8种易裂果实中果胶酶生物信息学分析
2020-04-28刘亚令解潇冬耿雅萍杨俊强张鹏飞
刘亚令,解潇冬,耿雅萍,杨俊强,张鹏飞
(1.山西农业大学 生命科学学院,山西 太谷 030801;2.山西农业大学 园艺学院,山西 太谷 030801;3.山西省农业科学院 园艺研究所,山西 太原 030031)
裂果是一种多发生在果实膨大期的果树常见病,主要表现为果实崩裂与开裂2种现象。樱桃、西瓜、番茄、枣等多种水果常因裂果病的发生造成不同程度的减产而致使经济损失严重。比较常见的枣一般年份有30%裂果率,严重情况下能达到60%的裂果率甚至绝收[1];樱桃每年会有30%裂果率,严重时也会达到80%[2];葡萄、甜橘等水果每年也会有50%的裂果产生,易裂番茄裂果率几乎达到100%[3]。裂果病的发生因素是多方面的,目前有关于裂果病的研究主要集中在气候、土壤、水分、矿质等外界条件,果实结构、遗传因素、内源激素、种质资源等内在因素2个方面展开[4~6]。在裂果病防御中,除了改善选种育种和农耕等措施之外,喷施CaCl2是一种广泛运用于多种果树裂果的有效防治方法[7~9],主要是由于Ca2+与植物细胞壁的主要成分之一的果胶结合形成果胶钙盐,从而增加细胞壁的稳定性以及维持细胞间作用力,以达到防止裂果的目的。
果胶酶(Pectinase)作为一种分解果胶的直接功能酶,其含量和活性直接影响到细胞壁的稳定性,进而影响裂果的发生。郭红彦等发现钙处理壶瓶枣可以使果实中果胶及纤维素含量增加,细胞壁代谢相关酶——果胶酶和纤维素酶活性减低[10];李欢等在骏枣裂果研究中发现,相对于不裂果的骏枣,裂果骏枣中果胶酶和纤维素酶的相关基因表达明显上调[11];张川在裂果番茄细胞壁代谢研究中得出,随着番茄果实的成熟,果胶纤维素等物质的含量下降,同时果胶酶和纤维素酶等含量及活性增加[3];除此之外,在苹果、枇杷等多种水果中也发现果胶酶与裂果发生存在一定相关性[12,13],这些结果为裂果机制的研究以及裂果病的防治奠定了一定的基础。
果胶酶一般可以分为3类:果胶酯酶(Pectinesterase,PE)、多聚半乳糖醛酸酶(Polygalacturonase,PG)和果胶裂解酶(Pectatelyase,PL),其中前2种在植物和微生物中都有存在,第三种仅存在于微生物中。本文主要利用生物信息学软件和在线工具,对拟南芥(Arabidopsisthaliana(L.)Heynh.)、番茄(LycopersiconesculentumMill.)、苹果(MalusdomesticaL.)、白梨(PyrusbretschneideriRehd.)、葡萄(VitisviniferaL.)、枣(ZiziphusjujubaMill.)、甜樱桃(Cerasusavium(L.)Moench.)、甜橙(Citrussinensis(L.)Osbeck)、桃(AmygdaluspersicaL.)的果胶酯酶和多聚半乳糖醛酸酶进行分析。通过对2种酶的核酸和蛋白质序列的比对和预测,了解其理化性质、结构及同源关系等特点,为进一步研究果胶酶生理调控机制、裂果发生机制及其分子水平上的调控提供理论基础。
1 材料与方法
1.1 材料
本试验所选择的材料为易裂果的植物,包括番茄、苹果、白梨、葡萄、枣、甜樱桃、甜橙、桃等,拟南芥作为序列比对的模式植物,生物信息学分析数据所用的DNA、mRNA以及蛋白质序列材料均来源于NCBI(https://www.ncbi.nlm.nih.gov/),其基因ID、mRNA及蛋白质序列登录号如表1所示。
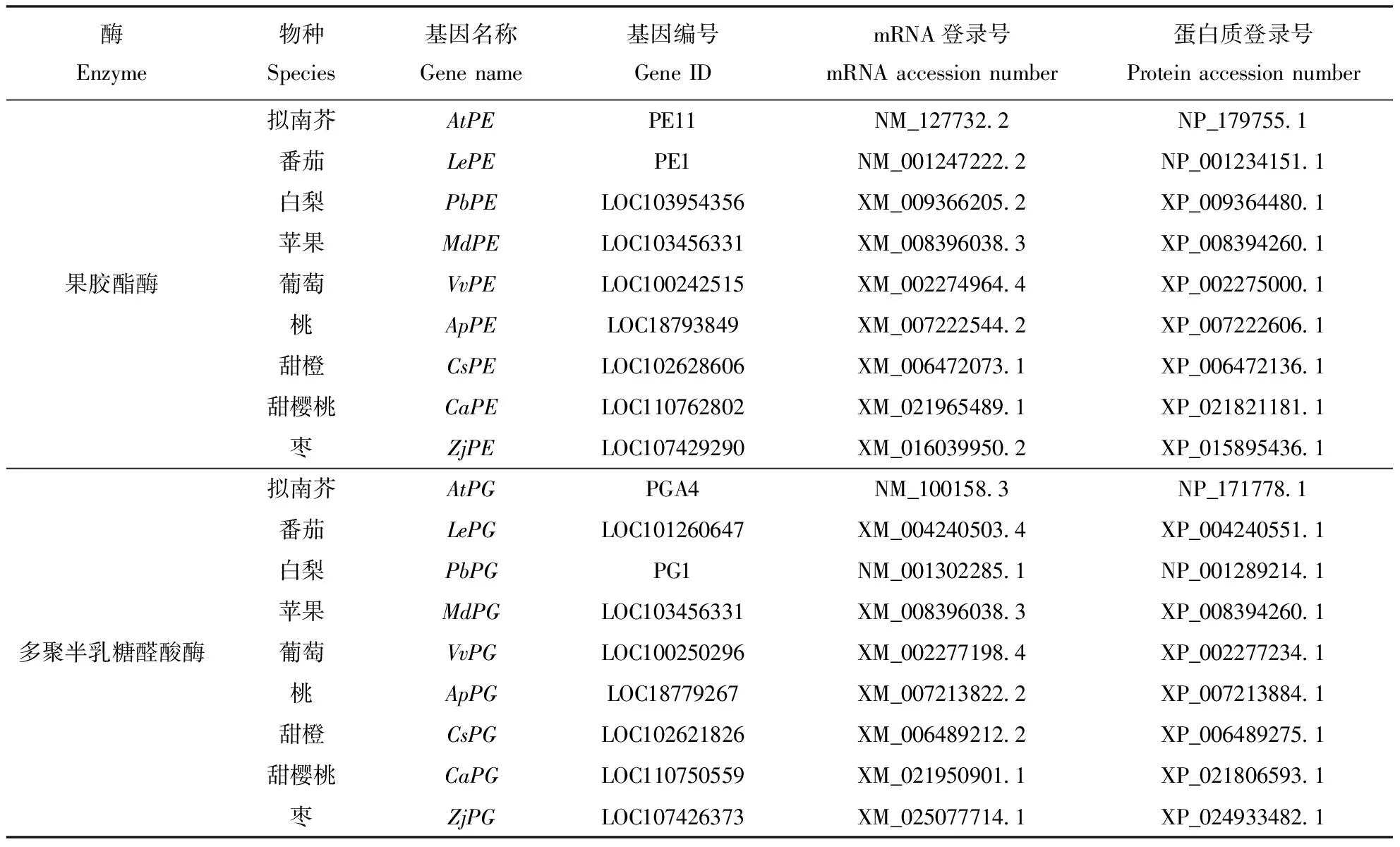
表1 材料信息
1.2 蛋白质理化性质及亲水性分析
利用在线工具ProtParam[14](http://web.expasy.org/protparam/)对蛋白质理化性质进行分析,利用ProtScale[15](https://web.expasy.org/protscale/)对蛋白质亲水性和疏水性进行分析。
1.3 蛋白质跨膜结构及亚细胞定位分析
利用德泰生物科技有限公司平台上在线工具(http://www.detaibio.com/tools/ transmembrane.html)对蛋白质跨膜结构进行分析,利用PSORT Prediction[15](http:// psort1.hgc.jp /form.html)对蛋白质亚细胞定位分析。
1.4 蛋白质修饰位点预测
蛋白质糖基化位点在NetOGlyc平台进行预测,其中O-糖基化位点在NetOGlyc 4.0(http://www.cbs.dtu.dk/services/NetOGlyc/),N-糖基化位点在NetNGlyc 1.0(http://www.cbs.dtu.dk/services/NetNGlyc/)进行预测,磷酸化位点在NetPhos 3.1进行预测(http://www.cbs.dtu.dk/services/NetPhos/)[14]。
1.5 蛋白质结构预测分析
蛋白质二级结构利用PRABI-GERLAND在线平台(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_ automat.pl?page=npsa_sopma.html)进行预测,蛋白质三级结构利用RaptorX(http://raptorx.uchicago.edu/ StructPredV2/predict/)进行预测[15]。
1.6 进化树的构建及保守结构域预测分析
先利用ClustalX对蛋白质序列进行比对,再利用软件DNAman对所得结果作序列同源性比对图;利用MEGA7软件构建系统发育进化树。利用MEME[16]在线平台(http://meme-suite.org/tools/meme)对蛋白质的保守结构域进行分析。
2 结果与分析
2.1 蛋白质理化性质及亲水性分析
蛋白质的理化性质结果见表2。蛋白质的理化性质在蛋白质功能行使及应用等方面有重要影响。蛋白质的等电点(Point Isoelectric,pI)是蛋白质在溶液状态下,溶液中正负离子所带电荷达到“中和”时的溶液pH值,在蛋白质分离提纯中有重要作用。不稳定系数是判断蛋白质稳定性的一种标准,当不稳定系数小于40,则蛋白质稳定,大于40则蛋白质处于不稳定状态。以拟南芥为对照,分析8种易裂果水果的果胶酯酶和多聚半乳糖醛酸酶:在所有的果胶酯酶中,拟南芥果胶酯酶最小由352个氨基酸组成,相对分子质量为39 057.97 Da,其他水果的果胶酯酶大小相差不大,氨基酸长度范围为536~571 aa,相对分子质量大小为59 007.66~62 856.40 Da。其中番茄、甜橙和枣的果胶酯酶等电点比较低,蛋白所带负电荷,其余均为正电荷。稳定性方面,所有的果胶酯酶蛋白都比较稳定。所有多聚半乳糖醛酸酶的大小相差不大,氨基酸序列长度范围为393~460 aa,相对分子质量41 257.55~49 449.76 Da,其中番茄、葡萄、桃、枣的多聚半乳糖醛羧酶所带负电荷,等电点较低,甜樱桃为中性电荷,等电点为7.12,拟南芥、白梨、苹果、甜橘、甜樱桃则带正电荷。稳定性方面,苹果多聚半乳糖醛酸酶表现较为不稳定,其余物种的多聚半乳糖醛酸酶酶都相对稳定(表2)。
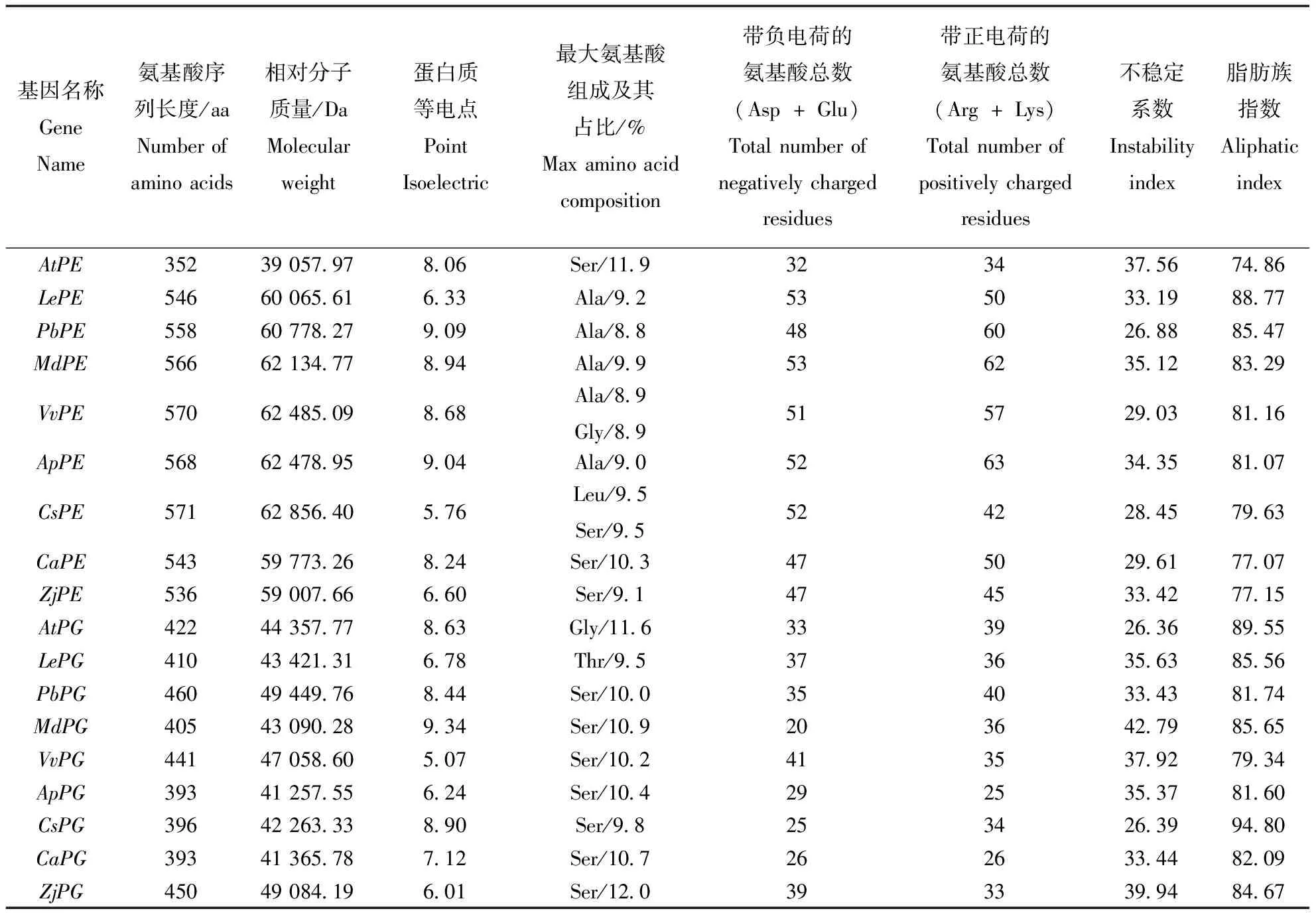
表2 蛋白质理化性质
蛋白质的亲水性和疏水性是组成其所有氨基酸亲水性和疏水性的平均表现,氨基酸的亲水性或疏水性是由所带基团决定的,根据相似相溶,极性基团能够与水亲和,而非极性基团亲和力则比较小。通过对拟南芥和8种水果的果胶酯酶和多聚半乳糖醛酸酶的亲水性和疏水性进行预测(表3):拟南芥(图1a)和所有果胶脂酶都表现为亲水性,番茄果胶脂酶亲水性较小,仅为-0.086。在所有的多聚半乳糖醛酸酶中,拟南芥(图1b)、苹果、桃、甜橙、甜樱桃的多聚半乳糖醛酸酶表现为疏水性,其余4种则表现为亲水性。在所有的酶蛋白亲水性和疏水性预测中,最强疏水性位置多在丙氨酸(Ala)、缬氨酸(Val)、亮氨酸(Leu)、异亮氨酸(Ile)、苯丙氨酸(Phe)等非极性氨基酸上和少数在中性天冬酰胺(Asn)上,最强亲水性位置多在谷氨酰胺(Gln)、半胱氨酸(Cys)、酪氨酸(Tyr)等极性氨基酸和天冬氨酸(Asp)、谷氨酸(Glu)等酸性氨基酸上。通过了解蛋白质的亲水性和疏水性,可以知道其作用环境,在亚细胞定位上也有一定程度的指示作用。
2.2 蛋白质跨膜结构及亚细胞定位分析
蛋白质的跨膜结构在蛋白质转运及与质膜结合行使功能时有重要作用。通过对所有酶蛋白进行跨膜结构预测和亚细胞定位分析(表4):除甜樱桃果胶酯酶(图2b)、甜橙多聚半乳糖醛酸酶(图2c)、枣多聚半乳糖醛酸酶(图2d)3种没有跨膜结构,其余的酶蛋白都与拟南芥果胶酯酶(图2a)一样,有大约20 aa大小的跨膜结构。定位预测结果显示:大部分酶蛋白存在于细胞外,少部分分布在细胞质膜上,而枣多聚半乳糖醛酸酶仅有37%的概率存在于细胞外,在不含有跨膜结构的情况下,推测其还可能存在于细胞质或液泡中。
表4 蛋白质跨膜结构及亚细胞定位
Table 4 Protein transmembrane structure and subcellular localization

基因名称GeneName跨膜结构位置Transmembranestructurelocation亚细胞定位Subcellularlocalization概率ProbabilityAtPE13~35细胞外60.4%LePE20~42细胞质膜79.0%PbPE21~43细胞质膜79.0%MdPE7~29细胞外82.0%VvPE7~29细胞外82.0%ApPE7~29细胞外82.0%CsPE7~24细胞外82.0%CaPE-/-内质网膜82.0%ZjPE7~29细胞外76.7%AtPG13~35细胞外81.0%LePG7~29细胞外82.0%PbPG7~29细胞外59.0%MdPG13~35细胞质膜79.0%VvPG7~26细胞外62.8%ApPG7~29细胞外82.0%CsPG-/-细胞外76.2%CaPG7~29细胞外82.0%ZjPG-/-细胞外37.0%
2.3 O、N-糖基化位点、磷酸化位点预测
糖基化对蛋白质包装及转运、蛋白质稳定性及功能调节等多个方面都有影响,它是蛋白质加工完成后对蛋白质的修饰作用,根据所连接的糖苷链的不同,可以分为N-糖基化和O-糖基化,前者开始于内质网,最终完成在高尔基体,后者整个过程都是在高尔基体中完成。磷酸化是另一种蛋白质修饰进而调节蛋白质功能和活力的方式,磷酸化位点主要在氨基酸序列的丝氨酸(S)、酪氨酸(Y)、苏氨酸(T)残基上。由表5和图3可以看出,在果胶酯酶中,拟南芥糖基化位点(4个)和磷酸化位点(50个)最少,甜橙糖基化位点(22个)和磷酸化位点(72)最多;就多聚半乳糖醛酸酶而言,苹果、葡萄、桃的糖基化位点均为2个,其中苹果无O-糖基化位点,葡萄无N-糖基化位点,枣的糖基化位点最多为17个,且磷酸化位点(58)也是最多,磷酸化位点最少的是甜橙,仅为39个,而且甜橙没有O-糖基化位点。
2.4 蛋白质二级及三级结构预测
蛋白质二级结构是其氨基酸序列的主链在氢键作用力下,进行螺旋、折叠等一些简单的空间排列。通过预测发现(表6):在果胶酯酶中,拟南芥的α-螺旋仅为15.34%,其余物种的α-螺旋则在30.41%~36.80%,β-转角处于一个相对稳定的区间,在4.26%~7.04%,无规卷曲在序列中所占比例最大,占比为36.97%~48.86%,拟南芥扩展链(侧链)最大为31.54%,其余扩展链占比大约为20%;在多聚半乳糖醛酸酶中,无规卷曲在氨基酸序列占到近一半的比例,扩展链占32.22%~40.97%,α-螺旋、β-转角所占比例相对较小。
蛋白质三级结构是在二级或超二级结构的基础上,在疏水相互作用、氢键、范德华力和静电作用等作用力的加持下,进一步盘曲折叠形成的稳定的空间结构。通过对所有酶蛋白结构预测发现:果胶酯酶是由小的β-折叠片层通过无规卷曲链接在一起,在经过α-螺旋形成的三维结构,中间或旁边还链接有一些小的α-螺旋结构,除了拟南芥之外的其他物种的三级结构中还有由3或4个长的α-螺旋结构域转角连接在一起的结构,2个部分共同组成果胶脂酶(图4)。多聚半乳糖醛酸酶和果胶酯酶结构上有些类似,都是由小的β-折叠片层链接在一起,再经过α-螺旋形成三维结构,但是多聚半乳糖醛酸酶的小α-螺旋片段连接在侧链或者序列末端,而不是夹在其间,或者独立成结构(图5)。
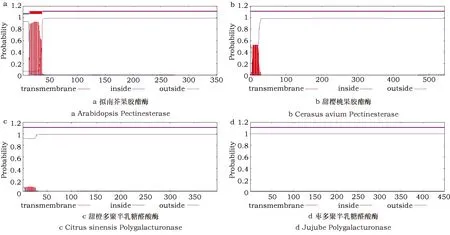
图2 跨膜结构分析
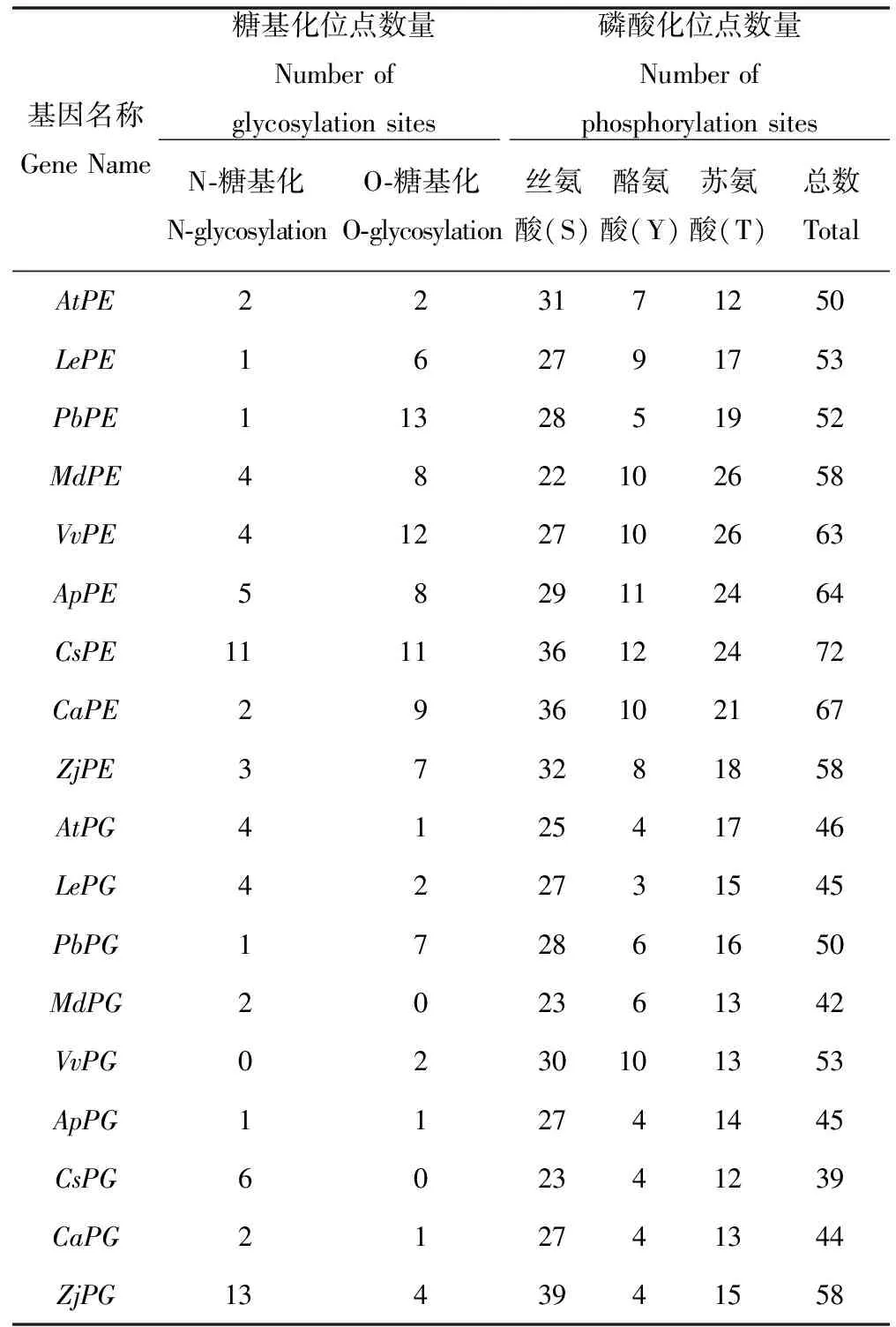
表5 蛋白质修饰位点
2.5 进化树的构建及保守结构域预测分析
进化树和序列比对均能揭示物种或基因之间的同源性及亲缘关系,通过构建系统发育进化树(图6)及序列比对(图7)得出:果胶脂酶中,拟南芥为独立一支,8种水果为一大支,其中枣与甜樱桃、番茄与白梨、葡萄与桃和苹果各自成小支,序列比对同源性为58.23%,结合保守结构域分析(图8a)得出结构上分类与进化树结果一致,拟南芥只有4个motif,其余物种则有11~13个motif,其中motif 1、motif 2、motif 3、motif 4为所有物种所具有,并且motif 1中TFNG序列、motif 2中AVARCYQDGDFig.序列、motif 3中LGRPWYRVPGWWTYEG序列、motif 4中FW序列高度保守,与序列比对结果相一致;在多聚半乳糖醛酸酶的分析中,枣为独立一支,其余物种为一支,甜樱桃与桃和苹果、葡萄与白梨、番茄与拟南芥各自为支,通过序列比对及保守结构域分析(图8b),发现它们的同源性为57.33%,且除了枣中仅有4个motif外,其余8个物种含有10~12个motif,motif 1、motif 5为共有的motif。

图3 拟南芥果胶酶蛋白质修饰位点
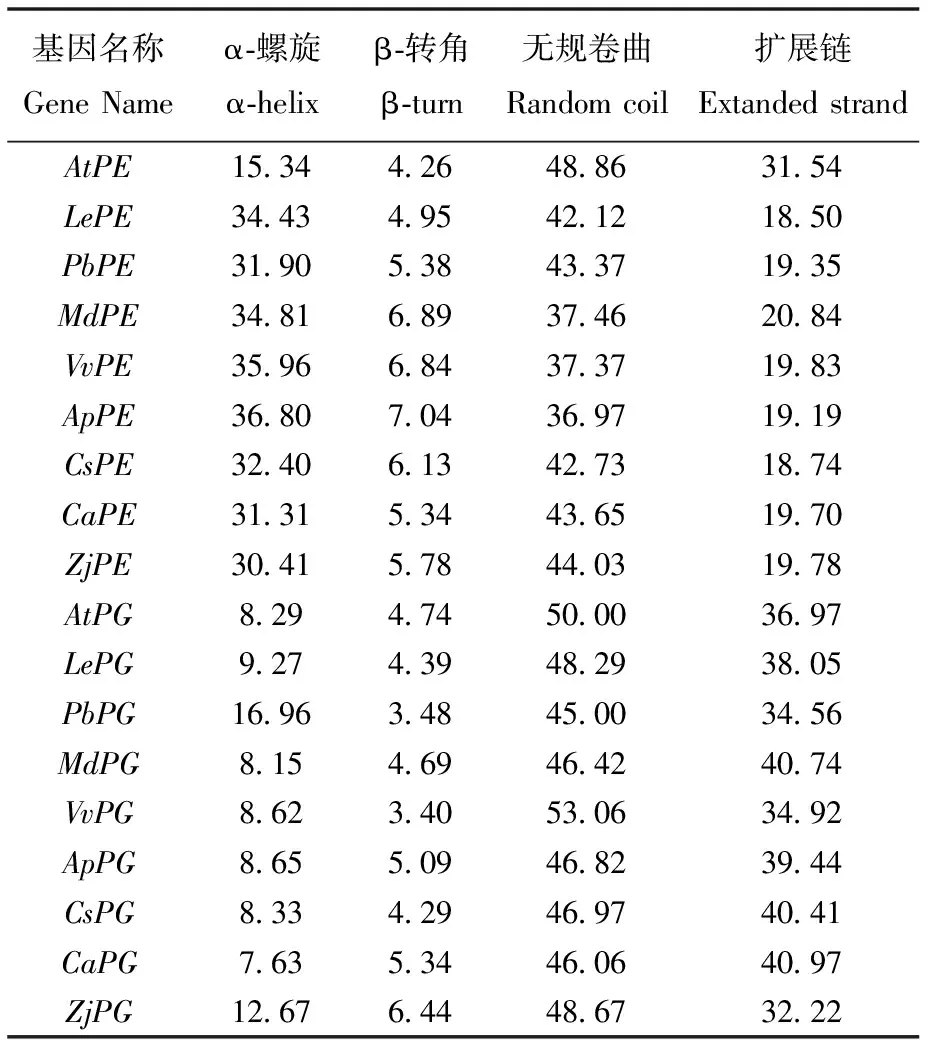
表6 蛋白质二级结构
3 讨论与结论
果胶酶作为果实裂果过程中一种关键酶,其功能主要是分解细胞壁中的果胶,破坏或降低细胞壁的延展性,从而增加了裂果的发生。在前人研究中发现,Ca2+可以有效的降低果胶酶的活性,如通过Ca2+处理的番茄,易裂果和耐裂果番茄中的果胶酶活性分别降低57.06%和30.21%,总果胶(原果胶和可溶性果胶)含量分别增加93.20%和60.19%[3];同样用Ca2+处理不同时期的壶瓶枣,发现果皮和果肉中的果胶酶活性分别下降84.5%~89.9%和76.2%~78.9%,原果胶含量分别增加110.4%~160.7%和212.3%~304.3%[10]。目前在苹果、番茄、枣等多种水果的研究中初步窥探到果胶酶是一种裂果病变表达上调的酶,为了进一步了解其作用机制,在分子水平上进行分析显得尤为重要。
本文以拟南芥为对照,分析对比了8种易裂果水果的果胶脂酶和多聚半乳糖醛酸酶,结果表明:果胶脂酶大小在352~571 aa,且大部分果胶酯酶蛋白都有一个22 aa的跨膜结构域,亚细胞定位多存在与细胞质膜或者细胞外,是一种亲水性蛋白,且多种蛋白质修饰来丰富和稳定其功能。而多聚半乳糖醛酸酶亚细胞定位多存在于细胞外,大小为393~460 aa不等,亲水性和疏水性在不同的物种中存在差异。这些结果与马婧[17]在杨树果胶酯酶基因家族的研究中的结果相一致,并且它们具有一样的motif基序。本文较全面的分析了这2种酶的理化性质,为进一步从基因或蛋白等分子水平上研究裂果病变机理、裂果病变防御提供了理论基础。
本文通过分析得出,果胶酶在不同物种中同源性较高,进化比较保守,所以推测裂果病在不同物种中的作用机制可能存在一定程度的相似性,虽然喷施Ca2+在不同易裂果水果中都有一定的防治效果,但并不能从根本上防止裂果的发生,效果并不是特别理想。Dautt-Castro等在芒果、葡萄、柑橘等水果中对果胶酶在分子水平进行了一些研究[18~20],能够初步了解果胶酶在果实成熟过程中作用机制,结合运用生物信息学分析及基因编辑等新技术,寻找降低果实成熟过程中果胶酶表达水平的途径,有望有效地从根本上降低或防止裂果的发生,同时也为分子水平培育新品种提供新的启示。

图4 不同物种果胶酯酶三级结构
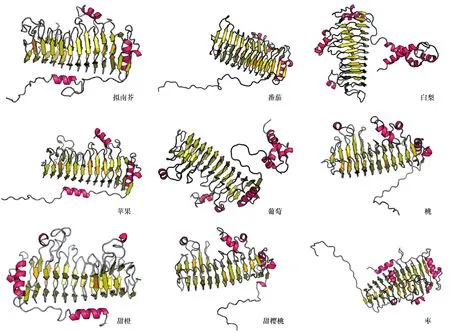
图5 不同物种多聚半乳糖醛酸酶三级结构
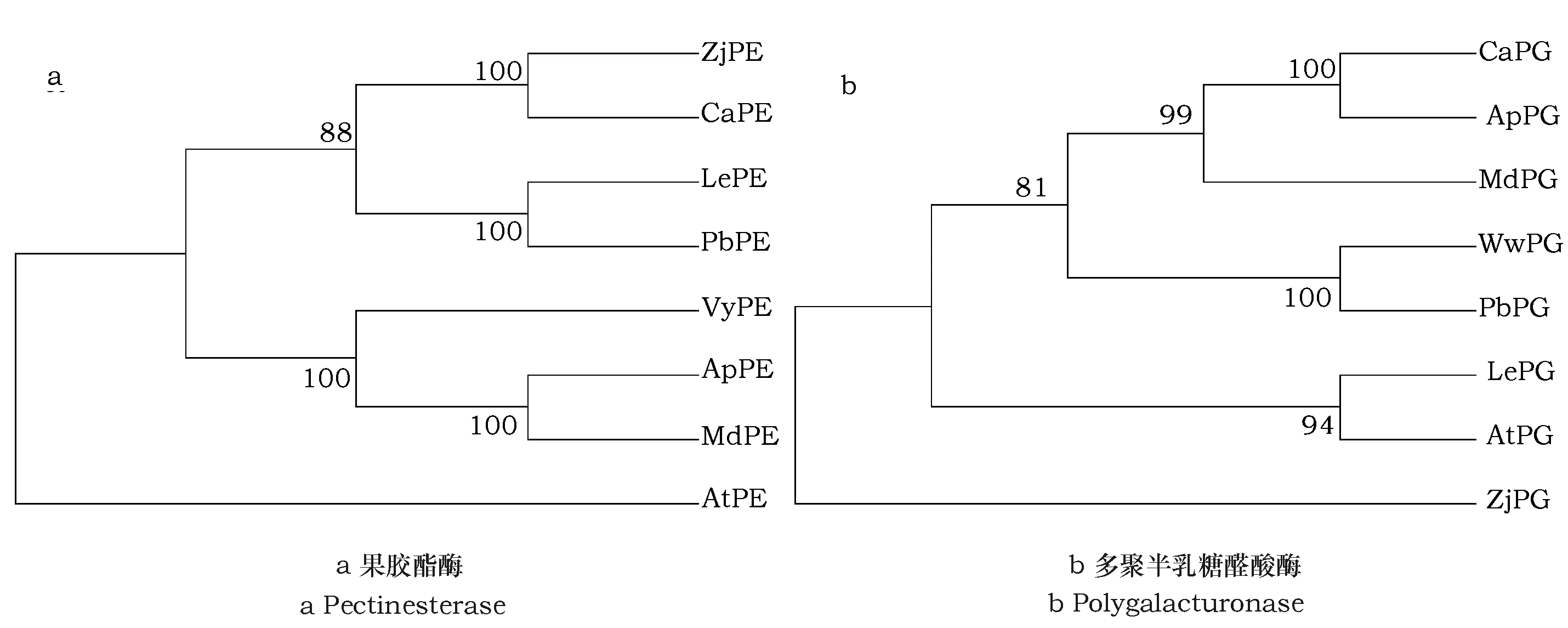
图6 系统发育进化树

图7 序列比对分析

图8 保守结构域预测
