紫距淫羊藿的形态变异式样及描述修订
2020-05-26何益明黄华蒋勇李仁清徐艳琴
何益明 黄华 蒋勇 李仁清 徐艳琴

摘 要: 紫距淫羊藿为根据少量栽培个体描述的中国特有种,狭域分布于湖南省天平山,自发表以来未见后续形态学研究。为了掌握紫距淫羊藿形态变异式样并为该物种及其近缘种的分类提供依据,该研究在模式产地湖南省桑植县天平山,于花果期开展野外居群的形态观测,对紫距淫羊藿主要性状进行变异分析以及形态描述的完善或修订,并提供了该物种的形态图片资料。结果表明:基于野外观测,紫距淫羊藿主要形态性状的变异范围均远超出物种发表原始文献的描述,如株高12.0~68.0 cm,大幅度高于原始描述的12 cm;居群内结节状根茎和细长根茎并存,且两种类型的个体数量相当;其余性状,如叶长、叶宽、花序轴长、花色等都呈现丰富变异。此外,还补充描述了紫距淫羊藿的果实和种子的性状,蓇葖果长8~13 mm,内含种子2~11粒,种子具油质体。因此,分类学描述应重视野外观察,特别要注意个体间的变异并进行性状分析,这是分类研究的基础。
关键词: 紫距淫羊藿, 特有种, 形态变异, 补充, 修订
中图分类号: Q949 文献标识码: A
文章编号: 1000-3142(2020)04-0452-10
Abstract: Epimedium epsteinii, a species endemic to China, and distributes in Tianping Mountain of Hunan Province, which was described based on several cultivated individuals that. No subsequent morphological observation has been conducted since its publication. In order to obtain the accurate and comprehensive morphological variation of E. epsteinii and to provide a basis for clarifying the taxonomic relation of this species and its allied taxa, morphological variation analysis and supplementary description of E. epsteinii were done through the field investigation conducted in flowering period, based on a population in Tianping Mountain (type locality) of Sangzhi County, Hunan Province. The results are as follows: The variation range of the main characters of E. epsteinii is far beyond the original descriptions. For example, the height of flowering stems is 12.0-68.0 cm, which is significantly higher than the original description of 12 cm; Both short, compact rhizomes and long creeping rhizomes are observed within the population, and the individuals of each type are almost equal in number; The other characters, such as length and width of middle leaflet, length of inflorescence and flower colour are present abundant variation; In addition, the fruit and seed characters were described for the first time. Follicles are about 8-13 mm long and containing 2-11 seeds, which with a large oily caruncle (elaisome) attached. Furthermore, photos on the morphology of living plants in the wild are provided to fill the gap of absence of colour illustration of E. epsteinii in extant literature. It is the first very important thing for taxonomic study to pay more attention to taxonomic description after field investigation, especially the analysis of individual variation.
Key words: Epimedium epsteinii, endemic species, morphological variation, supplementary description, revision
正確的物种概念来自对植物形态变异式样及其分类学价值的正确且充分理解(徐炳声,1998)。观察尽可能多的样本,充分研究其变异性是分类研究的基本功(Nooteboom,1992;洪德元,2016)。由于缺乏广泛的野外居群观察,对居群水平的变异缺少了解,中国的植物区系中大量类群的真实身份还有讨论余地(徐炳声,1998),大量种级水平的分类学问题有待解决(杨亲二,2016)。
淫羊藿属(Epimedium L.)隶属于小檗科(Berberidaceae),是分类上争议较多的类群(Zhang et al., 2014; Guo et al., 2018)。该属目前报道约60种,其中85%为中國特有(Xu et al., 2019)。中国类群的数量近三十年来迅速增长,从1975年的13种(应俊生,1975)增加到目前约50种,但近一半的物种为外国研究者命名(徐艳琴等,2014),其中19种为根据栽培于国外的少量栽培个体命名。这些个体无法代表一个居群,更无法涵盖一个物种的全部变异,对性状变异未能进行可靠的比较和分析。由于缺乏对自然生境个体的观察,导致重要生物学特性难以得到客观和全面反映,对物种的形态变异幅度调查还不够,对性状的分类学价值把握不准。致使淫羊藿属分类的难点主要集中在中国类群(Zhang et al., 2014, 2015; 徐艳琴等,2014;刘少雄等,2016; Liu et al., 2017; Xu et al., 2019)。Buck(2003)明确指出淫羊藿属有些种类可能过分强调了某些差异,从而忽视了物种自身的变异范围。
对淫羊藿属的分子系统学研究表明,SSR(Yousaf et al., 2015)、AFLP(De Smet et al., 2012; Zhang et al., 2014)、ITS(Zhang et al., 2007; De Smet et al., 2012; Guo et al., 2018)、atpB-rbcL(Zhang et al., 2007)、matK(De Smet et al., 2012)、rbcL(Guo et al., 2018)和psbA-trnH(Guo et al., 2018)等广泛应用的DNA标记和条形码技术,均无法满意地解决淫羊藿属物种界定问题。形态学特征与地理分布式样分析却给物种区分提供了重要依据(Zhang et al., 2014, 2015; 刘少雄等,2016; Liu et al., 2017)。这可能与淫羊藿属存在自然杂交及基因渐渗有关,也提示淫羊藿属分类难题的解决和系统关系的清晰掌握,首先要进行足够的性状调查和分析。
紫距淫羊藿(Epimedium epsteinii)为中国特有种,由北京植物园于1994年采自湖南天平山,Darrell Probst先将其引种栽培于美国马萨诸塞州Hubbardston,Mass.,之后栽培于英国Blackthorn Nursery,Kilmeston,Hampshire,1997年Stearn(1997)根据英国栽培植株命名。尽管Stearn对紫距淫羊藿的描述相对完整,但所依赖的样本数量非常有限,无法体现个体间变异及变异式样,且缺乏果实性状描述。此外,多项研究表明,不同环境及移栽会对淫羊藿的生长和形态(如叶长、长宽比、株高和叶片数量等)产生较大影响(孙超等,2004;高辉等,2012;权秋梅等,2012)。因此,开展自然生境下的调查和形态观测对掌握物种的形态特征非常必要。但是,该物种自发表以来,尚未见相关研究。2001年,《中国植物志》收录紫距淫羊藿,对原始形态描述几乎未做修改,只是将“株高12 cm或更长(flower stem 12 cm or more long)”修改为“株高约15 cm”(应俊生,2001)。2002年,Stearn(2002)在淫羊藿属专著中,紫距淫羊藿沿用原始描述。2011年,Flora of China中,紫距淫羊藿的性状为《中国植物志》相应内容的英文翻译(Ying et al., 2011)。何顺志(2014)在《中国淫羊藿属植物彩色图鉴》中,提供了绝大多数物种的彩色图片,但紫距淫羊藿只有一幅墨线图,形态特征描述完全为《中国植物志》的转载。到现在为止,所有关于紫距淫羊藿的形态描述其实都仅限于Stearn(1997)基于少量异国栽培个体的观测。
关于紫距淫羊藿的分布,一直被认为狭域分布于天平山。除了模式标本Darrell Probst 940255以外,检索国家数字标本馆和国内外各大标本馆,目前仅在中国医学科学院药用植物研究所标本馆(IMD)查询到一份栽培于湖北省恩施市长岭岗药材场的标本(郭宝林和刘京晶0812)。据标本记载是由附近移栽而来,但该分布点暂未得到其他研究证实,本研究团队虽在恩施开展过多次淫羊藿属植物资源调查但未发现有紫距淫羊藿分布,恩施分布点有待确认。徐艳琴等(2016)曾报道紫距淫羊藿的新分布,但后续研究发现这两个居群的形态与紫距淫羊藿存在一些差异,鉴定存在争议(待发表资料),也有待进一步观察和确认。因此,紫距淫羊藿目前无争议的分布区仍然仅限于天平山。
2018年4月,本研究在紫距淫羊藿的模式产地湖南省张家界市桑植县天平山林区开展了野外调查工作。对野外植株进行全面观测和分析,发现在一些重要分类学形状上,紫距淫羊藿形态特征的变异范围远超出原始描述。因此,本文对紫距淫羊藿的形态学描述进行完善和修订,并补充果实和种子的形态描述以及植物形态图片资料。本研究结果将为紫距淫羊藿的物种界定、种间关系研究及其资源利用等提供科学依据。
1 材料与方法
1.1 材料
由于花部特征对淫羊藿属植物的分类具有重要意义,所以本研究于花果期开展。2018年4月下旬,在湖南省张家界市桑植县天平山林区,调查紫距淫羊藿的野外居群,采集植物标本,记录并拍摄形态特征。腊叶标本存放于江西中医药大学中药资源植物标本室(JXCM)中。
1.2 方法
在居群中选择20个开花植株进行形态观测和统计。用卷尺测量株高(茎基部至花序顶端的高度)和花序轴长,用直尺测量中间小叶叶长和中间小叶叶宽,用游标卡尺测量根茎直径、小花直径和花梗长、外萼长和宽、内萼长和宽、距长。统计花茎上叶的数量、小花数、结实率(结果数/小花数)、果实长度和种子数,共获得17个数量性状。同时,记录花序类型、花茎上叶的着生方式、花序轴被毛、花梗被毛、叶背毛、花各部分(外萼、内萼、花瓣和花药)颜色和根茎类型(结节状或细长)等10个质量性状。
首先,利用SPSS 19.0软件统计数量性状的平均值及标准偏差。然后,计算变异系数(CV)=标准差/平均数。最后,将本研究观测结果与原始描述进行比较分析。
2 结果与分析
2.1 生境概述
天平山林区为湖南省八大公山自然保护区的核心区域,地处武陵山脉北麓,北与湖北省鹤峰县交界。该区位于云贵高原东侧和鄂西山地南端,峰峦林立,植被丰富。该调查居群的地理位置为110°05′26″ E、29°46′59″ N,海拔为1 380 m。紫距淫羊藿稀疏分布于约300 m2的狭小区域,多生于林缘或灌丛。主要伴生植物包括盐肤木(Rhus chinensis)、鹿角杜鹃(Rhododendron latoucheae)、金樱子(Rosa laevigata)、山莓(Rubus corchorifolius)、灯台莲(Arisaema sikokianum)、鹿药(Smilacina japonica)、龙头草(Meehania henryi)、独蒜兰(Pleione bulbocodioides)、虎耳草(Saxifraga stolonifera)、一把伞南星(Arisaema erubescens)、三脉紫菀(Aster ageratoides)、唐古碎米荠(Cardamine tangutorum)和毛茛(Ranunculus japonicas)等。
2.2 紫距淫羊藿主要性状的变异式样
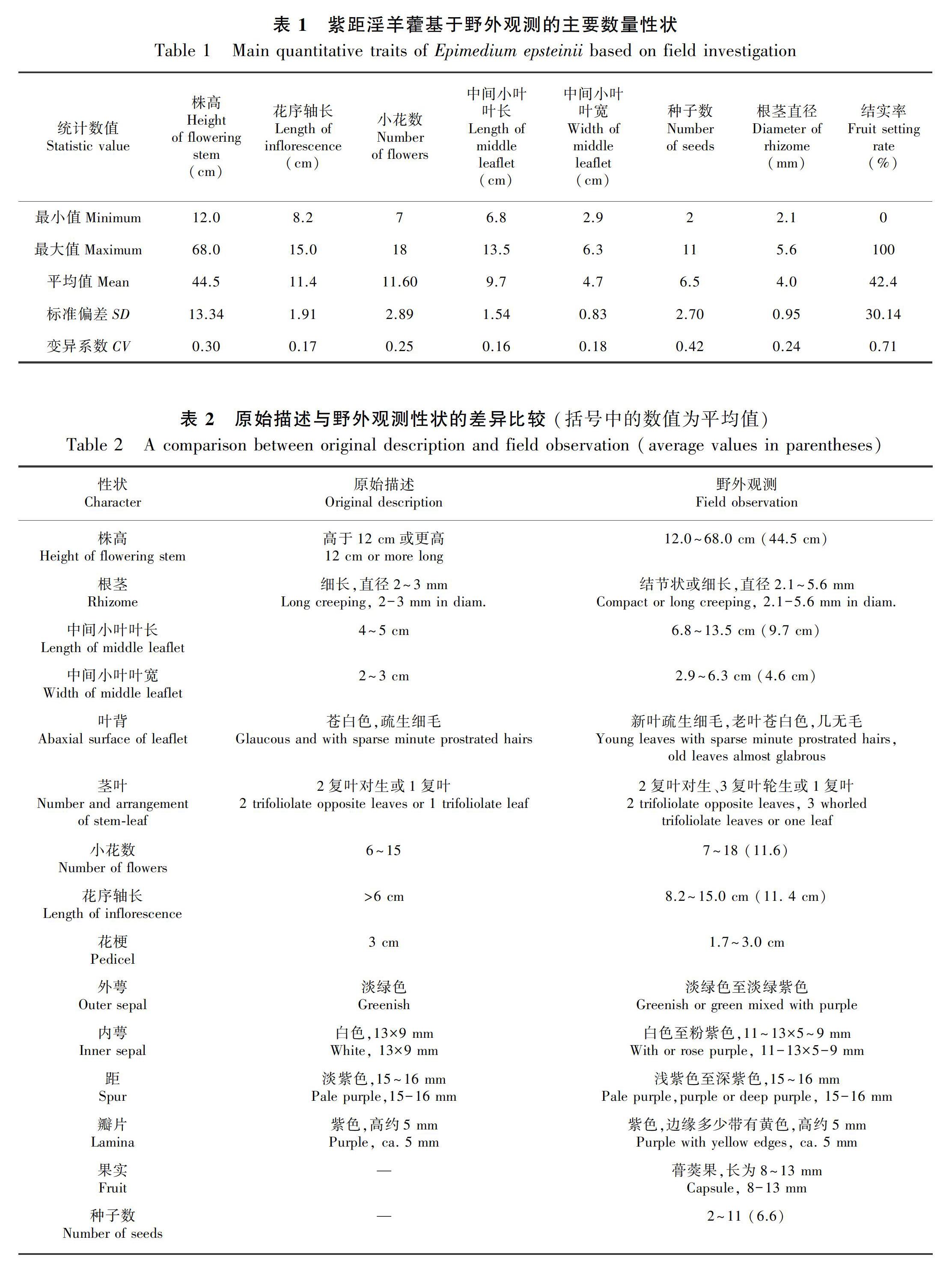
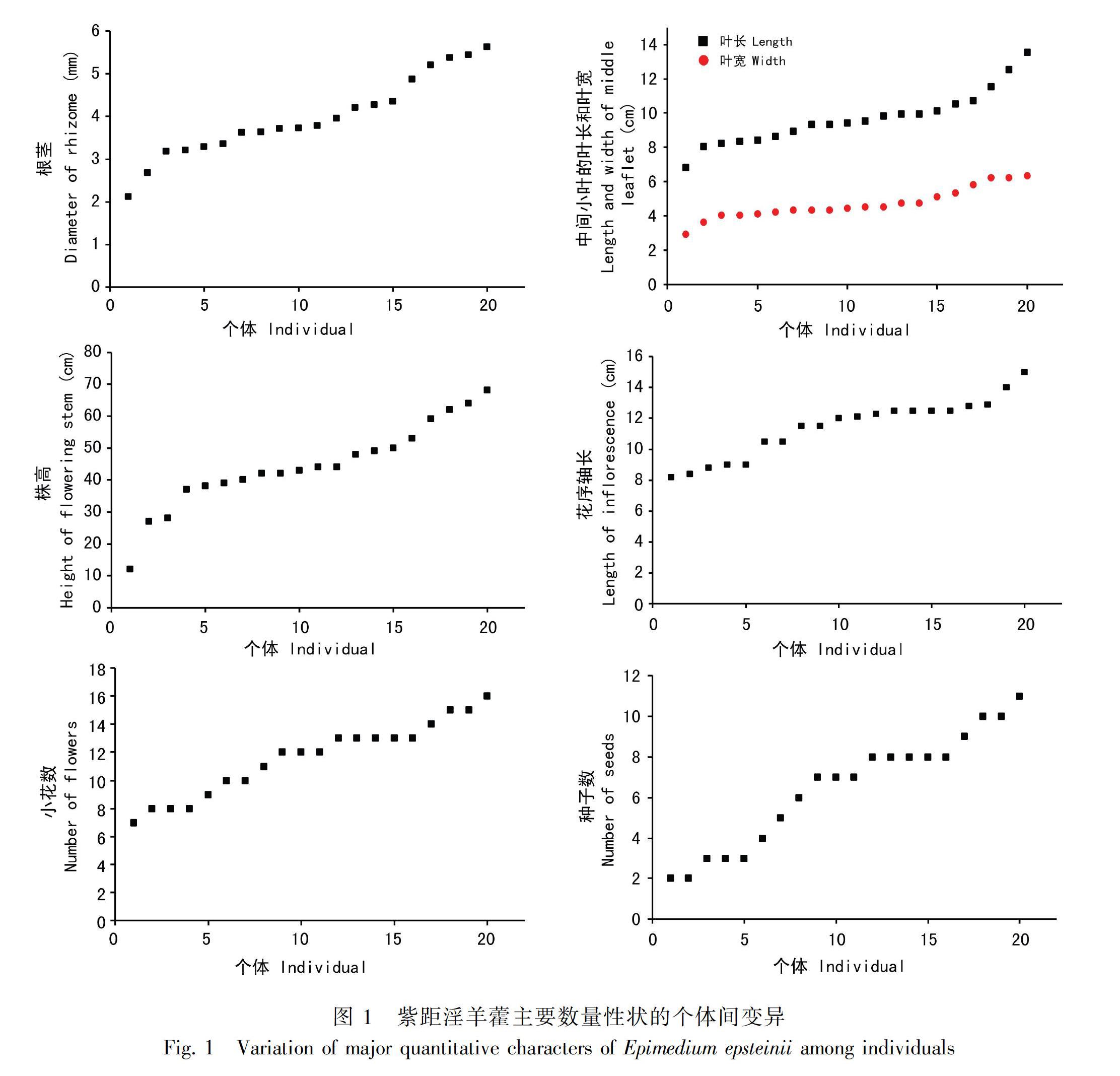
2.2.1 株高 紫距淫羊藿的株高变异范围较大。株高最小为12.0 cm,最高达68.0 cm,平均为44.5 cm。近半数的个体(9个)集中在40.0~50.0 cm之间。40.0 cm以下和50.0 cm以上的个体分别为6个和5个。株高在个体间整体呈连续变异(表1,图1)。
2.2.2 根茎 观察到两种根茎类型,结节状或细长。结节状的9株,细长的8株,两种类型并存的3株。细长的根茎延伸可长达19.0 cm。根茎直径从2.1 mm到5.6 mm连续变异,半数的个体(10个)集中在3.0~4.0 mm之间,3.0 mm以下和5.0 mm以上的个体分别为2个和4个(表1,图1,图2)。
2.2.3 叶片 一回三出复叶,小叶革质,狭卵形,先端急尖至渐尖。花茎多为2复叶对生,偶3复叶轮生或仅1枚复叶。幼叶绿色、紫红色或具紫红色斑块,紫红色会随叶片长大逐渐变为绿色。新叶(当年生叶片)叶背浅绿色,疏被细伏毛,老叶(往年生叶片)叶背苍白色,几乎无毛。中间小叶叶长为6.8~13.5 cm,多集中在9.0 cm左右,叶宽为2.9~6.3 cm,多集中在4.5 cm左右。叶长和叶宽在个体间呈连续变异(图1)。
2.2.4 花部特征 总状花序,偶基部1~2个花梗具2~3朵小花。花序轴仅幼嫩时疏被腺毛,后几乎光滑无毛。花序轴长为8.2~15.0 cm,平均11.4 cm。近半数的个体(9个)集中在12.0~13.0 cm之间,个体间呈连续变异(表1,图1,图2)。小花7~18朵,平均11.6朵,多集中在10~13朵(表1,图1)。小花花梗密被腺毛,结果后逐渐变疏,花梗长为1.7~3.0 cm。花大,直径约3 cm。外萼片狭卵形,淡绿色至淡绿紫色,早落,内萼片狭卵形至卵形,11~13 × 5~9 mm,白色至粉紫色。花瓣长于内萼片,距明显弯曲或稍弯曲,淡紫色、紫色或深紫色,长为15~16 mm,距先端和基部瓣片紫黄色、浅紫色或紫色,瓣片边缘黄白色、黄色或紫色,高约5 mm(图2)。
2.2.5 果实和种子 果实为蓇葖果,长为8~13 mm,宿存花柱约5 mm,内含种子2~11粒,种子一端具有白色附属物,内含油质体(表1,图1,图2)。统计20个花序的自然结实率为0%~100%,平均42.4%(表1)。
2.2.6 主要性状的变异系数 紫距淫羊藿主要数量性状的变异系数差异明显,种子数的变异系数最大(0.42),根茎、小花数和株高的变异系数介于0.24~0.30之间,花序轴长、中间小叶的叶长和葉宽变异系数较为接近,为0.16~0.18。
2.3 野外观测与原始描述比较
基于20个个体的野外形态观测,本研究结果与原始文献描述性状存在较大差异(表2)。整体而言,野外观测的形态性状呈现更广泛变异。如原始描述株高“高于12 cm或约15 cm”,但野外观测远高于12 cm。原始描述“根茎细长,直径2~3 mm”,野外观测其根茎结节状或细长并存,直径为2.1~5.6 mm。原始描述对叶色未做特殊说明(即为绿色),野外观测叶片颜色在幼叶、成叶和老叶等不同阶段可能有所不同,幼叶绿色、紫红色或具紫红色斑块,长成后逐渐变绿,老后暗绿色。中间小叶叶长(6.8~13.5 cm)和叶宽(2.9~6.3 cm)均远大于原始描述(4~5 × 2~3 cm)。叶背被毛特征受发育阶段影响,当年生叶背面疏生细毛,老叶几乎无毛。野外观查小花各部分颜色较原始描述呈现更丰富的变化,内萼片白色至粉紫色,距浅紫色至深紫色变异。花茎上叶的数量及着生方式除原始描述的2复叶对生或1复叶,还观察到有3复叶轮生。小花数、距长和瓣片高度与原始描述较为接近。
2.4 形态特征补充描述
紫距淫羊藿
Epimediumepsteinii Steam in Kew Bull. 52(3): 662, Fig. 3, 1997;中国植物志,29: 283. 2001;Flora of China, 19: 793. 2011;中国淫羊藿属植物彩色图鉴,96. 2014;Guihaia, 36(5): 619. 2016.
多年生草本,植株高为12~68 cm。根茎结节状或细长,直径为2.1~5.6 mm,多须根。一回三出月,果期4月—5月。
产于湖南省张家界市桑植县天平山,生于海拔1 400 m左右的林缘灌丛。
标本引证, 湖南:天平山,Probst 940255,1997-4-5,alt. 400~1 000 m(Holotype,K!);徐艳琴等2018003,2018-4-24,alt. 1 380 m(JXCM!)。湖北:恩施市,郭宝林和刘京晶0812,2008-4-13,alt. 1 670 m(IMD!)。
3 讨论
本研究基于紫距淫羊藿模式产地(湖南省张家界桑植县天平山)居群20个个体的形态观测和分析,较之原始描述,各性状都呈现出更广泛的变异。差异最突出的性状是株高,Stearn(1997)描述“株高12 cm或更高(flower stem 12 cm or more long)”,《中国植物志》(应俊生,2001)和Flora of China(Ying et al., 2011)将其修订为“约15 cm(ca. 15 cm tall)”。但是,本研究仅观测到一个株高12 cm的个体,其余均远高于此,最高可达68 cm,平均株高44.45 cm。分析其原因,一方面是由于原始描述观测的个体数量有限;另一方面的重要因素可能是栽培环境改变对性状的影响。研究表明,淫羊藿属植物栽培后,其叶片大小和株高等性状与野生个体存在较大差异,如巫山淫羊藿栽培后,株高仅为野生植株的一半,叶片明显变小(孙超等,2004)。对淫羊藿属近40个物种的园地栽培观测发现,栽培后植株普遍更矮小。提示淫羊藿属植物准确的形态描述应基于自然生境植株,栽培后,尤其是异国栽培后,其数量性状可能发生较大改变。因此,淫羊藿属依据少量移栽至国外植株命名的物种(19种)(徐艳琴等,2014),均有必要开展自然生境下的形态观察和描述。实际上,后来研究已经证实,这其中的多数类群存在描述不准确、不全面或分类处理欠妥当等问题(Zhang et al., 2014; Zhang et al., 2015; 刘少雄等,2016; Liu et al., 2017; Xu et al., 2019)。已對8个类群开展了修订或补充描述,如黔岭淫羊藿(Zhang et al., 2015; Xu et al., 2019)、直距淫羊藿(Xu et al., 2019)、绿药淫羊藿(Zhang et al., 2015; Xu et al., 2019)、钟花淫羊藿(Liu et al., 2017)、木鱼坪淫羊藿(刘少雄等,2016)、时珍淫羊藿(刘少雄等,2016)、强茎淫羊藿(Zhang et al., 2015)和短茎淫羊藿(Zhang et al., 2015)。
紫距淫羊藿原始描述根茎细长(long creeping)(Stearn, 1997),野外观测其根茎结节状或细长并存于居群内,且两种类型的个体数量相当,甚至有3个个体同时存在结节状和细长两种类型。根茎结节状细长,曾作为淫羊藿属一个重要的分类性状,以往认为淫羊藿不同类群的根茎类型固定,非此即彼(应俊生,2001; Stearn, 2002; Ying et al., 2011)。但是随着研究的深入和观察个体的增加,粗毛淫羊藿(Zhang et al., 2015)、黔岭淫羊藿(Zhang et al., 2015)、时珍淫羊藿(刘少雄等,2016)和腺毛淫羊藿(Xu et al., 2019)等类群都已报道根茎类型在居群间或个体间可能存在过渡和变异。因此,根茎性状的分类价值不能一概而论,不同类群应区别对待。
在紫距淫羊藿的狭域分布区内,尽管仅观察到20个开花植株,但本研究依然发现了较明显的花色变异。本研究中,内萼片从白色到粉紫色变异,距从淡紫色、紫色到深紫色变异,基部瓣片紫色,先端多少有一圈黄白色至黄色的边缘。Stearn(1997)的原始描述仅记载了白色内萼片和紫色花瓣,为本研究结果的子集。根据Stearn(2002)紫距淫羊藿的彩色图版,其瓣片为纯紫色,是本研究没有观察到的变异。暗示个体在相当程度上能反映物种的性状,但不能完全代表物种,只有足够多的样本观测才能获得物种较完善的性状变异幅度(陈家宽和王徽勤,1986;徐炳声,1998)。此外,本研究结果还说明淫羊藿属植物的花色变异可能普遍存在,这与作者前期的研究结果一致(Xu et al., 2019)。
关于果实性状,本研究在模式标本上有较完整的果实,但Stearn(1997)未进行描述,原因不详。本研究中,选择花果期进行调查,对果实性状进行补充描述。蓇葖果长为8~13 mm,宿存花柱长约5 mm。果实有2~11粒种子。关于淫羊藿属植物的果实类型,不同文献有不同描述,主要观点有蒴果和蓇葖果。《中国植物志》(应俊生,2001)和Flora of China(Ying et al., 2011)均描述其为蒴果(capsule)。石旭等(2010)对7种淫羊藿属植物进行解剖和观察后,将该属果实类型修订为蓇葖果(follicle)。不同版本的《药用植物学》教材也认为淫羊藿属植物为蓇葖果(谈献和和王德群,2013;刘春生,2016)。本研究仔细观察紫距淫羊藿果实性状,具1心皮、边缘胎座,为蓇葖果。
虽然大量淫羊藿属的分类研究已开展,但仍然存在较多问题,根本原因是近半数物种的命名是依据少量异国栽培植株,大多数物种缺乏广泛深入的性状调查,各类群性状变异幅度、变异式样和分类价值的研究不够。在样本数量不足的情况下,即使是一个连续的变异式样也会呈现间断的形式,连续变异性状中的若干子集,可造成分类上的困难甚至错误(徐炳声,1998)。变异是生物界最基本的特性之一,只有对性状的变异规律和变异模式分析到位,才能正确判断性状的分类价值,才有合理的物种划分(洪德元,2016)。如果要认识性状变异的规律,评估其在类群划分中的价值,就应充分重视“居群”概念,准确理解和处理物种内的个体变异和变异幅度(陈家宽和王徽勤,1986)。尽可能进行覆盖类群分布区的居群样本分析,这是分析性状变异的前提(徐炳声,1998;陈家宽,2016),也是解决淫羊藿属这类困难类群分类问题的前提和基础。
致谢 感谢中国医学科学院药用植物研究所标本馆(IMD)和英国皇家植物园标本馆(K)提供标本资料;感谢审稿专家对本稿件提出的宝贵修改意见。
参考文献:
BUCK WR, 2003. The genus Epimedium and other herbaceous Berberidaceae; Including the genus Podophyllum [J]. Brittonia, 55: 302-304.
CHEN JK, 2016. Population, species and biodiversity [J]. Biodivers Sci, 24(9): 1000-1003. [陈家宽, 2016. 居群、物种与生物多样性 [J]. 生物多样性, 24(9):1000-1003.]
CHEN JK, WANG HQ, 1986. The application of the concept and method of population to plant taxonomy [J]. J Wuhan Bot Res, 4(4): 377-383. [陈家宽, 王徽勤, 1986. 居群(population)概念和方法在植物分类学中的应用 [J]. 武汉植物学研究, 4(4): 377-383.]
DE SMET Y, GOETGHEBEUR P, WANKE S, et al., 2012. Additional evidence for recent divergence of Chinese Epimedium (Berberidaceae) derived from AFLP, chloroplast and nuclear data supplemented with characterisation of leaflet pubescence [J]. Plant Ecol Evol, 145(1): 73-87.
GAO H, WANG H, QUAN QM, et al., 2012. Influence of three cultivation patterns on growth of Epimedium wushanense [J]. Guihaia, 32(5): 663-668. [高辉, 王辉, 权秋梅, 等, 2012. 不同栽培方式对巫山淫羊藿生长的影响 [J]. 广西植物, 32(5): 663-668.]
GUO MY, XU YQ, REN L, et al., 2018. A systematic study on DNA barcoding of medicinally important genus Epimedium L. (Berberidaceae) [J]. Gene, 9(12): 637-648.
HE SZ, 2014. The genus Epimedium of China in colour [M]. Guiyang: Guizhou Science & Technology Press: 96. [何順志, 2014. 中国淫羊藿属植物彩色图谱 [M]. 贵阳: 贵州科技出版社: 96.]
HONG DY, 2016. Opinion of raising rationality in species delimitation [J]. Biodivers Sci, 24(3): 360-361. [洪德元, 2016. 关于提高物种划分合理性的意见 [J]. 生物多样性, 24(3): 360-361.]
LIU CS, 2016. Pharmaceutical botany [M]. Beijing: China Press of Traditional Chinese Medicine: 96. [刘春生, 2016. 药用植物学 [M]. 北京: 中国中医药出版社: 96.]
LIU SX, SHI HJ, XU YQ, 2016. Morphological comparison of the Epimedium franchetii Stearn species complex based on population observation and implications for taxonomy [J]. Plant Sci J, 34(3): 325-339. [刘少雄, 石慧君, 徐艳琴, 2016. 基于居群观测的木鱼坪淫羊藿复合种形态比较和分类学启示 [J]. 植物科学学报, 34(3): 325-339.]
LIU SX, LIU LJ, HUANG XF,et al., 2017. A taxonomic revision of three Chinese spurless species of genus Epimedium L. (Berberidaceae) [J]. Phytokeys, 78: 23-36.
NOOTEBOOM HP, 1992. A point of view on the species concept [J]. Taxon, 41(2): 318-320.
QUAN QM, GAO ZM, LI YX, 2012. Comparative analysis of morphological characteristics and effective composition content of Epimedium pubescens in different habitats [J]. Guihaia, 32(3): 350-354. [权秋梅, 高泽梅, 黎云祥, 2012. 不同生境中柔毛淫羊藿形态特征及其有效成分差异分析 [J]. 广西植物, 32(3): 350-354.]
SHI X, WANG YL, LI MD, et al., 2010. Morphological notes on gynaecium and fruit of seven species in the genus Epimedium L. (Berberidaceae) [J]. J Wuhan Bot Res, 28(2): 234-238. [石旭, 王玉良, 李牡丹, 等, 2010. 淫羊藿属7种植物的雌蕊及果实形态描述的订正 [J]. 武汉植物学研究, 28(2): 234-238.]
STEARN WT, 1997. Four new Chinese species of Epimedium (Berberidaceae) [J]. Kew Bull, 52(3): 659-671.
STEARN WT, 2002. The genus Epimedium and other herbaceous Berberidaceae, including the genus Podophyllum. A botanical magazine monograph [M]. Oregon: Timber Press: 26-351.
SUN C, ZOU JL, ZHONG Y, et al., 2004. The study on the cultivation of three Epimedium species [J]. Chin J Chin Mat Med, 29(3): 274-275. [孙超, 邹剑灵, 钟雁, 等, 2004. 淫羊藿属3种植物引种栽培研究 [J]. 中国中药杂志, 29(3):274-275.]
TAN XH, WANG DQ, 2013. Pharmaceutical botany [M]. Beijing: China Press of Traditional Chinese Medicine: 127. [谈献和, 王德群, 2013. 药用植物学 [M]. 北京:中国中医药出版社:127.]
XU BS, 1998. The species problem in plant taxonoy in China [J]. Acta Phytotaxon Sin, 36(5): 470-480. [徐炳声, 1998. 中国植物分类学中的物种问题 [J]. 植物分类学报, 36(5):470-480.]
XU YQ, LIU LJ, LIU SX, et al., 2019. The taxonomic relevance of flower for Epimedium (Berberidaceae), with morphological and nonmenclatural notes for five species from China [J]. Phytokeys, 118: 33-64.
XU YQ, SHI HJ, LIU SX, et al., 2016. New records of Epimedium epsteinii (Berberidaceae) in Guangdong and Hubei provinces and morphologic description [J]. Guihaia, 36(5): 619-624. [徐艳琴, 石慧君, 刘少雄, 等, 2016. 紫距淫羊藿的地理分布新纪录及形态描述 [J]. 广西植物, 36(5):619-624.]
XU YQ, XU Y, LIU Y, et al., 2014. Progress and open problems in classical taxonomic research on Epimedium L. [J]. Chin Trad Herb Drugs, 45(4): 569-577. [徐艷琴, 许瑛, 刘勇, 等, 2014. 淫羊藿属经典分类学研究进展及存在的问题 [J]. 中草药, 45(4):569-577.]
YANG QE, 2016. Comments on species-level taxonomy of plants in China [J]. Biodivers Sci, 24(9): 1024-1030. [杨亲二, 2016. 我国植物种级水平分类学研究刍议 [J]. 生物多样性, 24(9):1024-1030. ]
YING JS, 1975. On the chinese species of Epimedium L. [J]. Acta Phytotaxon Sin, 13(2): 49-56. [应俊生, 1975. 中国淫羊藿属植物的种类和分布 [J]. 植物分类学报, 13(2):49-56.]
YING JS, 2001. Epimedium L. [M]//YING JS, CHEN TC. Flora Reipublicae Popularis Sinicae. Beijing: Science Press, 29: 262-293. [应俊生, 2001. 淫羊藿属 [M]// 应俊生, 陈德昭. 中国植物志. 北京:科学出版社, 29:262-293.]
YING JS, BOUFFORD DE, BRACH AR, 2011. Berberidaceae [M]//WU ZY, RAVEN PH, HONG DY. Flora of China. Beijing: Science Press; St. Louis: Missouri Botanical Garden Press, 19: 787-799.
YOUSAF Z, HU WM, ZHANG YJ,et al., 2015. Systematic validation of medicinally important genus Epimedium species based on microsatellite markers [J]. Pak J Bot, 44(2): 477-484.
ZHANG ML, UHIK CH, KADREIT JW, 2007. Phylogeny and biogeography of Epimedium/Vancouveri (Berberidaceae): Western north American-East Asian disjunctions, the origin of European mountain plant taxa, and east asian species diversity [J]. Syst Bot, 32: 81-92.
ZHANG YJ, YANG LL, CHEN JJ, et al., 2014. Taxonomic and phylogenetic analysis of Epimedium L. based on amplified fragment length polymorphisms [J]. Sci Hortic, 170: 284-292.
ZHANG YJ, DANG HS, LI SY, et al., 2015. Five new synonyms in Epimedium (Berberidaceae) from China [J]. Phytokeys, 49: 1-12.
(责任编辑 蒋巧媛)
