茄子灰霉病原菌遗传多样性分析
2020-04-28冯宝珍李培谦
冯宝珍,李培谦
(运城学院 生命科学系,山西 运城 044000)
茄子(SolanummelongenaL)是我国重要的蔬菜作物之一,也是设施蔬菜栽培的重要种类。随着栽培模式的改进,设施蔬菜成为现代农业的重要组成部分。但由于保护地空气湿度大、种植密度大,常导致茄子灰霉病发生严重,已成为制约茄子质量和产量的重要因素[1]。
灰霉病主要由灰葡萄孢(Botrytiscinerea)引起的一类重要病害。该病原菌在世界范围均有分布,其寄主范围广泛,能够侵染大约1 000种植物[2]。灰霉病在植物生长过程及采后贮藏中都能发病,造成水果、蔬菜和观赏植物组织坏死、腐烂,造成严重经济损失,被列为世界第二严重的真菌病害[3]。
目前国内外已开始关注灰葡萄孢(B.cinerea)的种内多样性研究。利用RAPD[4]、RFLP[5]、AFLP[6], MP-PCR[7]、微卫星标记[8]以及DNA序列[9,10]均能证实灰霉病菌有丰富的多样性。Ma[7]利用MP-PCR技术研究了美国佛罗里达州不同寄主上灰霉病菌遗传结构,认为不同寄主的灰霉病菌分化程度高,菌群重组方法不一致。Leroch等[11]通过多基因测序证实德国草莓种植地灰霉菌为一个新的亚群,称之为Botrytis group S,与B.cinerea和B.fabae亲缘关系很近,而且该亚群在德国葡萄园未发现。刘双清等[12]对湖南省草莓灰霉病菌群体遗传结构、分化及变异规律进行分析,认为湖南省各地草莓灰霉菌群体遗传多样性丰富,且遗传差异主要来自群体内部,与地理位置关系小。张静[13]利用形态学和分子系统学对湖北省作物灰霉病菌研究,病原物鉴定出14个种,主要病原为B.cinerea。张佳[14]对我国草莓主产区灰霉病多样性进行研究,发现灰霉菌株间在培养性状、交配型、转座子类型、抗性方面均存在明显差异。
研究灰霉菌群内多样性对于明确田间优势种群及灰霉病防治具有重要意义。本研究对运城保护地茄子灰霉病菌表型多样性和遗传多样性进行分析,以明确茄子灰霉病原菌优势种群特征,为灰霉病防治提供科学依据。
1 材料与方法
1.1 供试菌株分离及鉴定
供试15个茄子灰霉菌株于2017-2018年冬春季节采于山西运城盐湖区茄子种植保护地。采用单孢分离法获得菌株,在PDA斜面培养5~7 d,置于4 ℃冰箱保存。
挑取PDA平板上培养3 d的菌丝于铺有玻璃纸的新的PDA平板中央,放入20 ℃恒温箱中继续培养3 d,收集菌丝。收集的菌丝在液氮中研磨,按照真菌基因组DNA提取试剂盒(北京艾莱德生物科技有限公司)说明书提取基因组DNA。
将提取基因组DNA用于真菌ITS序列和灰葡萄孢特异序列PCR扩增,以鉴定灰葡萄孢。PCR扩增体系为:模板 1 μL,ddH2O 9.5 μL,上游引物1 μL,下游引物1 μL,2×Taq PCR Master Mix 12.5 μL,共25 μL。ITS序列扩增引物为ITS1(5’-TCCGTAGGTGAACCTGCGG-3’)和ITS4(5’-TCCTCCGCTTATTGATATGC-3’),灰葡萄孢特异序列扩增引物BCF(5’-CAGGAAACACTTTTGGGGATA-3’)和BCR(5’-GAGGGACAAGAAAATCGACTAA-3’)[15]。
反应条件:预变性94 ℃,5 min;94 ℃,30 s,55 ℃,30 s,72 ℃,1 min,共35个循环;而后72 ℃下延伸10 min;最后降为16 ℃稳定。
1.2 生物表型分析
菌株在PDA平板培养3 d后,用打孔器取最边缘的菌饼转移到新PDA平板中央,置于20 ℃恒温箱中培养30 d。分别在24 h和48 h测量菌落直径,并计算菌丝生长速率。每个菌株做3个重复。
计算公式:菌丝生长速率/cm·d-1=(48 h菌落直径-24 h菌落直径)/2。
15 d后观察菌株菌核形成情况,并挑取菌丝制备玻片,在光学显微镜下观察有无分生孢子及形态。
1.3 RAPD分子标记
本试验使用18条随机引物对供试菌株DNA进行遗传多样性初筛,筛选能产生3条带以上、重复性好且清晰度高的引物。所用引物由上海生物工程股份有限公司合成(表1)。
将RAPD扩增体系及反应条件进行优化。确定的反应体系为25 μL,其中ddH2O 9.5 μL,Taq Master Mix 12.5 μL,引物2 μL以及模板DNA 50 ng。扩增的反应条件为:预变性94 ℃,4 min;94 ℃,45 s,36 ℃,45 s,72 ℃,1 min,45个循环;72 ℃,10 min;最后降为4 ℃稳定。扩增结束后,将产物用1.5%的琼脂糖凝胶在1%的TAE缓冲液中电泳1 h,电压120 V[16]。加入golden-view染色,在凝胶成像系统中成像。

表1 RAPD扩增引物及序列
1.4 数据分析
根据扩增结果读取多态性条带,记录多态性的DNA条带数,并计算出多态性比率。将所有多态性DNA条带中有DNA条带的记为“1”,没有DNA条带的记为“0”,构建矩阵。利用POPGEN软件(version 1.32)计算菌株的遗传多样性参数,主要包括观测等位基因数(Na)、有效等位基因数(Ne)、基因多样性指数(H)和Shannon信息指数(I)[17]。使用UPGMA方法进行聚类分析,并通过Tree plot生成系统树[18]。
2 结果与分析
2.1 灰霉病菌株分离及鉴定
共分离纯化了15个茄子灰霉病菌株,分别命名为QZBC1-15。将这15个菌株分别提取基因组DNA后,进行ITS 和灰葡萄孢特异引物扩增。15个参试菌株均能扩增出大小一致的单一条带(图1),其中 ITS扩增条带约为550 bp,BCF/R引物扩增条带大小在300 bp左右,均与预期结果一致,说明分离纯化的灰霉菌株均为灰葡萄孢(B.cinerea)。

上图为ITS扩增结果,目的条带在550 bp左右;下图为灰葡萄孢特异引物BCF/R扩增结果,目的条带大约300 bp。M为DNA分子标记;1~15为供试菌株;CK为样性对照;- 为阴性对照
2.2 生物表型分析
将分离纯化的15个灰葡萄孢菌株在PDA平板上于20 ℃恒温箱中培养30 d,前48 h测量生长速率(图2)。15个菌株生长速率存在明显差异,其中QZBC4生长最慢,为0.5 cm·d-1;其次是菌株QZBC2和QZBC6生长较慢;而菌株QZBC1、8及13生长较快,达到1.4 cm·d-1。

图2 茄子灰葡萄孢菌株生长速率
参照前人划分菌落标准[14],将15个菌株在PDA上20 ℃培养30 d,其菌落主要存在4种形态(图3)。菌丝型,不形成菌核,气生菌丝茂盛,产孢量极少,菌株QZBC3、8、6、7、9、14和15属于这种类型,如图3-a所示。孢子型,每皿菌核少于10粒,其生菌丝茂盛,产孢量极大。菌株QZBC10、11、12、13属于孢子型,如图3-b所示。细小菌核型,菌核极小,数目极多,分散分布,产孢较多,如图3-c 所示,菌株QZBC1、2、5属于此类型。大菌核型,菌核鼠粪状且数目多,呈圆形规则分布于培养皿中,菌丝和产孢量较少,QZBC4属于此类型,如图3-d所示。产菌核的菌株,继续培养可在菌核上萌发产生菌丝和孢子。

图3 菌落菌核形态及分生孢子产量情况
2.3 RAPD分析
从18条随机引物中筛选出4条引物,这4条随机引物的扩增条带均在3条以上,具有很高的重复性,且清晰度高(表2)。将筛选出的4条随机引物分别对15个茄子灰霉病菌菌株基因组DNA进行PCR扩增,扩增结果显示片段长度范围在175~2 000 bp,产生的总条带数为30条,其中扩增出的多态性条带数为22条,多态性比率为73.3%。4条随机引物分别扩增的总的条带数中,总条带数最多的引物为S7,扩增出11条,最少的引物为S8,扩增出5条,平均每条随机引物扩增的总的条带数为7.5条。
2.4 遗传多样性分析
将整理好的矩阵图使用POPGEN软件计算遗传多样性参数,得出观测等位基因数(Na)为2.0000,有效等位基因数(Ne)的最高值为1.9912,最低值为1.0000,平均值为1.3708,基因多样性指数(H)最高值为0.4978,最低值为0.0000,平均值为0.2352,Shannon指数(I)最高值为0.6909,最低值为0.0000,平均值为0.3709,其中在S7、S8、OPF-01和OPF-04这4条引物中,Ne值、H值、I值最高的引物是S7和S8,值最低的引物是OPF-01(表3),遗传多样性较为丰富。
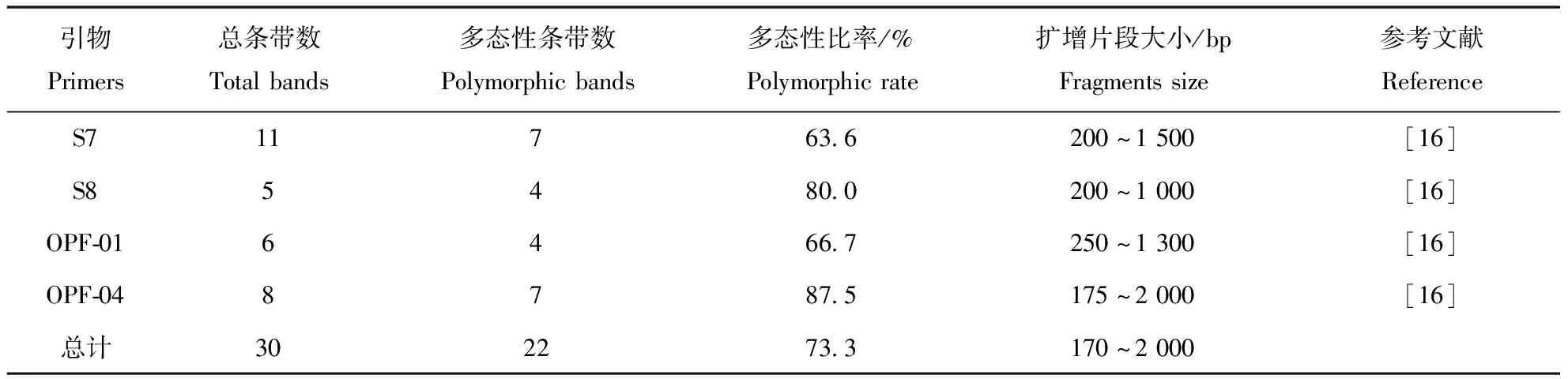
表2 筛选出的4条RAPD引物及扩增结果

表3 4条RAPD引物的遗传多样性参数
由图4可以看出,在相似数0.76处,15个茄子灰葡萄孢菌株被划成4个类群。其中Ⅰ类群有3个,为菌株QZBC1、2和5。该类型菌株均能产生细小菌核。Ⅱ类群有5个菌株有划为2个类群,即包括孢子型菌株QZBC10、11、12和13,还包括能产生菌核的菌株 QZBC4。Ⅲ类群有3个菌株,为QZBC3、8和9;Ⅳ类群有4个菌株,为QZBC6、7、14和15,这2个类群的菌株均为菌丝型。

图4 15个茄子灰葡萄孢菌株聚类分析图
3 讨论与结论
本研究对15株茄子灰霉病原菌遗传多样性进行分析,首先利用ITS引物和灰葡萄孢特异引物确定了这些菌株均为B.cinerea,然后对参试菌株的生长速率、菌落形态、菌核形成分布和产孢情况进行分析,并利用RAPD分子标记进一步分析了遗传多样性。15个茄子灰葡萄孢菌株表现出明显的遗传多样性。
经过ITS引物和灰葡萄孢特异引物扩增,明确了茄子灰霉病菌均为B.cinerea,说明运城茄子灰霉病菌优势种为B.cinerea。其他学者研究发现,作物灰霉病病原除了B.cinerea外,还有B.sinoviticola和B.pelargonii等[19,20]。生长速率测定显示15个菌株的生长速率存在明显差异,这与前人研究结果一致[19]。张佳[14]对我国草莓灰霉病研究中发现,不同灰葡萄孢菌株的菌落和菌核各异,并根据产孢量、菌丝疏密情况和菌核数量分布,将菌落分为3大类型即菌丝型、孢子型和菌核型,将菌核型又细分为5种类型。本研究中15个菌株的菌落类型均在上述范围之中,以菌丝型为主有7个菌株,孢子型和菌核型各有4个,其中大菌核型有1个,小菌核3个。后续还需要对更多灰葡萄孢菌株进行分析以明确茄子灰霉病菌菌落特征的多样性。
张蕊等[16]利用RAPD法分析了Amphobotrysricihi的种内多样性,从50条引物里筛选出8条引物,试验结果显示种内菌株存在明显的遗传多样性。Kandan等[21]采用RAPD标记从50条引物里筛选出10条引物分析高粱靶斑病菌多样性。本研究利用RAPD法从18条随机引物里,筛选出4条引物,可用于后续茄子灰霉病菌多样性分析。本研究中15个菌株划分为4个类群,并且类群划分与菌落形态有相关性,说明表型多样性与遗传多态性相统一,同时也证明了同一寄主的灰葡萄菌株间存在明显的遗传分化。范咏梅等[18]利用RAPD法对新疆的灰葡萄多态性进行分析,12个菌株被划为4个类群,证明了灰霉菌株间存在遗传分化,与本研究结果一致。
综上所述,运城茄子灰霉病菌优势种为B.cinerea,种内存在形态多样性。后续研究将对灰霉菌株的毒力和抗性差异进行分析。研究结果可为灰霉病的防控提供理论依据。
