Chemerin对奶牛乳腺上皮细胞炎症和内质网应激及自噬的影响
2020-05-18胡辩红韩萍萍唐宇杰崔萍萍韩兆玉李惠侠
胡辩红,韩萍萍,唐宇杰,崔萍萍,韩兆玉,李惠侠
(南京农业大学 动物科技学院,江苏 南京 210095)
奶牛乳腺组织的健康与养殖者的经济效益和乳品质有着直接关系。尽管广泛实施控制措施,奶牛乳房炎仍然是一个棘手的问题。乳房炎的发生通常伴随着促炎性细胞因子的分泌,但是炎性因子的过量产生,特别是 IL-1β 和 TNF-α 的过度分泌,会促使一系列继发性炎症介质的产生并导致局部/全身病理学疾病[1]。活性氧(ROS)作为一种促炎性介质,是导致各种炎症性疾病继发性的关键原因。当其积累超出机体的抗氧化能力时,会对宿主细胞和吞噬细胞本身造成损害,主要通过降低细胞外基质与乳腺上皮细胞的黏附性,引起乳腺上皮细胞死亡、脱落及泌乳功能的损伤[2]。内质网应激是对细胞内外应激具有处置应答能力的另一亚细胞行为,也是细胞对内质网内大量未折叠或错误折叠蛋白积累的一种适应性应答方式。内质网应激是炎症的有效组成部分,不仅参与炎症反应,同时也是引起炎症的重要原因[3]。内质网应激同时也可促进Ca2+释放激活多种激酶参与自噬信号转导[4]。自噬是细胞内一种依赖溶酶体的降解途径,可发挥改善内质网应激的作用[5]。越来越多的证据表明自噬与各种生理病理状况有关。自噬异常激活将会导致过度的自我消化和细胞死亡,成为致病机制之一[6]。因此自噬成为疾病研究的新靶标。
Chemerin也称为视黄酸受体反应蛋白2(RARRES2),是2007年新确认的脂肪因子,在白色脂肪组织和肝脏中高度表达。现已证实,Chemerin在脂肪脂解与分化、糖代谢、生殖调控、肿瘤等方面都发挥一定的调控作用[7]。而在免疫功能方面,许多试验证据支持Chemerin/ CMKLR1的促炎作用,但也有研究表明该信号传导途径具有抗炎功能[8],这主要取决于Chemerin的酶切位点。Chemerin在奶牛乳腺中表达也被证实,并上调与脂肪酸合成、葡萄糖摄取和酪蛋白合成相关的基因[9]。但是,关于Chemerin对奶牛乳腺炎和内质网应激及自噬的研究目前还比较少。本试验拟通过Chemerin处理奶牛乳腺上皮细胞以分析炎症反应,及这个过程当中内质网应激与自噬的变化,为探究炎症与内质网应激及自噬之间的关系奠定基础,进一步完善乳腺疾病的发病机制。
1 材料与方法
1.1 主要试剂重组小鼠蛋白Chemerin(2325-CM-025)购自R&D公司并参考文献[10]所描述的方法溶解;IL-6(GB11117)购自北京索莱宝生物公司;IL-1β(A1112)购自ABclonal公司;LC3B (NB100-2220SS) 购自 NOVUS生物公司;Bax(50599-2-1g)、Bcl-2(12789-1-AP)、p62/SQSTM1 (18420-1-AP)、GRP78(11587-1-AP)和GAPDH(10494-1-AP)均购自Proteintech公司;Caspase-3(bs-0081R)抗体购自Bioss公司;CHOP(2895)抗体购自Cell Signaling Technology公司;Cytokeratin 18(ab52459)购自Abcam公司;荧光二抗Goat Anti-Rabbit IgG(FMS-Rbaf64701)购自福麦斯生物公司;Hoechst33342染色液(KGA212-50)购自凯基生物公司;ROS检测试剂盒购自南京建成生物研究所。
1.2 细胞培养与冻存本试验所采用的奶牛乳腺上皮细胞系哈佛大学孙友平博士所赠送。将处于对数生长期且融合率达到90%的细胞用PBS冲洗后消化,待大部分细胞变圆且有少部分从培养瓶上脱落悬浮时,加入1 mL完全培养基终止消化,充分晃匀后3 000 r/min离心4 min,去上层液并加入DMSO配制的冻存液,分装封口后4℃ 30 min→-20℃ 2 h→-80℃ 过夜,最后在液氮中长期保存。从液氮罐中取出冻存的奶牛乳腺上皮细胞并迅速置于37℃水浴锅中,轻轻摇晃使其快速融化并转移到超净台中,加入提前预热的完全培养基离心洗涤(3 000 r/min,4 min)2次,重悬后接种在T25培养瓶中,于37℃、5% CO2、100% 湿度的二氧化碳培养箱内培养。12 h待细胞完全贴壁后轻轻更换培养基以去除死细胞,当细胞生长汇合至90%时进行消化传代培养,在此期间,每2~3 d更换新鲜培养基。
1.3 试验设计和处理为了分析Chemerin对奶牛乳腺上皮细胞炎症、内质网应激及自噬的影响,用重组蛋白Chemerin(0.1 mg/L)体外处理细胞24 h,分为对照组和Chemerin组,处理结束后分别收集细胞并进行相应指标的检测。试验浓度和时间的选择主要依据于我们团队先前的研究,证实0.1 mg/L Chemerin处理24 h具有最低的细胞毒性(数据未列出)。
1.4 Western blot分析按照试验方案收集各处理组细胞,每瓶细胞加400 μL含PMSF的裂解液,于冰上裂解30 min后提取细胞总蛋白。用BCA试剂盒测定蛋白浓度并加入上样缓冲液高温变性。通过12%SDS-聚丙烯酰胺凝胶电泳分离等量的蛋白质样品,并电转移到PVDF膜上以形成蛋白印迹,然后将膜与5%脱脂牛奶室温封闭1 h,加入一抗后4℃过夜,彻底洗涤后,在室温下与HRP偶联的二抗孵育1 h,TBST洗3次(10 min/次),最后用ECL显影液避光显色拍照,并通过Image J软件对蛋白质条带进行灰度分析。
1.5 细胞内ROS的检测将各组细胞制成细胞爬片,处理后用提前预热的PBS轻轻洗涤3次,根据ROS检测试剂盒说明书进行如下操作,细胞与10 μmol/L DCFH-DA 在37℃条件下避光孵育1 h,然后用PBS洗涤3次以去除未进入细胞而残留的DCFH-DA,用Hoechst33342染液对细胞核复染15 min,PBS洗3次/(3 min/次),最后在共聚焦显微镜下随机选择5个高倍视野拍摄并用图像分析软件计算出绿色荧光强度的平均值。
1.6 免疫荧光检测将细胞均匀的接种到24孔爬片上,待细胞融合度达到80%时进行相应处理,然后按照免疫荧光步骤进行如下操作,吸出培养基,用PBS彻底洗涤后加入4%多聚甲醛固定30 min,PBS洗3次/(5 min/次),随后用5% Triton-x100膜透化20 min,PBS清洗后用5% BSA孵育1 h以封闭非特异性结合位点,并在4℃条件下分别与按1∶500稀释的一抗角蛋白18和p62孵育过夜,PBS洗3次/(5 min/次),然后与荧光二抗(1∶500)在室温下避光孵育1 h,用PBS清洗后加入Hoechst33342染核15 min,PBS洗3次后用甘油封片,最后在共聚焦显微镜下观察。
2 结果
2.1 奶牛乳腺上皮细胞的体外培养和鉴定细胞角蛋白是属于中间丝蛋白家族的一员,其中角蛋白18是乳腺上皮细胞骨架的主要组成成分。在本试验中,使用角蛋白18的免疫荧光染色鉴定奶牛乳腺上皮细胞。 如图1所示,细胞形态清晰,界限分明,呈“铺路石”状生长。此外,细胞角蛋白18的阳性染色率大于95%,证明我们培养的细胞中奶牛乳腺上皮细胞占90%以上,可作为细胞模型用于后续试验。
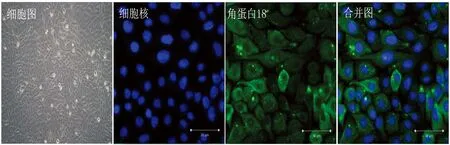
图1 奶牛乳腺上皮细胞体外培养及角蛋白18免疫荧光鉴定细胞(取生长状态良好的奶牛乳腺上皮细胞,以1×106 /mL密度接种于24孔板爬片,角蛋白18荧光染色后利用共聚焦显微镜观察,试验独立重复3次,每张细胞爬片随机取5个视野拍摄保存)
2.2 Chemerin对奶牛乳腺上皮细胞促炎性炎症的影响将奶牛乳腺上皮细胞与Chemerin孵育24 h后在蛋白质水平检测炎症相关因子的表达结果见图2A,B。与对照组相比, Chemerin组IL-1β和IL-6的蛋白表达显著升高(P<0.01);同时,通过免疫荧光染色检测发现,ROS的绿色荧光在Chemerin处理后明显增强(P<0.01)(图2C,D),表明Chemerin通过上调促炎性因子和促炎性介质的表达诱发奶牛乳腺上皮细胞的炎症反应。
2.3 Chemerin对奶牛乳腺上皮细胞内质网应激的影响内外环境中的多种刺激都会引起内质网中未折叠蛋白积聚导致应激反应。为进一步分析Chemerin对奶牛乳腺上皮细胞的作用,本试验采用Western blot检测内质网的分子伴侣GRP78和内质网应激标志基因CHOP的蛋白表达,结果见图3A,B。Chemerin的处理使GRP78和CHOP的蛋白表达显著升高(P<0.05),证实Chemerin在奶牛乳腺上皮细胞中激活内质网应激反应。
2.4 Chemerin对奶牛乳腺上皮细胞自噬的影响由图4A,B可知,Chemerin处理显著增加LC3-Ⅱ并降低p62的蛋白表达(P<0.01);进一步通过免疫荧光染色观察到p62表达明显减弱(P<0.001)(图4C,D)。这些结果表明,在奶牛乳腺上皮细胞中Chemerin通过调控自噬标志基因的表达激活自噬反应。

图2 Chemerin对奶牛乳腺上皮细胞细胞炎症的影响(将奶牛乳腺上皮细胞以1×106 /mL密度接种于6孔板和24孔板爬片中,待细胞融合度达到80%时在培养基中添加0.1 mg/L的Chemerin处理24 h,分为对照组和Chemerin组)1.A为通过蛋白免疫印迹法检测IL-1β和IL-6的表达; 2.B为通过Image J软件量化炎症相关蛋白的水平;3.C为用DCFH-DA染色后,在共聚焦显微镜下观察细胞内ROS表达,每张爬片随机选取5个视野拍摄,绿色荧光代表ROS,蓝色荧光代表细胞核,比例尺:50 μm;4.D为ROS阳性细胞相对于体内细胞核总数的定量分析;5.所有的数据表示为平均值±标准误每个处理一式3份进行;6.与对照组相比,*P< 0.05,**示P<0.01。下同
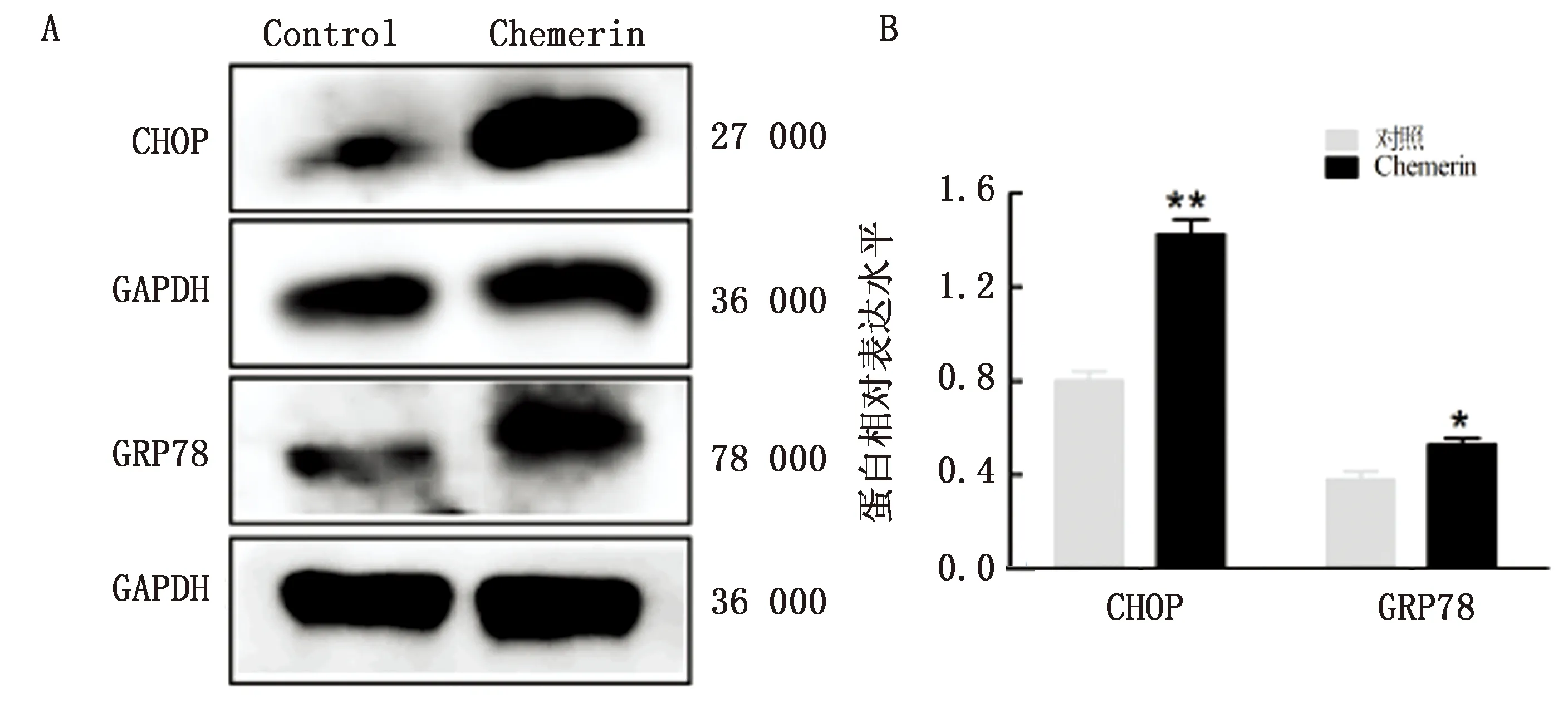
图3 Chemerin在奶牛乳腺上皮细胞中对内质网应激关键蛋白CHOP与GRP78表达的影响(体外调整奶牛乳腺上皮细胞密度为1×106 /mL并接种于6孔板中,将汇合度达到80%的细胞与0.1 mg/L的重组蛋白Chemerin共同培养24 h,分为对照组和Chemerin组)1.A为通过蛋白免疫印迹法检测CHOP和GRP78的表达;2.B为通过Image J软件量化内质网应激相关蛋白的水平

图4 Chemerin对奶牛乳腺上皮细胞自噬的影响(取生长状态良好的奶牛乳腺上皮细胞,以1×106 /mL密度接种于6孔板和24孔板爬片中,待细胞融合度达到80%时在培养基中添加0.1 mg/L的Chemerin孵育24 h,分为对照组和Chemerin组)1.A为LC3和p62蛋白表达的电泳图;2.B为LC3-Ⅱ和p62蛋白的相对表达量;3.C为免疫荧光技术检测p62蛋白的表达,每张细胞爬片随机取5个视野拍摄保存,红色荧光代表p62蛋白,蓝色荧光代表细胞核,比例尺:50 μm;4.D为通过Image J软件分析p62的平均荧光强度。
3 讨论
Chemerin是一种分泌蛋白,病理状态下,其大量存在于人体不同的炎性体液和组织中,通过Chemerin/CMKLR1轴实现对炎症反应的调节[11]。本试验结果表明,Chemerin及其受体CMKLR1在患有临床性乳腺炎的奶牛乳腺组织中高表达(数据未列出),与文献[11]的研究结果高度一致,提示Chemerin/CMKLR1轴可能参与奶牛乳腺炎的发生与发展。
在炎症或组织损伤部位Chemerin具有募集和激活免疫细胞的特点,其含量升高通常伴随着炎性因子TNF-a、IL-1β、IL-6 和C反应蛋白的升高[12]。IL-1β在乳腺感染的早期能大量分泌促进免疫细胞活化,发挥招募聚集第二级细胞因子的能力,并与内源性炎症因子IL-6协同作用以形成级联效应导致组织损伤。乳腺炎发生过程中TNF-a与IL-1β不仅发挥促进中性粒细胞迁徙、黏附聚集细胞因子的功能,同时还促进细胞内ROS的释放[13]。在一些炎症相关的肺部疾病中,活化的炎性细胞被认为积聚在下呼吸道中并释放大量的ROS,继而引发相关疾病的发展[14]。Chemerin可通过ChemR23依赖性方式刺激ROS产生诱导炎症[15]。在本试验中,Chemerin的处理能够显著上调炎性因子IL-1β和IL-6的蛋白表达,同时促进ROS的积累,这表明Chemerin通过促进炎性因子IL-1β和IL-6的分泌以及炎性介质ROS的产生参与奶牛乳腺上皮细胞炎症的发生发展。
内质网是细胞内重要的多功能细胞器之一,内质网功能失常诱导炎症反应的能力被认为在疾病发病机理中发挥重要作用。最近的证据表明,内质网应激与促炎因子的激活有关[3]。适度的内质网应激能缓解由TNF-α诱导的NF-κB激活,从而抑制炎性细胞黏附因子的上调[16]。Chemerin在体内分泌的减少能抑制氧化应激和内质网应激反应[17]。在肥胖诱导的慢性炎症中,Chemerin水平与内质网应激相关蛋白GRP78呈正相关[18]。GRP78主要负责感应内质网应激信号,从而调节下游的CHOP基因引发一系列的细胞反应[19]。在本试验中,Chemerin同样上调GRP78和CHOP的蛋白表达激活乳腺上皮细胞内质网应激。近来发现内质网应激是自噬的重要调控通路。自噬是依赖于溶酶体的降解过程,在维持内环境稳态上发挥重要的重要。内质网应激可通过调节LC3和ATG5的转录从而影响自噬体的形成[20]。内质网应激负调节mTOR途径以增强自噬[4]。此外,内质网应激导致钙的释放和随后AMPK的激活,AMPK抑制ROS从而促进自噬[21]。在奶牛乳腺上皮细胞中我们发现Chemerin显著诱导了LC3-Ⅱ蛋白的表达,降低了p62蛋白和免疫荧光的表达。LC3-Ⅱ主要并附着于自噬体膜上,而p62是作为自噬底物存在,高LC-Ⅱ低p62的模式是自噬发生的黄金指标。因此,Chemerin在奶牛乳腺上皮细胞中可诱导自噬发生。而自噬又和先天免疫系统相互调节,通过消除内源性炎性体激动剂以及对与免疫信号传导相关的介质分泌的影响来调节炎症[22]。但异常积累的自噬导致急性炎症性疾病和细胞死亡[23]。因此,在奶牛乳腺上皮细胞中,Chemerin激活的内质网应激是否介导自噬调控奶牛乳腺上皮细胞炎症,目前尚不明确,还需进一步研究。
综上所述,细胞因子Chemerin能促进奶牛乳腺上皮细胞炎症的发生,同时激活内质网应激与自噬反应,但具体的机制及内质网应激-自噬反应和炎症之间的关系还需进一步深入研究,这将为临床上的靶向奶牛乳腺炎药物研发提供部分理论数据。
