单核细胞、记忆性T细胞和树突状细胞表面的一种跨膜G蛋白偶联受体,与其结合的配体为人巨噬细胞炎性蛋白-1(macrophage inflammatory protein-1α and 1β, MIP-1α, MIP-1β),调节激活正常T细胞表达和分泌的细胞因子(regulated on activation normal T cell expressed and secreted factor, RANTES),单核细胞趋化蛋白(monocyte chemotactic protein 1, MCP-1, MCP-2, MCP-3 and MCP-4)[8],CCR5在病原感染后诱导炎性细胞活化、驱化、增殖和免疫调节中起到了重要的作用[2]。
本研究对中国20个不同民族群体的2 788例健康个体进行了CCR5Δ32突变检测,观察CCR5Δ32突变在中国不同地区人群中的分布情况,有助于了解该基因位点在中国人群中的分布扩散。结果显示,在鄂温克族CCR5Δ32突变频率为1.25%、回族(宁夏)CCR5Δ32突变频率为0.68%、塔吉克族

表1 2 788例研究群体及CCR5 Δ32突变情况Tab.1 2 788 study groups and CCR5 Δ32 mutations
CCR5Δ32突变频率为1.11%、云南汉族CCR5Δ32突变频率为0.04%,其余群体中CCR5Δ32突变频率为0.00%。本研究结果与已报道的中国不同民族群体的CCR5Δ32突变频率一致:喀什地区维吾尔族CCR5Δ32频率为3.48 %[9],新疆伊犁地区维吾尔族HIV感染高危人群为4.47%[10],蒙古族为1.12 %[11];而中国南方的民族群体CCR5Δ32突变频率却很低,德宏地区傣族和景颇族[12]、广西壮族[13]、深圳汉族[9]等CCR5Δ32频率均为0%,四川彝族为0.84%[14]。研究显示,除人类外未发现其他非人灵长类动物存在CCR5Δ32突变[15],这表明CCR5Δ32是进化过程中的新发突变。在全世界范围内CCR5Δ32突变的分布具有明显的地理分布特征,在欧洲群体中,CCR5Δ32的频率呈现出由北到南逐渐降低的趋势:欧洲北部阿什肯纳基犹太人(Ashkenazi Jews)的CCR5Δ32频率高达20.93%[16],另外,俄罗斯的Ugro-Finnic人群的CCR5Δ32频率也很高(15%~18%)[17],在欧洲中部的一些国家则频率开始下降,西班牙北部人群频率为8.16%、斯洛文尼亚为8.7%、克罗地亚为7.1%,而欧洲南部CCR5Δ32频率相对较低(2%~5%)[17]。对亚洲人群的研究发现,亚洲人群CCR5Δ32的突变频率很低,泰国人群[18]、印度北部人群[19]都未检测到CCR5Δ32突变。在非洲、美洲印地安人中CCR5Δ32突变的频率也极低或者缺乏[7]。关于CCR5Δ32突变的起源和蔓延最经典的是维京假说认为CCR5 Δ32出现于公元1000-1200年前斯堪的纳维亚,向北传入冰岛,向南传入中欧和南欧,向东传入俄罗斯。中国的汉族群体和南方地区的各民族的CCR5Δ32突变频率很低,但北方的一些群体如维吾尔族等却有相对高的CCR5Δ32突变频率。前期对不同人群遗传结构的研究显示,中国南北民族群体之间具有显著的遗传差异,新疆维吾尔族(阿尔泰语系,突厥语族)、塔吉克族(印欧语系,伊朗语族)等西北人群是由东亚人与迁徙而来的高加索人混合而成[20-21],遗传结构分析显示维吾尔族人群有30%~55%的欧洲人遗传成分[22],鄂温克族(阿尔泰语系、满-通古斯语族)和回族(汉藏语系、汉语族)也是典型的北方群体,可能这些群体的CCR5Δ32突变是来源于欧洲,因此基因频率由北向南逐渐降低。另外,对汉族人群HLA基因多样性的研究发现,云南汉族群体介于北方群体和南方之间,因此云南汉族群体遗传结构兼具了南北方的特征[23]。另外,关于CCR5Δ32突变的起源和频率有选择压力假说认为,CCR5Δ32突变起源于约800年前,经过一个很强的古老选择压力而被保留下来,这一选择作用可能为黑死病(鼠疫)。在公元1346-1352年间,欧洲爆发了广泛的黑死病,这场黑死病造成了欧洲2 500万人死亡,这种选择压力导致了欧洲人的后裔中CCR5基因发生了遗传漂变(欧洲人群CCR5Δ32突变平均频率为10%)[7,24]。也有学者提出,天花病毒对人群也具有较强的选择作用导致欧洲后裔CCR5Δ32突变频率的升高。中国不同地区的人群除遗传背景差异之外,南北方不同的病原选择压力也是造成南北方基因突变频率差异的原因。
研究发现CCR5与多种病原体的感染与关联,如HIV、痘病毒(poxvirus)[25]、流感病毒(influenza)[26]、巨细胞病毒(cytomegalovirus)[27]、单纯疱疹病毒(herpes simplex virus, HSV))[28]、西尼罗河病毒(Nile virus)[29]以及蜱媒脑炎病毒(tickborne encephalitis virus)[30]等。CCR5基因变异也与许多病原感染的易感性相关,如HIV感染、丙型肝炎病毒(hepatitis C virus, HCV)感染[31]、乙型肝炎病毒(hepatitis B virus, HBV)感染、结核杆菌(tuberculosis)感染[32]等。其中CCR5在HIV感染及其在艾滋病进程中作用的研究最为广泛。研究发现,CCR5是HIV-1入侵集体细胞的主要辅助受体之一,CCR5的N-端和胞外结构域2(extracellular loop domain, ECL2)在HIV-1感染时起着关键作用,N-端Tyr-3、Tyr-10、Tyr-14及Tyr-15四个酪氨酸残基的硫酸化有助于CCR5与HIV包膜蛋白复合物gp120/CD4结合,ECL2是HIV-1包膜蛋白gp120与CCR5相互作用的关键区域[3, 33]。CCR5Δ32突变是CCR5第185位氨基酸之后(554nt-585nt)发生了32 bp碱基的缺失,具有CCR5Δ32纯合性突变的基因在翻译过程中会导致阅读框错位,从而导致在淋巴细胞膜表面产生截短的、无功能的CCR5跨膜蛋白,从而使HIV-1的gp-120不能与CCR5 Δ32有效结合,导致HIV-1病毒不能进入宿主细胞[7]。对于CCR5Δ32杂合子来说,虽然它无抵抗病毒感染的作用,但却可以明显推迟艾滋病的进程,原因可能是CCR5Δ32杂合子会减少CCR5在细胞表面的表达,从而减少进入体内的病毒量,并在慢性感染早期,减少病毒复制,明显减少HIV-1病毒RNA在血清或血浆中的含量[34]。近年研究发现,真核生物基因表达的调控的一个重要方面是通过无义介导的mRNA降解(nonsense-mediated mRNA decay,NMD)途径识别含有提前终止密码子(premature translation termination codon,PTC)的异常转录产物,并快速降解异常的mRNAs以防止有害的截短蛋白(truncated proteins)的产生。Ashton等[35]发现,人CCR5 mRNA序列第407位核苷酸之后存在一个程序化核糖体移码(programmed -1 ribosomal frameshift, -1 PRF)信号序列,该-1 PRF可引导翻译核糖体识别CCR5 mRNA上的PTC,进而启动NMD途径对异常CCR5 mRNA进行降解,从而降低CCR5的表达量;同时CCR5 mRNA序列上的-1PRF也可通过miRNA-1224降低CCR5的表达水平。由此推测,含有CCR5Δ32突变的mRNA也可能被NMD所降解,从而降低淋巴细胞膜表面CCR5的表达量,导致无法发挥HIV辅助受体的功能。也有研究发现CCR5Δ32突变蛋白有利于机体自行清除HCV肝炎引发的炎症效应[36]。也有研究发现,CCR5Δ32可以增加HBV感染后的恢复和降低大约50%的HBV慢性感染后的病程发展[7]。
终上所述,本研究发现CCR5Δ32突变在中国不同民族群体中较为罕见,总体上说CCR5Δ32基因突变在北方群体分布较多,南方群体中频率几乎为0,CCR5Δ32突变频率符合“北高南低”的趋势。本研究结果有助于了解CCR5Δ32突变在中国不同地区人群中的分布情况,为不同人群遗传背景和病原驱动的选择压力研究提供一定的数据。
