Wx与ALK主要等位基因不同组合对稻米品质的影响
2020-05-30陈专专杨勇冯琳皓孙晔张昌泉范晓磊李钱峰刘巧泉
陈专专 杨勇 冯琳皓 孙晔 张昌泉, 2 范晓磊, 2 李钱峰, 2 刘巧泉, 2,*
与主要等位基因不同组合对稻米品质的影响
陈专专1杨勇1冯琳皓1孙晔1张昌泉1, 2范晓磊1, 2李钱峰1, 2刘巧泉1, 2,*
(1扬州大学 农学院 植物功能基因组学教育部重点实验室/江苏省作物基因组学和分子育种重点实验室, 江苏 扬州 225009;2扬州大学 江苏省作物遗传生理重点实验室/江苏省粮食作物现代产业技术协同创新中心, 江苏 扬州 225009;*通信联系人, E-mail: qqliu@yzu.edu.cn)
【】稻米蒸煮和食味品质主要受和两个主效基因的调控。这两个基因在栽培稻中存在多个复等位变异,本研究旨在明确和基因的主要等位变异及其组合对稻米品质的影响。将的3个主要等位基因(Wx、Wx和)与的2个主要等位基因(ALK和ALK)进行不同的组合,在粳稻品种日本晴(Nip)背景下创建6个含有和基因不同等位变异组合的近等基因系。通过测定近等基因系的稻米理化品质明确不同等位基因及其组合对稻米品质的效应。在相同的基因型背景下,含Wx等位基因稻米的糊化温度极显著低于型和Wx型的近等基因系。稻米黏滞性受和等位基因组合影响较大,等位变异主要影响稻米的峰值黏度、峰值时间和冷胶黏度;在相同背景下,等位基因主要影响稻米RVA谱的起浆温度,对稻米崩解值和消减值的影响不显著。总体来看,Nip-Wx/ALK和Nip-Wx/ALK型稻米具有较高直链淀粉含量和崩解值,较低的胶稠度,因而食味值较差;Nip-/ALK和Nip-Wx/ALK型水稻具有适中的直链淀粉含量、较软的胶稠度以及较低的糊化温度,因此这两个近等基因系热饭和冷饭的食味值较好;虽然Nip-Wx/ALK的糊化温度较高,但因其适中的直链淀粉含量和较高的胶稠度,其冷饭食味值降幅较小;Nip-/ALK稻米的热饭食味值与Nip-/ALK、Nip-Wx/ALK和Nip-Wx/ALK无差异,但因其较高的糊化温度,其冷饭食味值显著下降。基因变异主要影响稻米直链淀粉含量和胶稠度,对糊化温度有一定的影响;基因变异主要影响糊化温度,对直链淀粉含量无显著影响,对胶稠度存在微效影响。
蜡质基因;糊化温度基因;等位基因;近等基因系;稻米品质
淀粉是稻米胚乳的主要成分,约占水稻种子总质量的80%~90%[1],因而淀粉含量和理化特性极大地影响稻米品质。淀粉的理化特性主要包括直链淀粉含量、糊化温度、胶稠度和快速黏度分析(rapid visco analyzer, RVA)特性等,这些指标可以直观地反映稻米食用及蒸煮品质的优劣[2-3]。其中,直链淀粉含量是由基因编码的颗粒结合淀粉合成酶GBSSⅠ(granule bound starch synthesisⅠ)调控的,基因的等位变异会导致水稻胚乳中直链淀粉含量的差异[4-6];编码可溶性淀粉合成酶SSⅡa (soluble starch synthesisⅡa),该基因的等位变异是导致品种间稻米糊化温度差异的主要原因[5,7]。同时,和也是控制稻米胶稠度和RVA特性的主效基因[5]。鉴于和及其等位变异对稻米品质的重要影响,明确水稻和基因主要等位变异组合对品质的效应,能为稻米品质的改良提供重要的实践指导意义。
栽培稻中基因存在多个复等位变异,包括Wx、Wx、Wx、Wx、Wx、Wx、等[8],其中在我国育成品种中应用最多的是Wx、Wx和三个等位基因[9]。有无直链淀粉是区分非糯稻与糯稻的判断依据,这种差异是由基因的缺失突变引起的,非糯稻的基因型为,糯稻的基因型为。我国非糯水稻品种中主要应用的是Wx和Wx两种等位变异,对应高和中低直链淀粉含量[10]。以上3个等位基因调控直链淀粉合成的分子机理及其对稻米品质的影响较为明确[4-6, 8]。基因的等位变异是引起支链淀粉结构差异进而导致稻米糊化温度差异的主要原因[7, 11]。虽然基因在自然界中存在较多的单倍型,但根据其编码的酶是否有活性,主要分为两大类[12]:控制高糊化温度的ALK(又称a)和控制低糊化温度的ALK(又称a)。与基因相似,基因也存在籼粳稻之间的分化,其中ALK主要存在于籼稻中,ALK主要存在于粳稻中。
目前,虽然有不少学者围绕和等位基因的功能及其对品质的效应开展了大量研究,但鲜有研究分析二者不同等位基因组合对稻米品质的具体效应。有学者在已知基因型的水稻材料中测定糊化温度,认为稻米糊化温度与基因型不相关,推断糊化温度与位点无相关性[13-14],但该研究中没有区分相同等位基因背景下的等位变异对稻米糊化温度的影响。Saiyavit等[15]以11个泰国水稻品种为试验材料,发现糊化温度与直链淀粉含量显著正相关。苏文丽等[16]利用遗传背景相对一致的重组自交系BC1F8株系中的58个单株为材料,发现稻米直链淀粉含量与糊化温度呈极显著负相关。Xiang等[17]考查了17个水稻品种中的等位变异和的等位变异对稻米理化品质的效应,发现Wx背景下的低糊化温度ALK/ALK型的直链淀粉含量和糊化温度较低,胶稠度较软,品质较好;并且发现利用RVA曲线可以直观地区分直链淀粉含量和胶稠度对品质的影响,但不能区分糊化温度对品质的影响。以上这些研究明确了的等位变异和的等位变异及其组合对稻米品质的可能影响,但都是在背景不同的品种中开展的。为此,本研究聚焦基因和基因的主要等位变异(Wx/Wx/和ALK/ALK),通过创建近等基因系来研究这些等位变异组合对稻米品质的效应。近等基因系材料的创建和使用消除了遗传背景和生长环境对品质性状的影响,在相同的遗传背景和环境条件下研究和不同等位基因组合对稻米理化品质的影响,对于围绕稻米品质相关的重要基因开展优质稻米的分子设计育种有着重要的指导意义。
1 材料与方法
1.1 水稻近等基因系的构建与种植
本实验室在前期研究中已构建了在日本晴(Nipponbare, Nip)背景下分别含有、Wx和Wx不同等位基因三个近等基因系Nip-/ALK, Nip-Wx/ALK和Nip-Wx/ALK,它们的位点都是来自日本晴本身的ALK等位基因[8],和Wx的供体亲本分别为粳糯稻品种苏御糯和籼稻品种龙特甫(表1)。以上述三个近等基因系为受体亲本,进一步以高糊化温度的籼稻品种中恢9308(携带ALK等位基因)为供体亲本,通过杂交和多次回交并结合分子标记辅助选择,分别构建含有不同等位基因、相同ALK等位基因的另三个近等基因系Nip-/ALK, Nip-Wx/ALK和Nip-Wx/ALK(表1)。所用近等基因系都经过至少6代回交和多次自交选育获得。所用水稻材料在扬州大学农学院水稻试验田和海南陵水育种基地种植(加代繁殖),按常规的水肥条件进行栽培管理。用于品质测定的材料同季种植于扬州大学农学院水稻试验田。
1.2 DNA抽提与分子标记检测
水稻基因组DNA的抽提按Murray和Thompson[18]的方法进行。针对糯稻品种中基因第2外显子中前部23 bp的插入,设计分子标记ND以区分糯稻和非糯稻[19]。针对Wx和Wx两个等位基因在第一内含子处CT序列变异设计分子标记QRM190[20]。基于基因第8外显子4211 bp(A/G多态性),设计四引物4211以区分ALK和ALK。引物设计采用Primer 5.0软件,引物序列详见表2,PCR产物用3%琼脂糖凝胶电泳分离鉴定。
1.3 样品制备与稻米理化品质的测定
米粉的制备以及直链淀粉含量和胶稠度的测定参照农业部颁布的标准《米质测定方法》(NY147–1988)进行。米粉热力学特性的测定参照Zhang等[21]的方法进行,采用差示扫描量热仪DSC 200 F3(德国耐驰公司)配套的软件进行作图分析[22]。稻米的黏滞性采用3-D型淀粉黏滞性快速测定仪(Newport Scientific,澳大利亚)进行测定[23],并用配套软件进行分析。米粉粗蛋白含量的测定根据中华人民共和国国家标准[24],采用全量凯氏定氮法进行。
1.4 米饭食味值的测定
米饭食味值采用RCTA-11A型米饭食味计(日本佐竹)进行测定,具体步骤如下:称取整精米20.0 g,使用配套的淘米用具在自来水下清洗,清洗完毕后转移至盛有40 mL蒸馏水的铝制容器中,用电饭锅蒸煮30 min后室温冷却20 min,称取8.0 g米饭,装入金属环内,正反方向各压10 s,置于食味仪上测得热饭食味值。测过热饭食味值的样品于4℃下保存过夜后用相同的程序测得冷饭食味值。
1.5 数据分析
数据采用Microsoft Excel 2016 以及SPSS 19.0 软件进行统计分析;采用Adobe Illustrator制图;采用最小显著性差异(LSD)法进行差异显著性检验。
2 结果与分析
2.1 近等基因系中Wx和ALK基因型及植株表型的鉴定
用引物ND区分与Wx(或Wx),分子标记鉴定结果显示,Nip-背景下的PCR产物比Nip-Wx背景下的产物大23 bp(图1-A)。引物QRM190可以区分Wx与Wx,QRM190引物的分子标记鉴定结果显示,Nip-Wx背景下的PCR产物条带较大,Nip-Wx背景下的PCR产物条带较小(图1-B)。引物4211可以区分ALK(4211A)和ALK(4211G),4211(A/G)四引物的分子标记鉴定结果显示,Nip-ALK背景下的PCR产物条带为503 bp和197 bp,Nip-ALK背景下的PCR产物条带为503 bp和343 bp(图1-C)。6个近等基因系在田间生长期的表型无显著差异,株高、抽穗期、粒形和粒重等也无显著差异。
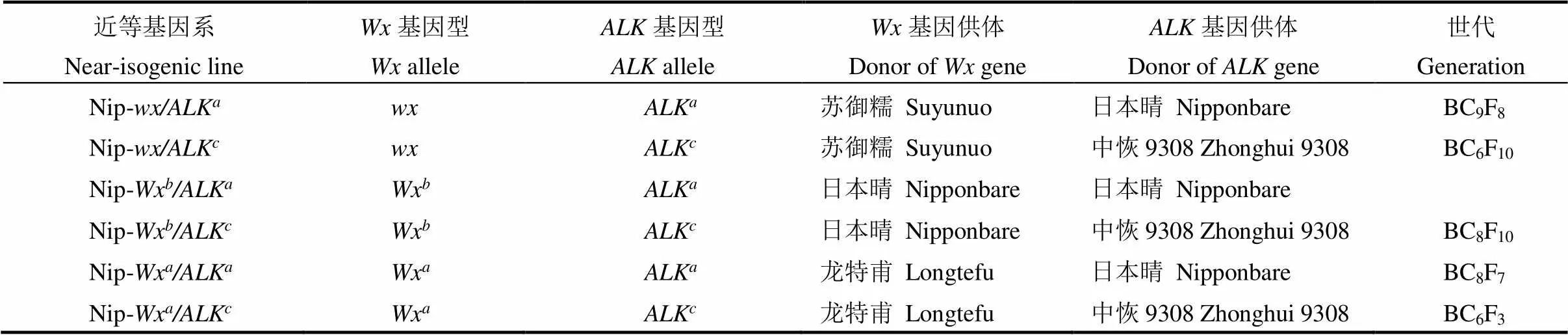
表1 近等基因系的基本信息
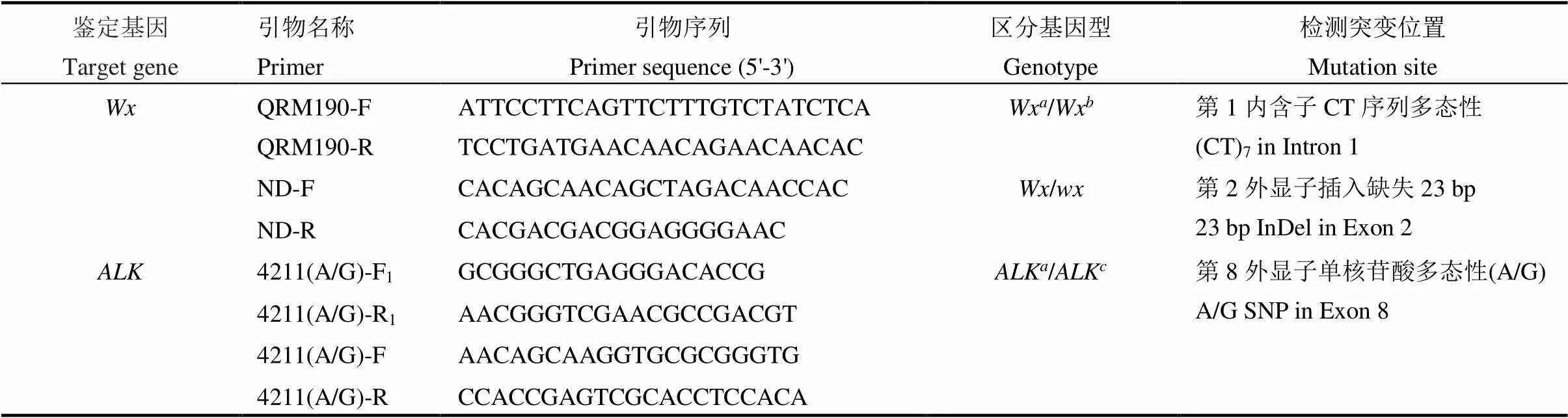
表2 用于鉴定水稻Wx和ALK不同基因型的分子标记

A-采用引物ND进行PCR鉴定。B-采用引物QRM190进行PCR鉴定。C-采用引物4211进行PCR鉴定。
Fig. 1. Identification ofandgenotypes in NILs by PCR.
2.2 ALK等位变异对稻米直链淀粉含量的影响
对成熟籽粒的品质进行比较发现,含有Wx、Wx和不同等位基因的三组近等基因系稻米直链淀粉含量存在极显著差异,表现为Wx(直链淀粉含量为28.1%~29.4%)>Wx(14.2%~14.8%)>(4.0%~4.1%)(图2-A)。而在相同的Wx、Wx或背景下,ALK和ALK近等基因系稻米之间的直链淀粉含量无显著差异,说明等位变异对稻米直链淀粉含量无显著影响。前人研究表明,具有相同基因型的一些水稻品种的直链淀粉含量依然存在较大的差异,说明稻米直链淀粉含量除了受主效基因控制外,还会受到其他基因的影响。此前,有研究表明调控糊化温度的基因与直链淀粉含量无显著相关性[7, 25-26]。
2.3 Wx等位变异对稻米糊化特性有显著影响
由表3和图3可见,ALK近等基因系稻米的起始糊化温度(T)、峰值糊化温度(T)和终止糊化温度(T)都极显著低于ALK,说明的等位变异是引起不同稻米间糊化温度差异的最主要原因。进一步分析发现,在相同ALK背景下,不同等位基因的近等基因系间糊化温度有一些差异,尤其是Wx基因型稻米的糊化温度显著低于Wx或型;在相同ALK背景下的三个等基因系中,也出现类似的情况。具体表现为Nip-Wx/ALK和Nip-/ALK,Nip-Wx/ALK和Nip-/ALK之间的峰值糊化温度几乎无差异;但Nip-Wx/ALK稻米的峰值糊化温度极显著低于Nip-Wx和Nip-背景下的ALK近等基因系;同样地,Nip-Wx/ALK稻米的T极显著低于Nip-Wx和Nip-背景下的ALK近等基因系。前人利用不同的水稻材料进行研究,认为糊化温度与直链淀粉含量呈显著负相关[25, 27]。在本研究中Wx背景下近等基因系稻米的糊化温度低于Wx和背景下相应的近等基因系,说明稻米糊化温度不仅受主效基因控制,也会受到基因的影响。
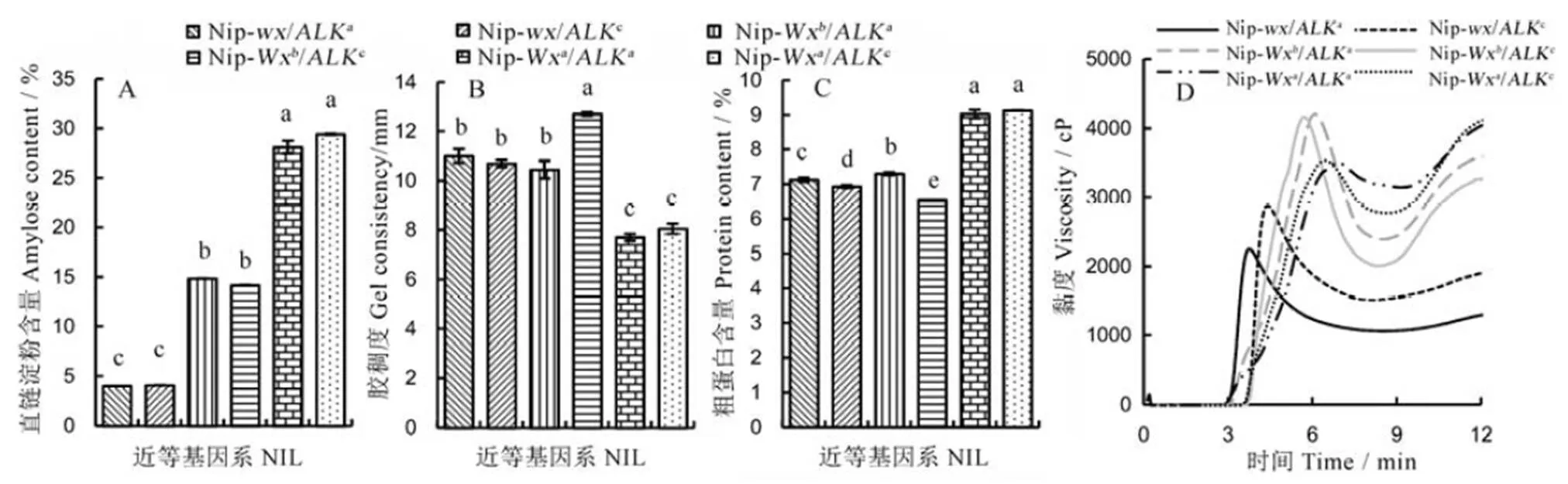
对近等基因系的品质性状进行方差分析,并用新复极差法进行多重比较。相同小写字母表示近等基因系间无显著差异(n=3)。
Fig. 2. Effect ofandallelic diversity on rice eating and cooking qualities.

图3 Wx与ALK等位基因组合对米粉差示扫描量热(DSC)图谱的影响
Fig. 3. Effect ofandallelic diversity on differential scanning calorimetry(DSC) curve of rice flour.
2.4 Wx和ALK不同等位变异组合对其他品质性状的影响
本研究中日本晴背景下6个近等基因系稻米的胶稠度均超过50 mm,都属于优质米范畴。其中,基因的等位变异对稻米胶稠度的影响较大,具体表现为高直链淀粉含量的Nip-Wx背景下稻米胶稠度数值极显著低于Nip-Wx和Nip-型(图2-B);而中低直链淀粉含量的Nip-Wx与糯稻Nip-的稻米之间的胶稠度几乎无差异。基因的等位变异对稻米的胶稠度也有一定的影响,在糯稻Nip-和高直链淀粉含量的Nip-Wx背景下,ALK型稻米与ALK型稻米之间的胶稠度没有显著差异;但在中低直链淀粉含量的Nip-Wx背景下Nip-Wx/ALK型稻米的胶稠度显著高于Nip-Wx/ALK型。
从图2-D和表4可以看出,基因等位变异主要影响稻米的峰值黏度和峰值时间。Nip-Wx的峰值黏度最高,Nip-的峰值黏度最低;Nip-的峰值时间最早,Nip-Wx的峰值时间最迟。相同背景下等位变异对米粉黏度影响不大。前人对稻米淀粉RVA谱特征值与其他食味品质指标的关系进行研究,发现稻米消减值与表观直链淀粉含量极显著正相关[28]。在本研究中Nip-背景下的两个近等基因系稻米的RVA曲线整体较低,其峰值黏度、热浆黏度和冷胶黏度都较低,Nip-Wx背景下稻米的峰值黏度最大,加上这两个近等基因系稻米的冷胶黏度极显著低于Nip-Wx的近等基因系,因此Nip-Wx稻米的崩解值大于Nip-Wx型稻米。等位变异主要影响稻米的起浆温度,表现为ALK背景的近等基因系稻米的糊化温度高于ALK背景下相应的近等基因系,相同的ALK或ALK背景下,Wx近等基因系米粉的起浆温度低于Wx和的近等基因系,这与糊化温度的规律一致。稻米崩解值受到基因和基因的共同影响,表现为ALK近等基因系稻米的崩解值要高于ALK型。在相同的背景下,基因型对稻米崩解值的影响表现为Wx>>Wx。结合消减值来看,Nip-Wx近等基因系稻米的消减值较高,因而食味值较差;Nip-Wx背景下高糊化温度的ALK型稻米的崩解值最大,消减值较小。
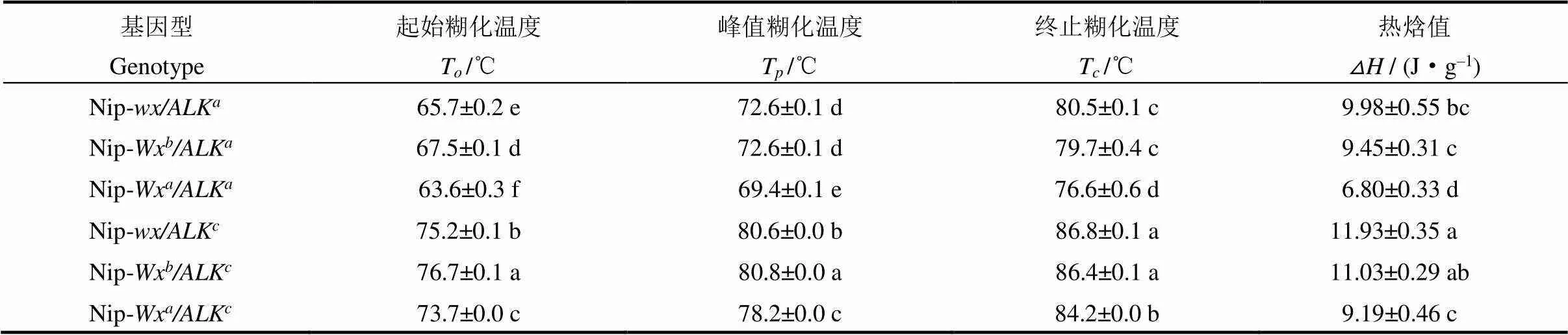
表3 Wx与ALK等位基因组合对米粉差示扫描量热(DSC)特征值的影响
表中数据均采用平均值±标准差表示(=2)。同列相同字母表示基因型间无差异。下表同。
The data in the table are represented by mean ±SD(=2). In a column, the same letter indicate no difference among genotypes. The same as below.

表4 Wx与ALK等位基因组合对米粉黏滞性的影响
2.5 Wx和ALK不同等位变异组合对稻米食味值的影响
从表5中的热饭食味值可以看出,Nip-Wx背景下的近等基因系稻米的外观、黏度和平衡度均极显著低于Nip-Wx和Nip-背景下相应的近等基因。此外,Nip-Wx近等基因系稻米的硬度极显著高于Nip-Wx和Nip-的近等基因系,Nip-Wx背景下近等基因系热饭食味值的综合评分极显著低于Nip-Wx和Nip-背景下的近等基因系,具有中低直链淀粉含量的Wx型与糯稻型稻米的热饭食味值无显著差异。热饭食味值在相同背景下的两个近等基因系之间无差异。就冷饭食味值而言,Nip-Wx背景下的两个近等基因系的综合评分依然最低,在热饭食味值评分相对一致的Nip-Wx和Nip-的4个近等基因系中,ALK的近等基因系冷饭的食味值降幅要大于ALK的近等基因系。其中,Nip-/ALK近等基因系稻米的外观、黏度和平衡度极显著下降,导致其综合评分极显著下降,但在中低直链淀粉含量的Nip-Wx背景下,高糊化温度ALK近等基因系的冷饭食味值虽有下降,但评分显著高于Nip-Wx和Nip-背景下的ALK近等基因系。
3 讨论
3.1 直链淀粉主要受Wx基因影响,不受ALK基因影响
前人在对稻米品质的相关研究中,以直链淀粉含量为切入点开展的研究较多。曾瑞珍等[29]在243个水稻品系中检测到15种复等位基因。有研究者认为栽培稻中的等位基因均由祖先基因Wx经过自然进化和人工选择而来[8]。在所有的等位变异中,以Wx、Wx和三个等位基因占比较大。其中籼稻中以Wx为主,携带该等位基因的稻米属于高直链淀粉类型,直链淀粉含量可以达到25%以上;粳稻中以Wx为主,直链淀粉含量属于中等至较低水平(15%~18%),在糯稻品种中等位基因是,由于第2外显子上存在23 bp碱基的插入,导致基因的转录提前终止,不能翻译完整的GBSSⅠ来催化合成直链淀粉,因此糯稻的直链淀粉含量极低。本研究结果表明稻米直链淀粉含量主要受基因影响,不同基因型稻米的直链淀粉含量含量高低排序为Wx>Wx>。并且在本研究中等位变异对稻米直链淀粉含量无影响,表现为在相同的Nip-Wx、Nip-Wx或Nip-背景下相应的ALK和ALK近等基因系稻米之间的直链淀粉含量无显著差异。这是由于基因主要通过改变支链淀粉结构来影响糊化温度[30],对直链淀粉含量没有影响。但有研究表明,直链淀粉含量相似的水稻品种之间的品质仍然存在较大的差异[31],推测参与支链淀粉合成的其他基因等可能会对稻米品质产生一定的影响[32-33]。

表5 不同Wx和ALK等位基因组合对精米食味值的影响
3.2 糊化温度主要受ALK基因影响,同时受Wx基因调控
目前研究表明稻米糊化温度受主效基因调控。通过对不同糊化温度水稻品种的基因序列进行比较,发现基因的外显子区域存在和糊化温度高度相关的SNP。本研究选取的基因C末端两个SNP的等位变异型,分别为控制高糊化温度的ALK(G-GC)和控制低糊化温度的ALK(A-GC)。在相同的背景下,ALK型稻米的糊化温度极显著高于ALK型,但在不同的背景下,ALK近等基因系和ALK近等基因系之间糊化温度的差值存在差异。在高直链淀粉含量的Nip-Wx背景下,近等基因系之间峰值糊化温度的差值显著高于Nip-Wx和Nip-背景下近等基因系之间峰值糊化温度的差值。在相同的背景下,和Wx型稻米的糊化温度差异不大,但Wx型稻米的糊化温度极显著低于型和Wx型。以上表明,在支链淀粉含量差异较大的材料中,稻米糊化温度与直链淀粉含量呈负相关,这可能是由于高直链淀粉含量材料中颗粒结合淀粉合成酶的高活性以及它和可溶性淀粉合成酶对底物的竞争性导致的。和基因分别影响直链淀粉和支链淀粉的结构,对短直链和长直链的数量有明显的影响[8],而糊化温度可能与短直链和长直链的数量有关;基因的等位差异主要导致单层淀粉中链和短链的相对量变化,从而导致糊化温度的差异[34]。
3.3 胶稠度主要受Wx基因影响,也受ALK调控
不同水稻品种间的胶稠度差异较大,其遗传基础比较复杂,但由于稻米胶稠度与直链淀粉含量负相关,所以胶稠度与基因有关联[35-36]。本研究中Wx型稻米的胶稠度极显著低于Wx和型。这主要是由Wx型稻米较高的直链淀粉含量导致的,直链淀粉含量过高的品种米饭较硬,导致米粒的延伸性不好,胶稠度较低[37]。在Nip-Wx背景下高糊化温度的ALK近等基因系的胶稠度较高,说明基因在中低直链淀粉含量的Nip-Wx背景下对稻米胶稠度的变异有一定的修饰作用,这与前人的研究一致[7],主要表现为高糊化温度的ALK近等基因系稻米的胶稠度要高于低糊化温度的ALK型。
3.4 稻米RVA特性受Wx和ALK等位基因组合影响较大
直链淀粉含量差异对RVA曲线影响较大,在高直链淀粉含量的近等基因系中,稻米胶稠度和崩解值较小,热浆黏度和冷胶黏度较高,峰值时间相对较长。RVA二级数据中的崩解值、消减值和回复值与稻米食用品质的优劣显著相关。通常,崩解值大于100 RVU而消减值小于25 RVU的品种食味较好,相反,食味差的品种稻米崩解值小于36 RVU,而消减值大于80 RVU[38]。从RVA谱分析可以推测,Nip-Wx背景下近等基因系稻米的食味较差,主要是由其较高的消减值导致的,而Nip-Wx背景下的近等基因系由于较高的崩解值和较低的消减值,因而稻米的食味较好。这可能与该背景下近等基因系稻米较高的胶稠度有关。等位基因主要影响起浆温度,稻米崩解值和消减值在相同背景下的近等基因系之间差异不大。
4 结论
本研究进一步明确了不同和主要等位变异组合对稻米品质形成的效应,得到以下结论:稻米直链淀粉含量和胶稠度主要由基因控制,稻米糊化温度和淀粉黏滞性由和基因共同调控。就和基因不同等位变异组合对品质的具体影响来看,Nip-Wx/ALK和Nip-Wx/ALK型稻米具有较高的直链淀粉含量和崩解值,以及较低的胶稠度,因而食味值较差,但其稻米的蛋白质含量较高,具有较好的营养价值;Nip-/ALK和Nip-Wx/ALK型稻米具有适中的直链淀粉含量、胶稠度和蛋白质含量以及较低的糊化温度,热饭和冷饭的食味值较好;Nip-Wx/ALK型稻米虽然糊化温度较高,但其适中的直链淀粉含量和较高的胶稠度导致其冷饭食味值降幅较小;Nip-/ALK稻米的热饭食味值与Nip-/ALK、Nip-Wx/ALK和Nip-Wx/ALK无差异,但因其较高的糊化温度,其冷饭食味值显著下降。以上研究明确了这两个品质相关主效基因的不同组合对稻米理化品质的效应,对优质稻米的分子设计育种具有重要的指导意义。
[1] Vandeputte G E, Delcour J A. From sucrose to starch granule to starch physical behaviour: A focus on rice starch[J]., 2004, 58(3): 245-266.
[2] Singh N, Kaur L, Sandhu K S, Kaur J, Nishinari K. Relationships between physicochemical, morphological, thermal, rheological properties of rice starches[J]., 2006, 20(4): 532-542.
[3] 包劲松. 稻米淀粉品质遗传与改良研究进展[J]. 分子植物育种, 2007, 5(6): 1-20.
Bao J S. Progress in sutdies on inheritance and improvement of rice starch qualiy[J]., 2007, 5(6): 1-20. (in Chinese with English abstract)
[4] Wang Z Y, Zheng F Q, Shen G Z, Gao J P, Snustad D P, Li M G, Zhang J L, Hong M M. The amylose content in rice endosperm is related to the post-transcriptional regulation of thegene[J]., 1995, 7(4): 613-622.
[5] Tian Z X, Qian Q, Liu Q Q, Yan M X, Liu X F, Yan C J, Liu G F, Gao Z Y, Tang S Z, Zeng D L, Wang Y H, Yu J M, Gu M H, Li J Y. Allelic diversities in rice starch biosynthesis lead to a diverse array of rice eating and cooking qualities[J]., 2009, 106(51): 21760-21765.
[6] Cai X L, Wang Z Y, Xing Y Y, Zhang J L, Hong M M. Aberrant splicing of intron 1 leads to the heterogeneous 5′ UTR and decreased expression ofgene in rice cultivars of intermediate amylose content[J]., 1998, 14(4): 459-465.
[7] Gao Z Y, Zeng D L, Cheng F M, Tian Z X, Guo L B, Su Y, Yan M X, Jiang H, Dong G J, Huang Y C, Han B, Li J Y, Qian Q., the key gene for gelatinization temperature, is a modifier gene for gel consistency in rice[J]., 2011, 53(9): 756-765.
[8] Zhang C Q, Zhu J H, Chen S J, Fan X L, Lu Y, Wang M, Yu H X, Yi C D, Tang S Z, Gu M H, Liu Q Q.Wx, the ancestral allele of ricegene[J]., 2019, 12(8): 1157-1166.
[9] 朱霁晖, 张昌泉, 顾铭洪, 刘巧泉. 栽培稻中基因的等位变异及育种利用研究进展[J]. 中国水稻科学, 2015, 29(4): 431-438.
Zhu J H, Zhang C Q, Gu M H, Liu Q Q. Progress in the allelic variation ofgene and its application in rice[J]., 2015, 29(4): 431-438. (in Chinese with English abstract)
[10] Sano Y. Differential regulation ofgene expression in rice endosperm[J]., 1984, 68(5): 467-473.
[11] 范名宇, 王晓菁, 王旭虹, 唐亮, 徐铨, 徐正进. 稻米支链淀粉结构的研究进展[J]. 中国水稻科学, 2017, 31(2): 124-132.
Fan M Y, Wang X J, Wang X H, Tang L, Xu Q, Xu Z J. Research advances of amylopectin structure in rice[J]., 2017, 31(2): 124-132. (in Chinese with English abstract)
[12] Jeon J S, Ryoo N, Hahn T R, Walia H, Nakamura Y. Starch biosynthesis in cereal endosperm[J]., 2010, 48(6): 383-392.
[13] 孙业盈, 吕彦, 董春林, 王平荣, 黄晓群, 邓晓建. 水稻基因与稻米AC、GC和GT的遗传关系[J]. 作物学报, 2005, 31(5): 535-539.
Sun Y Y, Lv Y, Dong C L, Wang P R, Hang X Q, Deng X J. Genetic relationship amonggene, AC, GC and GT of rice[J]., 2005, 31(5): 535-539. (in Chinese with English abstract)
[14] 刘玉花, 栾丽, 龙文波, 王兴, 孔繁伦, 何涛, 涂升斌. 同源四倍体和二倍体水稻基因与淀粉品质的遗传关系[J]. 中国水稻科学, 2007, 21(2): 143-149.
Liu Y H, Luan L, Long W B, Wang X, Kong F L, He T, Tu S B. Genetic relationships betweengene and starch quality in autotereaploid and diploid rice[J]., 2007, 21(2): 143-149. (in Chinese with English abstract)
[15] Varavinit S, Shobsngob S, Varanyanond W, Chinachoti P, Naivikul O. Effect of amylose content on gelatinization, retrogradation and pasting properties of flours from different cultivars of Thai rice[J]., 2003, 55(9): 410-415.
[16] 苏文丽, 向珣朝, 徐艳芳, 龙小林, 康翠芳, 许顺菊. 水稻基因型对成糊温度校准的影响及糊化温度与直链淀粉含量的关系分析[J]. 植物遗传资源学报, 2014, 15(2): 348-353.
Su W L, Xiamg X C, Xu Y F, Long X L, Kang C F, Xu S J. The influence ofgenotype on the calibration of pasting temperature of rice and the relationship between gelatinization temperature and amylose content[J]., 2014, 15(2): 348-353. (in Chinese with English abstract)
[17] Xiang X C, Kang C F, Xu S J, Yang B W. Combined effects ofandhaplotypes on rice starch physicochemical properties[J]., 2016, 97(4): 1229-1234.
[18] Murray M G, Thompson W F. Rapid isolation of high molecular weight plant DNA[J]., 1980, 8(19): 4321-4325.
[19] Fan X L, Li Y Q, Lu Y, Zhang C Q, Li E P, Li Q F, Tao K Y, Yu W W, Wang J D, Chen Z Z, Liu Q Q. The interaction between amylose and amylopectin synthesis in rice endosperm grown at high temperature[J/OL]., 2019, 301: 125258.
[20] 张昌泉, 潘立旭, 周兴忠, 李钱峰, 刘巧泉. 不同等位组合对杂交稻天优3611品质的影响[EB/OL]. 北京:中国科技论文在线[2017-04-28]. http://www. paper. edu.cn/releasepaper/content/201704-764.
Zhang C Q, Pan L X, Zhou X Z, Li Q F, Liu Q Q. The combined effects of differentallele on grain quality of hybrid rice Tianyou 3611[EB/OL]. Beijing: Chinese Science Paper Online [2017-04-28]. (http://www. paper. edu.cn/releasepaper/content/201704-764). (in Chinese with English abstract)
[21] Zhang C Q, Zhou L H, Zhu Z B, Lu H W, Zhou X Z, Qian Y T, Li Q F, Lu Y, Gu M H, Liu Q Q. Characterization of grain quality and starch fine structure of two japonica rice () cultivars with good sensory properties at different temperatures during the filling stage[J]., 2016, 64(20): 4048-4057.
[22] Zhang C Q, Zhu L J, Shao K, Gu M H, Liu Q Q. Toward underlying reasons for rice starches having low viscosity and high amylose: Physiochemical and structural characteristics[J]., 2013, 93(7): 1543-1551.
[23] Zhu L J, Liu Q Q, Sang Y, Gu M H, Shi Y C. Underlying reasons for waxy rice flours having different pasting properties[J]., 2010, 120(1): 94-100.
[24] 戴宏林, 吴小骏. 用凯氏定氮法测定植物干样品中的氮含量[J]. 扬州大学学报: 农业与生命科学版, 1995, 16(3): 70.
Dai H L, Wu X J. Determination of nitrogen in dry plant samples by Kjeldahl nitrogen determination[J]., 16(3): 70. (in Chinesewith English abstract)
[25] 马玉清, 李仕贵, 王玉平, 黎汉云, 高克铭. 籼型杂交水稻稻米品质的初步研究[J]. 西南农业学报, 2002, 15(1): 28-31.
Ma Y Q, Li S G, Wang Y P, Li H Y, Gao K M. Preliminary study on quality traits of indica hybrid rice[J]., 2002, 15(1): 28-31. (in Chinese with English abstract)
[26] Bao J S, Sun M, Zhu L H, Corke H. Analysis of quantitative trait loci for some starch properties of rice (L.): Thermal properties, gel texture and swelling volume[J]., 2004, 39(3): 379-385.
[27] 夏明元, 李进波, 张建华, 万丙良, 戚华雄. 利用分子标记辅助选择技术选育具有中等直链淀粉含量的早稻品种[J]. 华中农业大学学报, 2004, 23(2): 183-186.
Xia M Y, Li J B, Zhang J H, Wan B L, Qi H X. Breeding of earlyrice with intermediate amylose content by molecular marker-aided selection[J]., 2004, 23(2): 183-186. (in Chinese with English abstract)
[28] 舒庆尧, 吴殿星, 夏英武, 高明尉, McClung A. 稻米淀粉RVA谱特征与食用品质的关系[J]. 中国农业科学, 1998, 31(3): 25-29.
Shu Q Y, Wu D X, Xia Y W, Gao M W, McClung A. Relationship between RVA character and eating quality inL[J]., 1998, 31(3): 25-29. (in Chinese with English abstract)
[29] 曾瑞珍, 张泽民, 何风华, 席章营, Talukdar A, 施军琼, 秦利军, 黄朝锋, 张桂权. 水稻复等位基因的鉴定及单片段代换系的建立[J]. 中国水稻科学, 2005, 19(6): 495-500.
Zeng R Z, Zhang Z M, He F H, Xi Z Y, Akshay T, Shi J Q, Qin L J, Huang C F, Zhang G Q. Identification of multiple alleles at thelocus and development of single segment substitution lines in rice[J]., 2005, 19(6): 495-500. (in Chinese with English abstract)
[30] 陈专专, 李先锋, 仲敏, 葛家奇, 范晓磊, 张昌泉, 刘巧泉. 籼稻背景下抑制不同等位基因表达对稻米品质的影响[J]. 中国水稻科学, 2019, 33(6): 513-522.
Chen Z Z, Li X F, Zhong M, Ge J Q, Fan X L, Zhang C Q, Liu Q Q. Effects on grain quality by down-regulation of the expression of differentallele inrice (L.)[J]., 2019, 33(6): 513-522. (in Chinese with English abstract)
[31] 李欣, 顾铭洪, 潘学彪. 常见水稻品种稻米品质的研究[J]. 扬州大学学报:农业与生命科学版, 1987, 8(1): 1-8.
Li X, Gu M H, Pan X B. A study on grain quality of rice for commercial varieties[J]., 1987, 8(1): 1-8. (in Chinese with English abstract)
[32] Han X Z, Hamaker B R. Amylopectin fine structure and rice starch paste breakdown[J]., 2001, 34(3): 279-284.
[33] Ong M H, Blanshard J M V. Texture determinants of cooked, parboiled rice. II: Physicochemical properties and leaching behaviour of rice[J]., 1995, 21(3): 261-269.
[34] Wang K, Hasjim J, Wu A C, Li E P, Henry R J, Gilbert R G. Roles of GBSSI and SSIIa in determining amylose fine structure[J]., 2015, 127: 264-274.
[35] 万映秀, 邓其明, 王世全, 刘明伟, 周华强, 李平. 水稻基因的遗传多态性及其与主要米质指标的相关性分析[J]. 中国水稻科学, 2006, 20(6): 603-609.
Wan Y X, Deng Q M, Wang S Q, Liu M W, Zhou H Q, Li P. Genetic polymorphism ofgene and its correlation with major grain quality traits in rice[J]., 2006, 20(6): 603-609. (in Chinese with English abstract)
[36] Bao J S, Corke H, He P, Zhu L H. Analysis of quantitative trait loci for starch properties of rice based on an RIL polulation[J]., 2003, 45(8): 986-994.
[37] 朱昌兰, 江玲, 张文伟, 王春明, 翟虎渠, 万建民. 稻米直链淀粉含量和胶稠度对高温耐性的QTL分析[J]. 中国水稻科学, 2006, 20(3): 248-252.
Zhu C L, Jiang L, Zhang W W, Wang C M, Zhai H Q, Wan J M. Identifying QTLs for thermo-tolerance of amylose content and gel consistency in rice[J]., 2006, 20(3): 248-252. (in Chinese with English abstract)
[38] 盛文涛, 吴俊, 姚栋萍, 庄文, 邓启云. 稻米食味品质的影响因素及中国籼稻食味品质改良存在的问题与策略[J]. 杂交水稻, 2014, 29(3): 1-5.
Sheng W T, Wu J, Yao D P, Zhuang W, Deng Q Y. Influencing factors of rice eating quality and existing problems and strategies of genetic improvement on eating quality ofrice in China[J]., 2014, 29(3): 1-5. (in Chinese with English abstract)
Effects of Different Combinations ofandMain Alleles on Rice Grain Quality
CHEN Zhuanzhuan1, YANG Yong1, FENG Linhao1, SUN Ye1, ZHANG Changquan1, 2, FAN Xiaolei1, 2,LI Qianfeng1, 2, LIU Qiaoquan1, 2, *
(1Key Laboratory of Plant Functional Genomics of Ministry of Education / Jiangsu Key Laboratory of Crop Genomics and Molecular Breeding, College of Agriculture, Yangzhou University, Yangzhou 225009, China;2Jiangsu Key Laboratory of Crop Genetics and Physiology / Jiangsu Collaborative Innovation Center for Modern Industrial Technology of Grain Crops, Yangzhou University, Yangzhou 225009, China;*Corresponding author, E-mail: qqliu@yzu.edu.cn)
【】Rice eating and cooking qualities are mainly controlled by two key genes,and. There are multiple alleles of both genes among rice cultivars. In this study, our aim is to clarify the effects of the main allelic variation combinations ofandgenes on rice grain quality.【】Three main alleles of(Wx,Wxand) and two of(ALKandALK) were combined within a series of near-isogenic lines (NILs) in theNipponbare (Nip) background. The effects of different allelic combinations on grain quality were determined by measuring the physical and chemical characteristics of NILs.【】Gelatinization temperature (GT) was regulated by the major geneand the minor gene. Under the sameallele background, there was no significant difference in GT between rice linescarryingandWxalleles, but the GT of Nip-Wxrice lines was significantly lower than that of Nip-Wxand Nip-. The starch viscosity was greatly affected by the combination ofandalleles. The breakdown value of Nip-Wxrice line was lower and the setback value was higher (lower tasting value) than those of Nip-Wxand Nip-. Under the sameallele background,allelic variation mainly affected the pasting temperature of RVA curve, but had little effect on breakdown value and setback value. Nip-Wx/ALKand Nip-Wx/ALKrice grains had poor tasting value due to the high AC and breakdown value and low GC. But the protein content in above two NILs was higher than other rice lines. Nip-/ALKand Nip-Wx/ALkrice grains had better tasting value of hot cooked and cooled cooked rice due to the moderate AC, GC and protein content, as well as lower GT. Although the GT of Nip-Wx/ALKgrains was higher, the moderate AC and high GC of this line resulted in a slight decrease in the tasting value of cooled cooked rice compared with hot cooked rice. The tasting value of hot cooked rice of Nip-/ALKrice remained the same as those of Nip-/ALK, Nip-Wx/ALKand Nip-Wx/ALK, but the tasting value of cooled rice decreased dramatically because of its high GT.【】Amylose content (AC) and gel consistency (GC) were mainly controlled by Wx gene; while ALK gene had no significant effect on AC, but a slight effect on GC.
waxy gene; alkali digestion gene; allelic variation; near isogenic lines; grain quality
Q755; S511.032
A
1001-7216(2020)03-0228-09
10.16819/j.1001-7216.2020.9140
2019-12-29;
2020-01-13。
国家自然科学基金资助项目(31825019, U19A2032);国家重点研发计划资助项目(2016YFD0100501);江苏省科技计划资助项目[BE2018357,CX(18)1001 和19KJA560006];扬州大学农学院本科生创新训练计划资助项目;江苏省研究生科研与实践创新计划资助项目(XKYCX17_050)。
