湿地松雄性不育和可育系小孢子叶球形态和细胞发育动态变化
2020-05-26周洋杨章旗黄永利
周洋 杨章旗 黄永利

摘 要: 为了解湿地松‘松泰小孢子叶球在发育过程形态是否有差异变化,明确其败育过程、败育方式及影响因素,为湿地松雄性不育品种利用和后期开展相关研究提供科学依据。该研究以‘松泰 s10败育系和s9可育系为材料,观察小孢子叶球形态发育变化,并对其小孢子叶球进行石蜡切片,在光学显微镜下观察小孢子发育过程。结果表明:s10败育系和s9可育系在小孢子母细胞减速分裂前无明显差异,小孢子叶球生长趋势也一致;四分體时期,s10小孢子细胞发育异常,小孢子叶球形态发育也出现异常,二者异常发育具有同步性;可育系从四分体到单核小孢子发育阶段的时间为5 d左右,而败育系持续发育长达20 d左右,持续时间为可育系的4倍。在此期间出现小孢子绒毡层细胞发育异常、降解缓慢,小孢子囊壁组织排列紊乱、降解延迟等现象,s10形成异常二核花粉,且无花粉散出。因此,推论s10小孢子败育的原因主要是小孢子囊壁细胞发育异常,其小孢子叶球形态异常,相对应的绒毡层在四分体时期发育异常,不能适时地分泌胼胝质酶来降解围绕着四分体的胼胝质壁,也不能适时地合成输送花粉形成所需能量物质,同时囊壁细胞出现降解延迟和层积,这一系列的异常变化导致不能形成正常四分体,从而使花粉败育。
关键词: 湿地松, 小孢子叶球, 雄性不育, 细胞发育, 形态变化
中图分类号: Q944.46 文献标识码: A
文章编号: 1000-3142(2020)04-0482-10
Abstract: In order to understand whether there are differences in morphology of microstrobilus of Pinus elliottii ‘Songtai during development, to clarify its abortion process, abortion mode and influencing factors, and to provide scientific basis for utilization and later research of male sterile varieties of Pinus elliottii, the morphological changes of microstrobilus were observed by using ‘Songtai s10 abortive line and s9 fertile line as materials, and the microstrobilus was sliced in paraffin and the development of microspore was observed under optical microscope. The results were as follows: There was no significant differences between s10 abortion line and s9 fertile line before meiotic phase of microsporocy deceleration and division, and the growth trend of microstrobilus was the same. In the period of tetrad, s10 microspore cell development and microstrobilus morphological development were abnormal. The time from tetrad to monocyte microspore development of fertile lines is about 5 d. However, the abortion line continued to develop for about 20 d, and the duration was four times that of the fertile line. In the meantime, the development of tapetum cells was abnormal, the degradation was slow, the tissue arrangement of microsporangium wall was disordered, and the degradation was delayed. Finally, s10 forms abnormal dinuclear pollen and no pollen dispersion. So it can be reasoned that the abnormal development of microsporangium wall cells, and the abnormal morphology of microstrobilus.The tapetum develops abnormally in the tetrad period, and the callose enzyme can not be secreted at appropriate time to degrade the callosum wall surrounding the tetrad, nor can it synthesize and transport the energy substances needed for pollen formation in a timely manner. At the same time, the cystic wall cells showed delayed degradation and lamination. This series of abnormal changes leaded to the normal tetrad could not be formed, which made the pollen abortion.
Key words: Pinus elliottii, microstrobilus, male sterile, cell development, morphological changes
湿地松(Pinus elliottii)原产于美国东南部温湿多雨的低海拔地区,海拔600 m以上,常绿大乔木(郑万钧等,1978;朱志淞,1993)。我国最早在20世纪30年代开始引种湿地松,并于1964年在广东省台山县建立了第一个湿地松初级种子园(唐国强,2015)。从20世纪70年代开始大面积推广湿地松以来,从海南岛屯昌县(110°26′ E、19°22′ N,年平均温度23.16 ℃)到山东半岛的青岛(120°25′ E、36°09′ N,年均温11.9 ℃),再到烟台(122° E、37°25′ N,年均温12 ℃),西至云南均有湿地松分布(潘志刚和游应天,1994)。目前湿地松在我国的人工林种植面积已超过300万公顷,成为了人工乔木林十大主要优势树种之一(张帅楠,2017)。
雄性不育(male sterility, MS)在高等植物中普遍存在,是指植物体不能产生正常有功能的花粉、花药或雄配子的遗传现象(范彦君等,2016)。“植物雄性不育”的概念最早在1876年由Coleman提出,迄今为止己在43个科、162个属、320个种中发现雄性不育现象(Kaul, 1988; Wise & Pring, 2002)。当前雄性不育的研究主要以经济农作物为主,研究也较为全面和深入,而林木由于育种时间长、不育材料发现少、生长周期长以及后期生长观测较难等原因,致使其研究较为复杂和困难,可查及的文献相对较少,研究深度也较浅,而松科类等裸子植物的雄性不育研究报道更为少见。‘松泰来源于美国佛罗里达州湿地松种源,于20世纪80年代,全国国外松协作组开展了湿地松引种种源试验,利用广西点种源试验的结果,1992年—1993年在南宁市林科所营建湿地松改良1代无性系种子园。团队经过连续5年以上的物候观测,‘松泰均无花粉散出,表现出雄性败育特征(唐国强,2015)。本次研究采用南宁市林科所湿地松改良1代种子园内‘松泰s10败育系和可育系s9为实验材料,通过对其小孢子叶球的形态发育过程、小孢子及其囊壁细胞结构发育过程进行观察,跟踪和了解‘松泰小孢子叶球的败育过程、败育方式及败育时间等,以期为湿地松雄性不育的败育机制及今后湿地松败育相关研究提供基础。同时,丰富林木雄性不育材料的不育机制理论,对松树杂交育种及其杂种优势利用提供依据。
1 材料与方法
1.1 材料
材料均采自广西南宁市林业科学研究所湿地松改良1代25年生种子园内‘松泰s10败育系和s9可育系植株。种子园位于108°00′ E、23°10′ N,属南亚热带南缘季风气候区,年均气温21.5 ℃,年均降雨量1 246 mm,年均蒸发量1 613.8 mm,全年无霜期358 d,干湿季节变化明显,平均相对湿度约79%,林地建在石灰岩峰林峰丛间宽阔的缓丘台地带,海拔高约120 m,土壤为赤红壤,pH为5.5~6.5,土层深厚。
1.2 研究方法
在湿地松种子园内分别确定3株‘松泰s10败育系和s9可育系作为取样植株,于2018年12月13日(小孢子叶球开始大量出现)开始取样,直到败育系小孢子叶球脱落(2月13号)和正常系小孢子叶球散粉完毕时(2月中旬)停止取样。2019年1月23号前,间隔6~7 d采一次;1月23号到2月7号,间隔3~4 d采一次;取样量每株树不少于15个小孢子叶球,并用FAA固定液固定于50 mL离心管内,贴上标签保存于4 ℃的冰箱内备用。
1.2.1 小孢子叶球形态观测 按照取样时间的不同选择具有代表性的小孢子叶球,观察记录其外部形态发育变化;每次采样随机选择30个小孢子叶球测量其长度与宽度,并记录分析其动态变化趋势。
1.2.2 小孢子及囊壁组织发育观测 采用常规石蜡制片方法,通过爱氏苏木精整体染色、脱水、透明、浸蜡、包埋、切片、封片等步骤(叶宝兴等,2011),对小孢子叶进行横切(切片厚度为5 μm),中性树胶封片,选取最具有代表性且清晰的样片,利用电动光学显微镜(型号:Axio Imagr Z2m)观测不同时期小孢子内部形态的发育变化并做好相关记录。
2 结果与分析
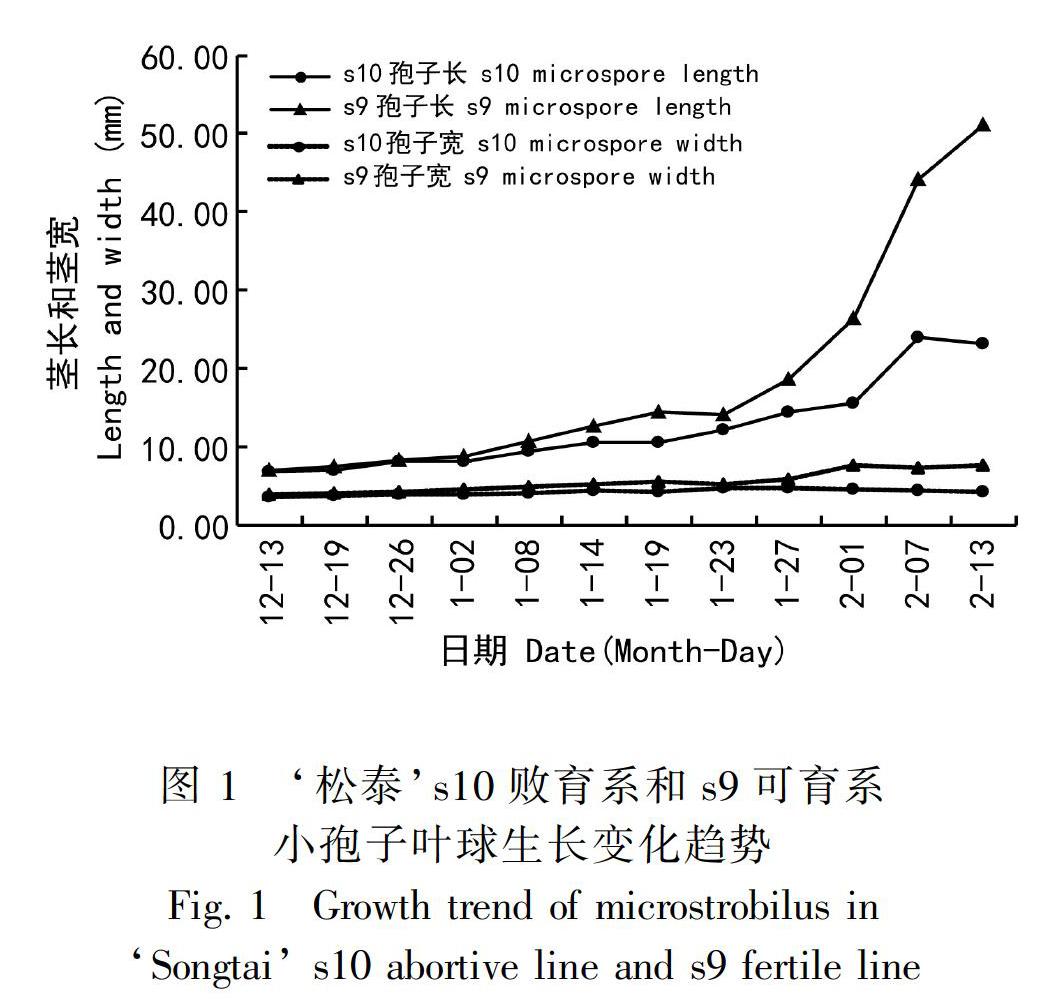
2.1‘松泰s10败育系与s9可育系小孢子叶球生长期发育的动态变化
通过两个月连续采样,‘松泰s10与s9小孢子叶球生长发育变化趋势见图1。由图1可知,1月中旬前,败育系和可育系小孢子叶球个体生长发育较慢,s10与s9小孢子叶球的孢子长和宽并无明显差异,表明二者在生长速率上差异不明显,生长趋势基本相同;1月下旬,小孢子叶球进入快速生长期,小孢子叶球茎长开始急速增长,s10与s9小孢子叶球的茎长生长量也开始出现差异,且随着生长时间的延长,差异越来越明显,猜测s10小孢子叶球中的小孢子在1月下旬时败育已经发生,其败育开始时间点在1月下旬前,这在形态观察及后期的切片观察实验中得到证实。2月份后,s9小孢子叶球生长放慢,s10小孢子葉球在2月初即停止生长发育,小孢子叶球干枯萎缩,体型缩小,s10与s9小孢子的茎长差异急剧加大,直至2月中旬小孢子叶球脱落时二者的茎长生长量差达到最大值;而s10与s9小孢子叶球的宽长从小孢子叶球的开始生长直到脱落均无显著差异。
2.2 ‘松泰s10败育系与s9可育系小孢子叶球形态发育比较
通过对‘松泰s10败育系与s9可育系小孢子叶球形态发育观察发现,‘松泰s10败育系在1月中旬时,小孢子叶球顶端开始出现干瘪枯散的异常现象,在1月底表现最为明显,且无花粉散出,而同时期s9小孢子叶球发育正常,故推测s10小孢子叶球败育时间开始于1月中旬,即顶端出现干瘪枯散现象时,具体不同时间阶段比较分析如下。12月中旬:小孢子叶球聚集成簇腋生,被鳞片包裹着,鳞片黄褐色,呈卵型(图2: a, s9),与s9形态发育相同,无异常发育(图2: A, s10)。12月底:顶端露出鳞片,小孢子叶呈紫黑色(图2: b, s9)。次年1月初:缓慢发育,呈圆锥形,小孢子叶呈紫黑色(图2: C, s9),顶端露出鳞片,小孢子叶呈紫黑色(图2: B, s10)。1月中旬:缓慢发育,呈圆锥形,小孢子叶呈紫黑色,并出现排列松散、不饱满等异常状态(图2: C, s10)。1月底:小孢子叶球快速生长露出鳞片,小孢子叶成列或交互对生的排列在小孢子叶球中轴上,紧密饱满,呈浅紫色(图2: d, s9),小孢子叶球露出鳞片,生长发育缓慢,小孢子叶排列与可育系相同,小孢子叶球顶端,干瘪枯散更加明显,呈浅紫色(图2: D, s10)。2月初:小孢子叶膨大饱满,小孢子叶球松软,呈黄色,开始散粉(图2: e, s9),小孢子叶球整体呈枯散干瘪状,顶端呈紫红色,底部呈淡黄色,开始脱落,但未见花粉散出(图2: E, s10)。2月中旬:散粉后期,小孢子叶球大量散粉,并开始脱落(图2: f, G, s9),小孢子叶球干瘪萎缩,明显小于可育系,大量脱落且无花粉散出(图2: F, G, s10)。
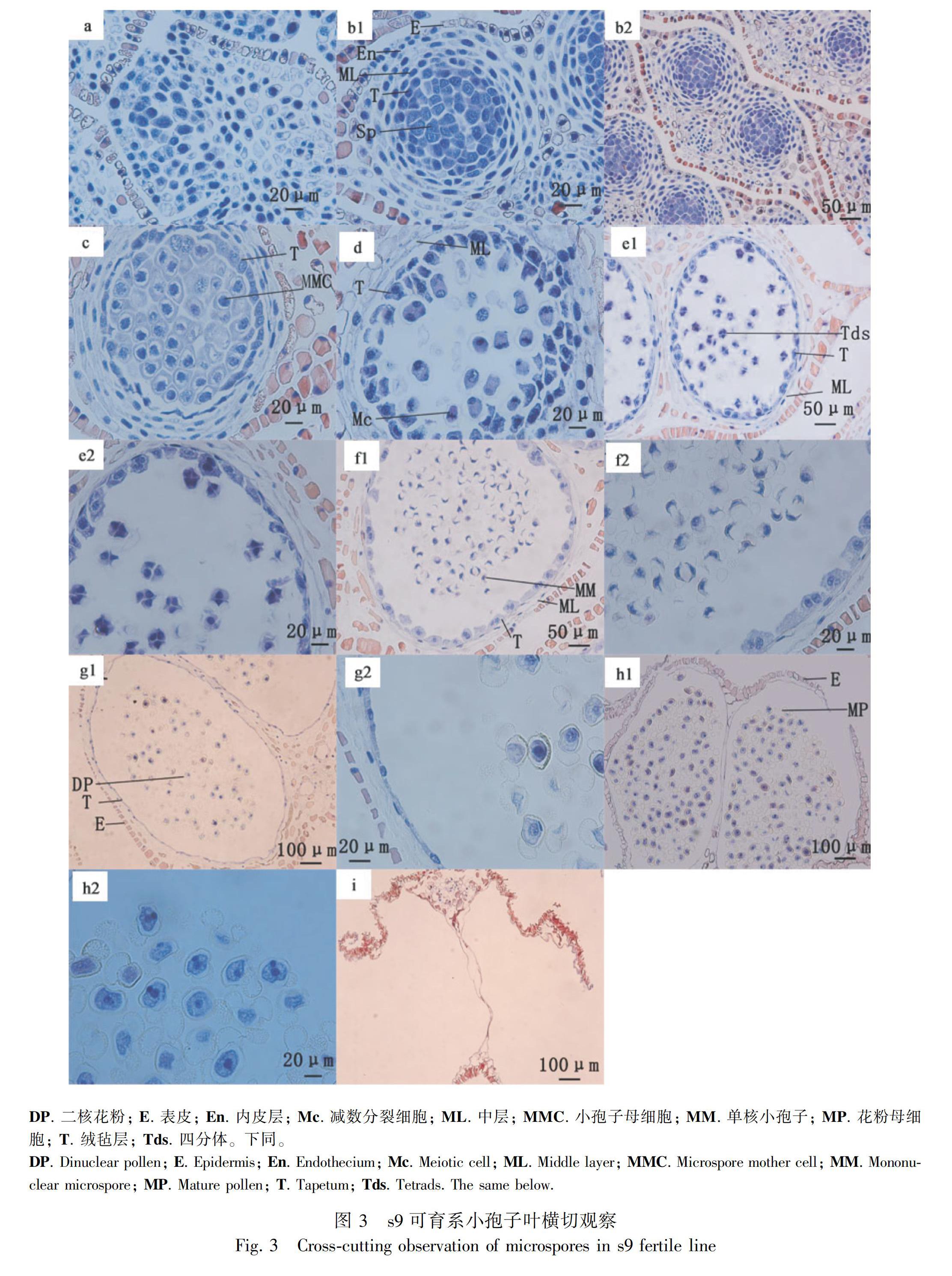
2.3 ‘松泰s10败育系和s9可育系小孢子叶球横切观察
松科植物的花粉(雄性配子体)是在小孢子囊中孕育而来(林金星,2013),小孢子发育过程主要历经初生造孢细胞(primary sporulation tissue period)、造孢细胞(sporulation tissue period)、小孢子母细胞(microsporocyte period)、小孢子母细胞减数分裂(meiotic phase of microsporocyte)、四分体(microspore tetrad period)、单核(mononuclear period)、二核细胞(dinuclear period)、散粉期(loose powder period)等时期(林金星,2013;Sanchez, 2018)。通过对小孢子叶球横切观察(图3,图4)发现,湿地松小孢子叶球从小孢子造饱细胞出现直到散粉完成需要60 d左右;与s9可育系小孢子相比,s10败育系小孢子在四分体时期发育异常,部分小孢子明显萎缩变形,并在后期的发育中同一小孢子叶球的发育情况不一致;小孢子囊壁组织也在四分体时期出现明显差异,s10败育系小孢子绒毡层细胞排列异常、降解缓慢,中层和内壁组织在单核小孢子时期也出现层积,降解缓慢延迟等现象;具体不同发育时期的比较分析如下。
初生造孢组织期:小孢子叶球生长发育初始阶段,小孢子叶还未形成成熟小孢子囊(图3: a, s9),与s9可育系发育情况一致(图4: A, s10)。
造孢组织时期:每个小孢子叶孕育两个小孢子囊腔,呈同心体对称结构。小孢子囊分层清晰,从内到外分别是造胞细胞、绒毡层、中层、内壁和表皮细胞;造胞细胞和绒毡层排列紧密分明,细胞质稠密,整个细胞着色;中层、内壁及表皮细胞围绕绒毡层排列整齐,核大染色清晰(图3: b1, b2, s9),与s9可育系发育情况一致,形成同心体结构,细胞层次清晰(图4: B, s10)。
小孢子母细胞时期:小孢子母细胞体积增大,呈圆形,核大染色深,核周围有一圈不着色区域(空泡化);绒毡层细胞核分裂,出现双核,体积增大,细胞质稠密,着色深,围绕小孢子母细胞排列;中层、内壁细胞拉长,核着色深;表皮细胞核开始消失(图3: c, s9),囊腔体积明显小于同时期s9可育系,其他细胞发育情况与s9一致(图4: C, s10)。
小孢子母细胞减数分裂期:每个小孢子母细胞散落分布在小孢子囊腔内,进行着减数分裂但不同步。绒毡层继续紧紧围绕在小孢子母细胞外(图3: d, s9),小孢母细胞进行减数分裂,小孢子散落分布于囊腔中;绒毡层围绕在小孢子外,被拉长。观察到同一个小孢子叶球,从顶端往下发育阶段不一致,分别是四分体时期、减数分裂中、后时期、母细胞减数分裂间期(图4: D, s10)。
小孢子四分体时期:小孢子母细胞形成四分体,4个小孢子被胼胝质壁包围,散落分布于小孢子囊腔中;绒毡层继续围绕小孢子四分体,染色变浅;小孢子囊壁细胞生长增快,囊腔体积增大明显,中层和内壁细胞被拉长降解,表皮细胞发生角质化加厚(图3: e1, e2, s9),小孢子母细胞分裂形成四分体,4个小孢子被胼胝质壁包围,部分小孢子形态明显萎缩变形;绒毡层在此时期排列不整齐,被拉长,核染色较深;小孢子壁细胞生长较快,囊腔体积增大(图4: E1, E2, s10)。
单核时期:胼胝质释放出小孢子,形成单核小孢子,核染色明显,具有两个气囊;绒毡层染色浅,开始降解;中层及内壁继续降解(图3: f1, f2, s9),单核时期同一小孢子叶球中发育情况不一致。a-c. s9小孢子叶球缓慢发育期; d-e. s9小孢子叶球急速增长; f. s9散粉期; A, B. s10小孢子叶球缓慢发育期; C. s10小孢子叶球顶端出现异常; D-F. s10小孢子叶球异常发育; G. 散粉期对比。
(1)有形态不一的单核花粉形成,有气囊;绒毡层降解明显,排列紊乱;中层、内壁细胞组织层积,降解延迟,细胞核染色可见。(2)无单核花粉形成,囊腔中只有一些形态怪异干瘪萎缩的残留物,形成空囊(图4: F1, F2, s10)。
二核时期:形成二核花粉,气囊膨大明显;绒毡层已经降解但还具有残留物;中层降解完,全部液泡化(图3: g1, g2, s9),同一小孢子叶球中发育情况不一致。(1)单核小孢子发生解体,破碎化,发生败育,但有少部分形态异常2核花粉形成,有气囊;绒毡层已降解,还有部分残留物;中层及内壁细胞膨大液泡化,开始大量降解。(2)小孢子囊腔形成空囊,只见一些残留物(图4: G1, G2, s10)。
散粉初期和散粉后期:囊腔内挤满发育成熟的二核花粉,绒毡层、中层及内壁完全消失,表皮层开裂,开始散粉(图3: h1, h2, s9),后期完成散粉,只见残留囊壁(图3: i, s9),无正常花粉散出,且同一小孢子叶球中发育情况不一致。(1)有部分形态异常成熟花粉形成,气囊萎缩不饱满;绒毡层,中层及内壁细胞消失,表皮不开裂;(2)小孢子囊腔中无花粉形成,只有一些干瘪萎缩的残留物(图4: H1, H2, s10)。
2.4 ‘松泰 s10败育系与s9可育系小孢子叶球及小孢子发育进程比较
结合小孢子叶球形态变化和小孢子石蜡切片观察比较发现,s10败育系和s9可育系在小孢子母细胞减速分裂前并无明显差异,小孢子葉球生长趋势也一致;四分体时期,s10小孢子细胞发育异常,小孢子叶球形态发育也出现异常,二者异常发育具有同步性;从四分体到单核小孢子发育阶段,s10败育系与s9可育系差异最为显著,s9小孢子叶球在这一阶段快速增长,发育迅速,直至散粉,s10小孢子叶球则增长缓慢,发育延迟,出现干瘪萎缩现象,直至脱落;因此,推测四分体到单核小孢子发育阶段是s10小孢子败育发生的主要时期。二者具体发育进程比较见表1。DP. 二核花粉; E. 表皮; En. 内皮层; Mc. 减数分裂细胞; ML. 中层; MMC. 小孢子母细胞; MM. 单核小孢子; MP. 花粉母细胞; T. 绒毡层; Tds. 四分体。下同。
3 讨论
对湿地松‘松泰小孢子叶球发育过程的研究结果表明:从1月中旬开始,小孢子叶球顶端开始出现干瘪松散异常现象,该时期的一个小孢子叶球从顶端到底部,小孢子发育时期不一致(顶端为四分体、中部为减数分裂中、后时期、底部为减数分裂间期),并在四分体释放单核小孢子这一时期出现异常,造成败育。因此研究认为‘松泰小孢子叶球败育时期开始于四分体时期,这与Kaul(1988)的研究结果类似, Kaul (1988)对多种雄性不育类型花粉败育时期进行统计发现,花粉败育在造胞细胞至二核花粉各时期均有可能发生,其中有70%的材料败育时期发生在四分体以后。而在四分体后续的发育过程中,同一个小孢子叶球中小孢子的发育分为两种趋势,一种是在小孢子叶球中上部分,胼胝质不能释放单核小孢子,没有单核小孢子的形成,直接降解萎缩,与绒毡层一起形成絮状物,形成空囊,直至小孢子叶球脱落,囊壁表皮不开裂;另外一种发育趋势则是在小孢子叶球的中下部分,四分体在胼胝质解体后,可以释放出小孢子,形成异常单核小孢子继续发育。这种同一个小孢子叶球中存在两种不同的败育方式,在杉木中也有发现(林金星等,2013)。而吕洪飞和余象煜(1999)在对杉木雄性不育细胞学研究中发现同一个小孢子叶球中同时存在无小孢子囊型、减数分裂I期前中层细胞增生型、减数分裂I期后中层细胞增生3种败育类型,该研究认为小孢子囊壁的中层、内壁出现层积和降解延迟、表皮不开裂的异常发育现象也会影响小孢子的后续发育,造成败育发生。
绒毡层是构成花药壁的最内层组织结构,围绕着小孢子分布,在物质代谢合成与分泌上都十分活跃,含有丰富的营养物质,是物质输送的最后中转站,对花粉的正常发育具有关键作用(范彦君等,2016;Tian et al., 2017)。本次研究结果显示,‘松泰小孢子叶球在四分体时期之前与可育系相比并无异常,但在四分体时期,小孢子叶球开始出现异常,其胼胝质溶解缓慢,绒毡层及中层细胞降解延迟;可育系从四分体到单核小孢子发育阶段的时间为5 d左右,而败育系持续发育长达20 d左右,持续时间为可育系的4倍。在此期间,败育系的绒毡层与可育系相比发育缓慢,降解延迟,导致其不能及时的分泌和转化小孢子发育所需能量物质,造成败育发生,这与唐国强(2015)对湿地松小孢子叶球败育研究结果一致,刘望舒等(2019)在对杨树雄性不育小孢子败育过程的研究中也认为绒毡层的延迟降解会导致小孢子发生败育;谭智文等(2013)与李六林(2007)分别对不同梨品种有关小孢子败育过程的研究中也发现存在绒毡层、中层细胞的异常发育现象;这种现象在小麦(赵卜等,2015)、棉花(孔祥军,2017)等其他植物有关雄性不育的细胞学观察中,也有发生。同时,败育系胼胝质降解缓慢,不释放或不能及时的释放单核小孢子,直接影响到小孢子的正常发育;而胼胝质合成于母细胞减数分裂时期,降解于四分体释放小孢子时期,在这过程中它的合成、积累、抑或降解,任何一个步骤出现异常,都可能会造成败育的发生( Down et al., 2008; Wan et al., 2011;杨莉芳和刁現民,2013)。
4 结论
小孢子发育过程的细胞学观察是了解湿地松雄性败育途径、方式的重要研究手段,也是湿地松雄性不育家系在杂交育种优势利用中的研究基础。本次研究以‘松泰 s10败育系和s9可育系为材料,通过对其小孢子叶球形态和细胞发育动态变化观察发现,s10败育系和s9可育系在小孢子母细胞减速分裂前无明显差异,小孢子叶球生长趋势也一致;但在四分体时期,‘松泰小孢子细胞组织发育异常,同时在小孢子叶球形态上也能观察到干瘪枯散现象,表明其外部形态变化和细胞异常发育具有同步性;此外,败育系从四分体到单核小孢子阶段与可育系相比发育时间较长,且在此期间败育系出现小孢子绒毡层细胞发育异常、降解缓慢,小孢子囊壁组织排列紊乱、降解延迟等现象,最后,s10形成异常二核花粉,且无花粉散出。因此推论s10小孢子败育的原因主要为小孢子囊壁细胞发育异常,其小孢子叶球形态异常,相对应的其绒毡层在四分体时期发育异常,不能适时地分泌胼胝质酶来降解围绕着四分体的胼胝质壁,也不能适时地输送花粉形成所需能量物质,同时囊壁细胞出现降解延迟和层积。这一系列的异常变化导致不能形成正常四分体,从而使花粉败育。
参考文献:
DOWN T, RAKYAN V, TUMER D, et al., 2008. A bayesian deconvolution strategy for immunoprecipitation-based DNA methylome analysis [J]. Nat Biotechnol, 26(7):779-785.
FAN YJ, WANG Y, LIU QY, et al., 2016. Advances in cytoplasmic male sterility in plant [J]. Chin Agric Sci Bull, 32(18): 70-75. [范彦君, 王瑜, 刘齐元, 等, 2016. 植物细胞质雄性不育研究进展 [J]. 中国农学通报, 32(18): 70-75.]
KAUL M, 1988. Male sterility in higher plants [M]. Springer Berlin Heidelberg.
KONG XJ, 2017. Studies on cytology and molecular biology of male sterility line H276A in Gossypium barbadense [D]. Nangning: Guangxi University. [孔祥军, 2017. 海岛棉H276A雄性不育的细胞学与分子生物学基础研究 [D]. 南宁:广西大学.]
LIN JX, HU YX, WU H, 2013. The biology of pollen in gymnosperms [M]. 1st ed. Beijing: Science Press: 37-40. [林金星, 胡玉熹, 吴鸿, 2013. 裸子植物花粉生物学 [M]. 1版. 北京:科学出版社: 37-40.]
LI LL, 2007. Study on the cytological and physiological characteristics of pollen abortion of ‘Niitaka (Pyrus pyrifolia Nakai) [D]. Nanjing: Nanjing Agricultural University. [李六林, 2007. ‘新高梨花粉败育的细胞学和生理特性研究 [D]. 南京:南京农业大学.]
LIU WS, HAN LZ, ZHU W, et al., 2019. Cytological observation on pollen development of the male sterile poplar variety [J]. J Nanjing For Univ (Nat Sci Ed), 43(1): 198-203. [刘望舒, 韩林芝, 朱嵊, 等, 2019. 杨树雄性不育品种花粉发育过程的细胞学观察 [J]. 南京林业大学学报(自然科学版), 43(1): 198-203.]
L HF, YU XY, 1999. Change of callose wall and spropollenin of microsporangium abortion in Cunninghamia lanceolata [J]. J NW Bot, 19(1): 108-112. [吕洪飞, 余象煜, 1999. 杉木小孢子囊败育过程中的胼胝质壁和孢粉素的消长 [J]. 西北植物学报, 19(1): 108-112.]
PAN ZG, YOU YT, 1994. Introduction and cultivation of major exotic tree species in China [M]. Beijing: Science and Technology Press: 79-92. [潘志刚, 游应天, 1994. 中国主要外来树种引种栽培 [M]. 北京: 科学技术出版社: 79-92.]
SANCHEZ D, SONIA V, 2018. Embryology of Mammillaria dioica (Cactaceae) reveals a new male sterility phenotype [J]. Flora, 241: 16-26.
TAN ZW, JIN BK, CAO HN, et al., 2013. Study on male sterility cytology of small perfume pear in Yanbian [J]. J Agric Sci Yanbian Univ, 35(1): 28-31. [谭智文, 金炳奎, 曹后男, 等, 2013. 延边小香水梨雄性不育细胞学研究 [J]. 延边大学农学学报, 35(1): 28-31.]
TIAN Y, XIAO S, LIU J, et al., 2017. MALE STERILE6021 (MS6021) is required for the development of anther cuticle and pollen exine in maize [J]. Sci Rep, 7(1): 16736.
TANG GQ, 2015. Study on the growth and physiological characteristics of Pinus elliottii male sterile clone (SG001) [D]. Nanning: Guangxi University. [唐国强, 2015. 湿地松雄性不育无性系(SG001)生长及生理特性研究 [D]. 南宁:广西大学.]
WAN L, ZHA W, CHENG X, et al., 2011. Arice β-1,3-glucanase gene Osg1 is required for callose degradation in pollen development [J]. Plant, 233(2): 309-323.
WISE R, PRING D, 2002. Nuclear-mediated mitochondrial gene regulation and male fertility in higher plants: Light at the end of the tunnel ? [J]. Proc Natl Acad Sci, 99 (16): 10240-10242.
YANG LF, DIAO XM, 2013. Progress in identification of plant male sterility related nuclear genes [J]. J Plant Gene Resour, 14(6): 1108-1117. [楊莉芳, 刁现民, 2013. 植物细胞核雄性不育基因研究进展 [J]. 植物遗传资源学报, 14(6): 1108-1117.]
YE BX, BI JJ, SUN YS, 2011. Research methods of plant cell and tissue [M]. Beijing: Chemical Industry Press. [叶宝兴, 毕建杰, 孙印石, 2011. 植物细胞与组织研究方法 [M]. 北京:化学工业出版社.]
ZHAO B, ZHANG XZ, ZHENG YJ, et al., 2015. Cytological observation on meiosis and development of microspores in F-type male sterile wheat line [J]. J Triticeae Crop, 35(7): 918-925. [赵卜, 张向展, 郑炜君, 等, 2015. F型小麦雄性不育系小孢子发育的细胞学观察 [J]. 麦类作物学报, 35(7): 918-925.]
ZHANG SN, 2017. Study on the evaluation technique of wood property and multiple trait selection of Pinus elliottii [D]. Beijing: Chinese Academy of Forestry. [张帅楠, 2017. 湿地松材性测定技术研究与多样性联合选择 [D]. 北京:中国林业科学研究院.]
ZHENG WZ, FU LG, 1978. Flora Reipublicae Popularis Sinicae [M]. Beijing: Science Press, 7: 273-275. [郑万钧, 傅立国, 1978. 中国植物志 [M]. 北京: 科学出版社, 7: 273-275.]
ZHU ZS, 1993. Pinus elliottii [M]. Guangzhou: Guangdong Science and Technology Press, 3: 1-4. [朱志淞, 1993. 湿地松 [M]. 广州:广东科技出版社, 3: 1-4.]
(责任编辑 周翠鸣)
