Na2CO3对黄秋葵种子萌发及生理特性的影响
2020-04-20
(山西农业大学生命科学学院,山西 太谷 030801)
黄秋葵(Abelmoschusesculentus(Linn.) Moench)又名咖啡黄葵,为锦葵科秋葵属,一年生草本植物,广泛栽培于热带和亚热带地区[1]。黄秋葵以采食嫩果为主,其花、种子均可以入药,有研究表明黄秋葵在抗疲劳、抗氧化、降血糖和降血脂、调节免疫功能、镇静催眠以及抗肿瘤等[2-4]方面有一定效果,具有广阔的药用前景。
目前,在世界范围内约有8.31亿hm2的土壤由于盐碱化程度过高而无法被有效利用,我国盐碱地总面积达9 913万hm2,约占全国土地面积的10%[5,6]。正常情况下,植物细胞的渗透压高于外界环境以利于吸收水分和矿物质。但高浓度的碱溶液会引发渗透胁迫能快速损害植物,而且对植物幼嫩组织的危害较大[7,8]。种子萌发期是植物生活史的起点,在此期间,其生理活动很容易受到外界环境的干扰,从而影响植物的生长和发育,而碱胁迫就是影响种子萌发的重要因素之一[9,10]。根系是最早暴露在土壤环境下的器官,萌发期的胚根对碱胁迫敏感。植物能否在碱环境中生存首先取决于它能否发芽,其次是取决于它是否能够抵抗出苗后的碱环境[11]。在碱胁迫下,种子的萌发状况可以直观反映种子受胁迫的程度。种子受碱胁迫时,MDA含量积累,为减轻逆境对种子造成伤害,抗氧化酶(Catalase,CAT;Peroxidase,POD;Superoxide dismutase,SOD)保护系统发挥作用,同时积累大量渗透调节物质(如脯氨酸)作为渗透调节剂,提高细胞渗透压,降低细胞内水势[12]。通过测定胚根的这些生理指标,可以从一定程度上了解种子萌发期对逆境的耐受性。
目前国内外对黄秋葵的研究主要集中在药食两用方面,关于逆境生理研究大多为盐胁迫[13,14]与水分胁迫[15],对其碱胁迫下的研究尚有欠缺,本试验采用不同浓度的Na2CO3溶液模拟不同碱性条件,研究不同浓度的碱胁迫对黄秋葵种子萌发及胚根生理特性的影响,探讨黄秋葵种子对Na2CO3的耐受能力,以期为生产实践中黄秋葵的碱化地栽培提供可靠的理论基础。
1 材料与方法
1.1 试验材料
供试材料为“台湾五福”黄秋葵。碱胁迫试剂为无水Na2CO3(分析纯)。
1.2 试验方法
1.2.1种子发芽方法
试验设置5组Na2CO3碱胁迫浓度梯度,每组4个重复,每个重复50粒种子。5组碱胁迫浓度分别为0(ck)、10、20、30、40 mmol·L-1。发芽试验于25 ℃下恒温培养箱内进行,采用标准纸上发芽法[16]。
1.2.2种子萌发参数测定及统计方法
从种子放入培养箱的次日开始,每天观察记录黄秋葵种子的发芽状况,统计发芽势、发芽率和发芽指数,并计算相对碱害率。黄秋葵种子的发芽势以前3 d种子的发芽总数计算,发芽率以7 d种子的发芽总数计算,以胚根露白2 mm视为发芽。发芽7 d后对能萌发为幼苗的处理组进行复水5 d观察其根系生长状况。各指标计算公式如下[17,18]:
发芽率(%)=(7 d内发芽种子总数/供试种子总数)×100%;
发芽势(%)=(前3 d发芽种子总数/供试种子总数)×100%;
发芽指数=∑(Gt/Dt)(式中,Gt表示在第t天种子的发芽数,Dt代表相应的发芽天数);
发芽率相对碱害率(%)=[(对照组发芽率-各处理组发芽率)/对照组发芽率]×100%;
发芽势相对碱害率(%)=[(对照组发芽势-各处理组发芽势)/对照组发芽势]×100%;
发芽期综合碱害率(%)=(发芽势相对碱害率+发芽率相对碱害率)/2。
1.2.3胚根长度测定方法
种子发芽后第3、4、5、6天,从各处理中分别挑选10粒发芽种子测量其根长,观察各处理组胚根生长状况,拍照记录并计算各处理组的胚根生长速度,公式如下:
胚根平均每天的生长速度=[(第4天胚根长度-第3天胚根长度)+(第6天胚根长度-第5天胚根长度)]/2。
对能萌发幼苗的处理组复水5 d后,观察其侧根生长数量。
1.2.4生理指标测定方法
待发芽至第3天,挑选各处理已发芽种子的胚根,测定抗氧化活性酶(POD、SOD、CAT)、脯氨酸和MDA[19-21],每个处理重复3次,取平均值。
1.3 试验数据处理
运用SPSS 23.0软件对数据进行方差分析,并采用最小显著差异法(LSD)进行多重比较分析。同时采用Microsoft Excel 2016软件对所有数据进行统计分析并作图。
2 结果与分析
2.1 碱胁迫对黄秋葵种子萌发特性的影响
黄秋葵种子通过不同碱浓度处理后,其发芽率、发芽势、发芽指数以及相对碱害率的趋势如图1所示。从图中可以看出前三者均随着碱胁迫浓度的增加基本呈降低的趋势,表明碱胁迫能抑制黄秋葵种子萌发。黄秋葵种子萌发的发芽率、发芽势及发芽指数在20~30 mmol·L-1浓度区间均出现一个“平台期”,而在40 mmol·L-1又大幅下降,说明碱胁迫可能已超过黄秋葵种子自身可承受范围。黄秋葵发芽势与发芽率的相对碱害率随碱浓度增加呈增加趋势,且在不同浓度的碱胁迫下,发芽势的相对碱害率均高于发芽率的相对碱害率,说明碱胁迫可以显著延缓种子的萌发。
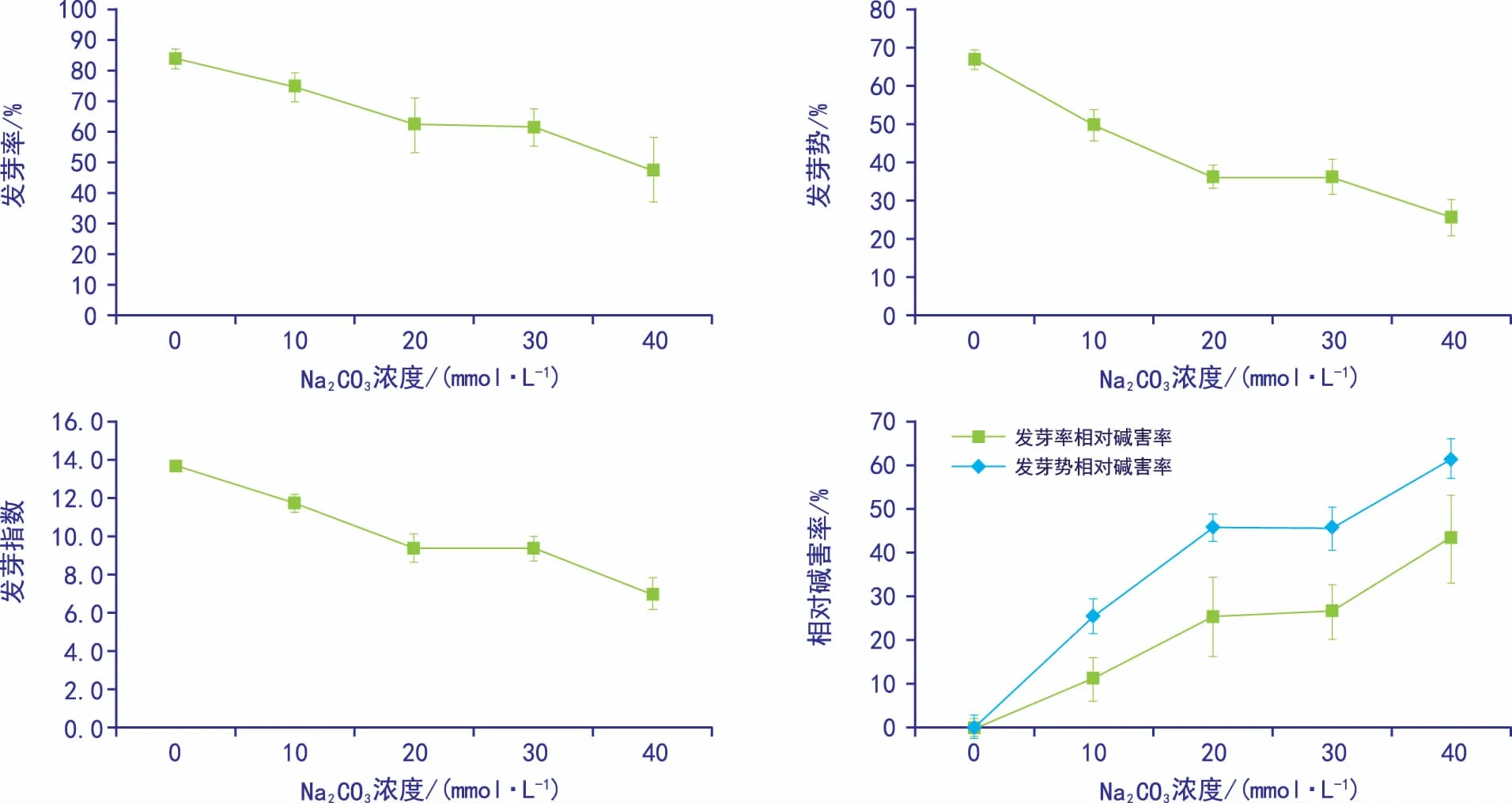
图1 碱胁迫对黄秋葵种子萌发的影响
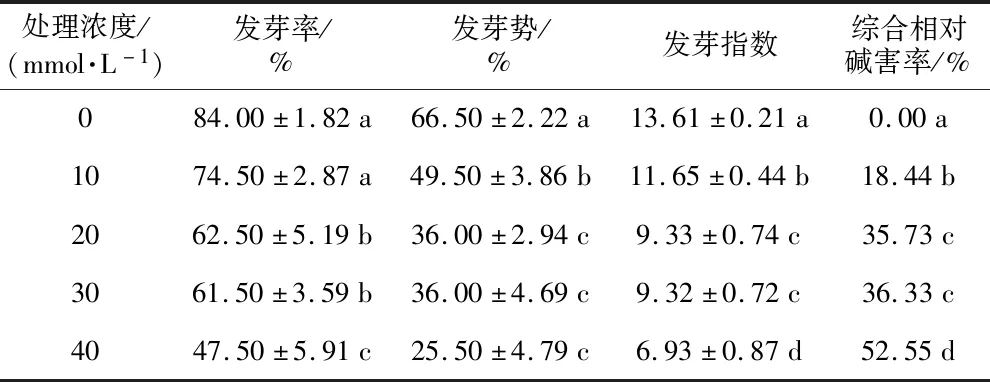
表1 不同浓度碱胁迫对黄秋葵种子萌发的影响
注:不同小写字母表示(p<0.05)差异显著。下同。
通过统计黄秋葵种子萌发参数(见表1),可以发现10 mmol·L-1Na2CO3胁迫的发芽率与对照组差异不显著,说明较该浓度碱胁迫对黄秋葵种子发芽率的影响较小。20 mmol·L-1和30 mmol·L-1处理组发芽率、发芽势和发芽指数无显著差异,其它处理组随碱浓度的升高而减小,综合相对碱害率则相反。而40 mmol·L-1处理组发芽率仅为47.5%,表明高浓度碱胁迫能抑制黄秋葵种子萌发。
2.2 碱胁迫对黄秋葵种子胚根生长的影响
测量种子萌发过程中胚根的长度,统计各处理组胚根的生长速度,见表2。结果表明,黄秋葵种子萌发过程中各处理组胚根长度与碱胁迫浓度呈负差异达显著水平,随着Na2CO3浓度的升高,胚根生长速度也大大降低,20~40 mmol·L-1处理组胚根生长速度均低于10%,说明高于20 mmol·L-1的碱胁迫能严重抑制胚根的生长。
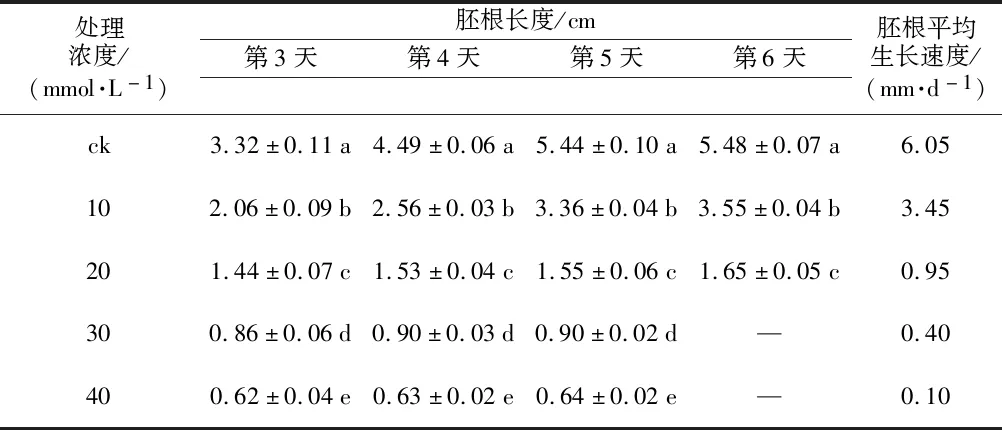
表2 不同浓度碱胁迫对黄秋葵种子新生胚根的影响
观察种子萌发7 d后各处理组胚根生长状况(图2,A),可以发现至第7天时,20~40 mmol·L-1处理组的胚根也出现断裂坏死,只有对照组和10 mmol·L-1处理组具有萌发为幼苗的能力,对其复水5 d(图2,B),观察到对照组根系的侧根数量多于10 mmol·L-1处理组,说明碱胁迫能对新生幼苗的根系造成一定程度的损伤。
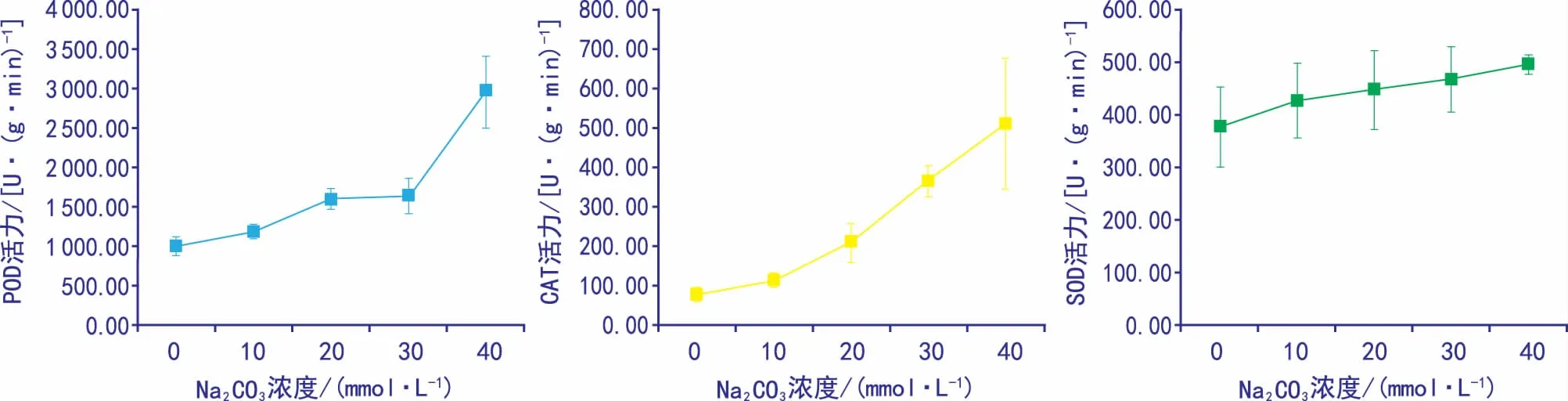
图3 碱胁迫对黄秋葵种子保护酶类活性的影响
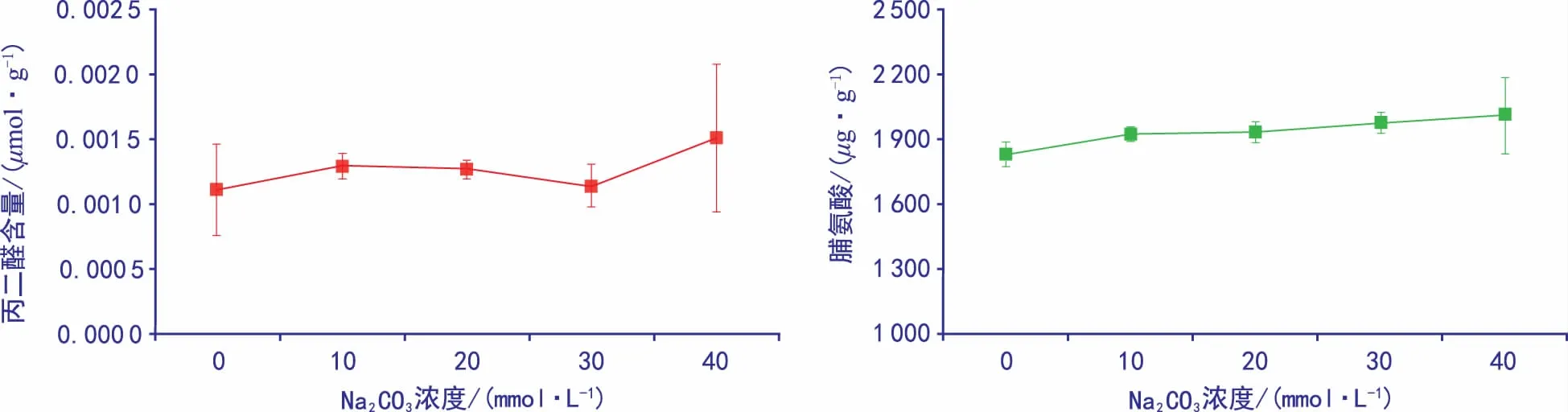
图4 碱胁迫下MDA含量与脯氨酸积累量变化
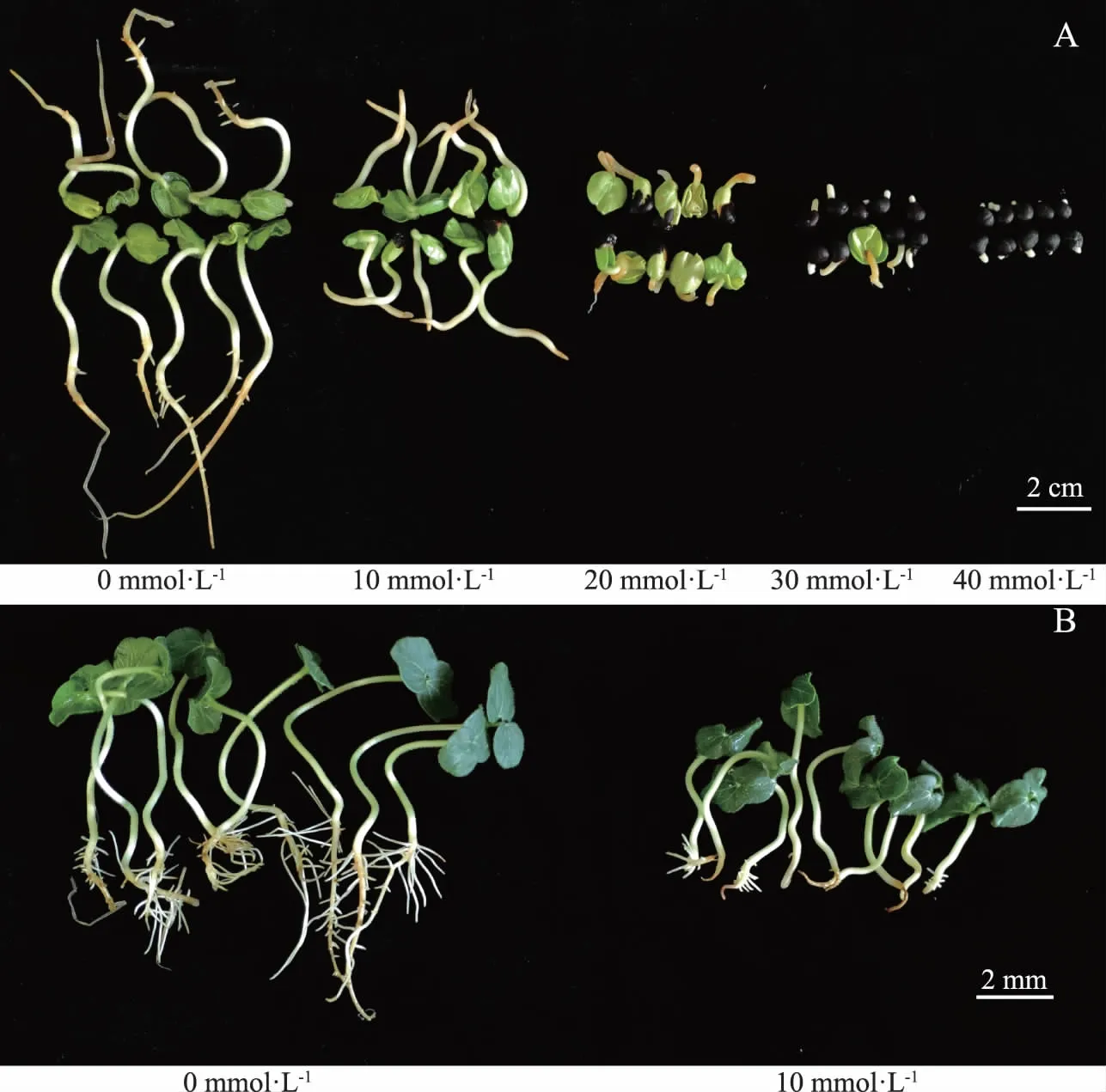
注:A为种子萌发第7天胚根生长图;B为复水5 d后幼苗生长图。图2 黄秋葵种子萌发期胚根长对比图
2.3 碱胁迫下黄秋葵的基本生理指标
2.3.1碱胁迫下黄秋葵种子新生胚根保护酶类的活性
表3和图3显示,随着碱胁迫浓度的增加,黄秋葵种子胚根的POD、CAT和SOD活性均表现为升高趋势,在10~30 mmol·L-1Na2CO3胁迫下POD活性与对照组的差异不显著,40 mmol·L-1Na2CO3胁迫下,POD活性为对照组的2.96倍,差异显著。
10 mmol·L-1的CAT活性与对照组无显著差异,20~30 mmol·L-1与对照组有显著差异,分别是对照组的2.74倍和4.75倍,40 mmol·L-1为对照组的6.63倍。但各处理组的SOD活性差异不显著。
2.3.2碱胁迫下黄秋葵种子胚根MDA与脯氨酸含量变化
从图4可看出,MDA含量随着碱胁迫浓度的升高呈上升趋势,脯氨酸的积累量也有所上升。40 mmol·L-1MDA含量比对照组增加72.28%。说明在高浓度碱胁迫下MDA会在新生胚根中大量积累,而脯氨酸含量在40 mmol·L-1碱胁迫下仅比对照组增加9.82%。说明脯氨酸在碱胁迫下积累缓慢,即表明黄秋葵种子在萌发期间脯氨酸对其渗透调节效果不明显。
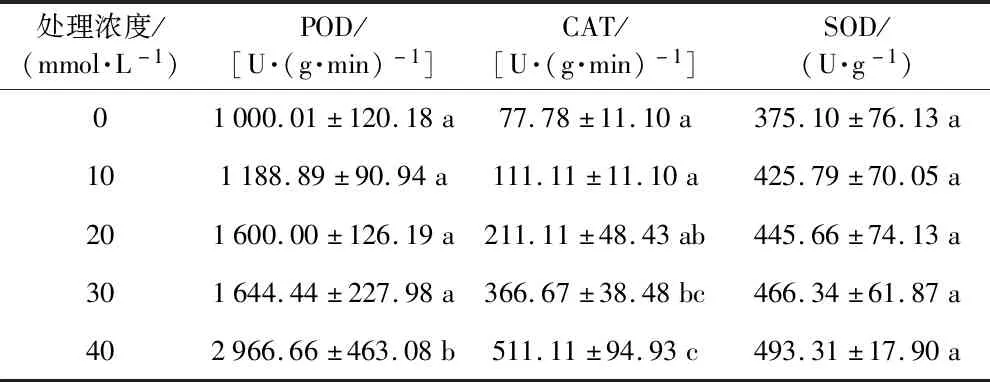
表3 不同浓度碱胁迫对黄秋葵种子保护酶类活性的影响
3 讨 论
在种子生物学领域,常采用发芽势、发芽率、发芽指数等指标来评价种子发芽情况,使用相对碱害率来描述种子在碱胁迫下受胁迫的程度[22]。发芽率可以反映种子发芽的数量,发芽势可以反映种子萌发的快慢和整齐度,发芽指数能够反映种子在整个发芽期的综合活力[23]。本研究结果表明,在Na2CO3胁迫处理下,黄秋葵种子的发芽率、发芽势、发芽指数与碱溶液的浓度呈显著负相关,即抑制效应随碱溶液的浓度增加而增加。碱溶液浓度达到40 mmol·L-1时,种子发芽率已经低于50%,这低于生产所需的发芽率;10 mmol·L-1Na2CO3与对照组相比,发芽势与发芽指数差异显著,说明10 mmol·L-1浓度延缓黄秋葵种子萌发,但二者发芽率差异不显著,仍为74.50%,可能由于黄秋葵种子对10 mmol·L-1碱胁迫具有一定的耐受性,可为碱化地的开发利用提供有价值的参考数据。模拟碱环境测定新生胚根的生长速度可以科学的解释碱胁迫对黄秋葵种子萌发成幼苗期间的危害,本试验中随着碱胁迫浓度的增大黄秋葵种子胚根长度也逐渐减小,且胚根生长速率降低,表明碱胁迫能一定程度抑制种子新生胚根的生长,这与蔺吉祥[23]的研究结果一致。
逆境影响植物体内渗透平衡,破坏膜结构及其组成。有研究结果表明随着碱胁迫浓度的增加,氧自由基含量升高,导致膜脂过氧化程度增加,而膜脂过氧化的产物主要是MDA[24],本研究结果表明,MDA随碱浓度增加呈升高趋势,可能胁迫期间新生胚根的膜脂系统受到了一定程度的破坏。植物大致有2种应对逆境的方法:一种是增加渗透物质如脯氨酸的含量[25],通过增加植物细胞的渗透压来调节植物水势,增强植物的抗逆能力;另一种是为保护细胞免受氧自由基对植物细胞的伤害,细胞产生抗氧化物酶(POD、CAT、SOD)消除过多的氧自由基[26]。本研究发现,脯氨酸的积累量变化并不明显,碱胁迫下黄秋葵种子新生胚根的POD、CAT、SOD活性均有所上升,但只有40 mmol·L-1Na2CO3浓度的POD与CAT活性与对照组有明显差异,而SOD活性在不同碱胁迫浓度处理下均没有显著差异,可能是由于新生的胚根对碱胁迫的适应力较差。
综上所述,碱胁迫能抑制黄秋葵种子萌发及其胚根、幼苗的生长,破坏新生胚根的膜脂系统,但黄秋葵种子对10 mmol·L-1碱胁迫具有耐受性,为黄秋葵选择适宜栽培环境提供了可靠的依据。
