基于GC-MS对水稻种子耐镉差异代谢物的筛选与分析
2020-04-20
(1.天津农学院农学与资源环境学院,天津 300384;2.天津农学院园林园艺学院,天津 300384;3.天津农学院工程技术学院,天津 300384)
重金属镉是世界上毒害作用最强和分布最广的元素之一[1],据2014年全国土壤染污状况调查公报显示[2],我国耕地污染点位超标率为19.4%,其中主要污染物镉超标达7%。在粮食作物中,水稻是较容易吸收和富集镉的植物,我国市面上出售的水稻约有10%存在镉含量超标的问题,“镉大米”已发展成为人类健康的隐患[3],因此,筛选出具有耐镉品质的水稻品种对于食品和人类安全具有十分重要的意义。代谢组学是对所有内源性小分子(分子量小于1 000)代谢物进行定性分析和定量分析的科学[4],这些小分子代谢物是细胞代谢和调控过程的终产物,也是对环境变化和遗传修饰的终端响应,而生物体的表型则是代谢物水平最终的表现[5]。自然界的代谢物有106万种[1,6],其中20万种存在于植物中[4],这些代谢物对植物的生长发育发挥着重要作用,植物对胁迫的响应会通过代谢物的变化反映,例如Mac等[7]和Yamakawa等[8]分别在大麦、水稻体内观测到葡萄糖与ADP葡萄糖含量在高温胁迫下显著积累。因此,利用植物代谢的差异可以有效的区分生理性状与表型变化,同时植物在生长发育过程结束后蛋白质、代谢物和转录本均储藏在种子中[9],Lugan等[10]与Korn等[11]通过对比胁迫敏感型与胁迫耐受型的种质、栽培种或物种的代谢响应方式,可以推断植物在受到非生物胁迫时的差异代谢物;薛水玉等[12]基于GC-MS对款冬花进行代谢组学分析,确定款冬花的合理采摘期为每年的11—12月,此时药效最高;Fiehn[13]课题组用气相色谱与质谱联用仪对4种不同基因型的拟南芥代谢物进行分析研究,并根据代谢产物轮廓对这4种基因型拟南芥进行分类。
1926年丁颖通过分离天然杂交种子进行品种选育,一直将选育出高产、优质、多抗的水稻品种作为育种方向,但田间传统选育方法耗时耗力[14]。肖国樱等在研究中表示,高效是当前和今后水稻育种的主导目标[15]。本研究利用气相色谱-质谱联用仪(GC-MS)的方法对小分子代谢物进行检测,通过统计学方法,找到耐镉水稻品种中的差异代谢产物,将生物学信息与遗传性状整合建模,建立一种高效育种方式,以期为水稻耐镉品种的筛选提供一种新方法,也为指导水稻耐镉品种的筛选提供了理论依据。
1 材料与方法
1.1 材料与试剂
供试材料为天津农学院农学与资源环境学院收集的24份不同遗传背景的水稻种质,-80 ℃冰箱保存。参照张锡洲等[16]方法,将水稻材料耐镉性分为2类,分别为低耐材料14份、高耐材料10份。
1.2 测定方法
1.2.1样品制备
将水稻种子液氮研磨至粉末称取150 mg于5 mL EP管中,加入1 mL体积比为1∶1∶1的乙腈-异丙醇-水混合提取剂,40 kHZ超声提取35 min,置于4 ℃离心机中,10 000 r·min-1离心1 min,取100μL上清液转移至进样小瓶,氮气吹干。进行衍生化处理,取10μL盐酸甲氧胺吡啶溶液(40 mg·mL-1)加入样品中置于30 ℃恒温90 min后,向样品中加入30μL衍生剂(MSTFA+1%TMCS)置于37 ℃恒温30 min,最后加入1 mL溶剂正庚烷(含20μg·mL-1内标物十一烷)过0.22μm微膜,转入GC进样瓶待上机。
1.2.2GC-MS条件
试验仪器采用美国Aglient公司7890 A-5975 C型气相色谱质谱联用仪。气相色谱条件:色谱柱为HP-5 MS(30 m×0.25 mm×0.25μm)毛细管气相色谱柱。载气为纯度99.999%的氦气,进样模式不分流,进样量1μL。进样口温度250 ℃,升温程序为先由常温升至60 ℃维持1 min后,以10 ℃·min-1速度升至280 ℃,最后在280 ℃维持5 min。质谱条件:电子碰撞内能为70 eV;离子源温度230 ℃;四级杆温度150 ℃;溶剂延迟4.5 min;扫描速度为2 scan·s-1,范围50~500 m·z-1。
1.3 数据处理
通过GC-MS光谱全波段扫描水稻种子代谢物质,Agilent-MassHunter工作站对原始数据进行特征提取,最后得到保留时间(rt)及峰面积(intensity);导出数据至Microsoft Excel软件进行整理,通过SIMCA-P进行主成分分析(PCA)及偏最小二乘法-判别分析(PLS-DA),SPSS软件进行t检验(student’s t-test),筛选差异代谢物,利用Omicshare平台进行聚类热图分析(HCA)。
2 结果与分析
2.1 GC-MS分析
通过气相色谱质谱联用仪得到低耐水稻品种与高耐水稻品种典型总离子流色谱图(图1),横坐标表示代谢物质保留时间(rt),纵坐标为峰高(相对丰度),图中不同的出峰时间对应代谢物不同,通过对比低耐与高耐品种总离子流图得出,耐镉水平不同的水稻种子在代谢水平上表现出较明显差异,介于0~10 min低耐品种出峰数量明显低于高耐品种,而16~26 min之间代谢物出峰位置与含量均有不同。
同时在24份水稻样品中共扫描到118个峰,在已有的数据库的支持下根据代谢物的化学结构,对已硅烷化的分子进行还原,对主要代谢物进行定性分析,根据已知内标物含量采用面积归一化法对代谢产物进行定量分析,在水稻种子中得到59种主要代谢产物(表1),可见不同水稻耐镉品种中代谢物含量间存在显著差异,因此,可以通过对代谢物的不同进而分析水稻种子的表型不同。

图1 低耐品种(左)与高耐品种(右)水稻典型总离子流色谱图

表1 不同耐镉水平水稻种子种代谢物定性与定量
注:LC表示低耐镉品种;HC表示高耐镉品种。
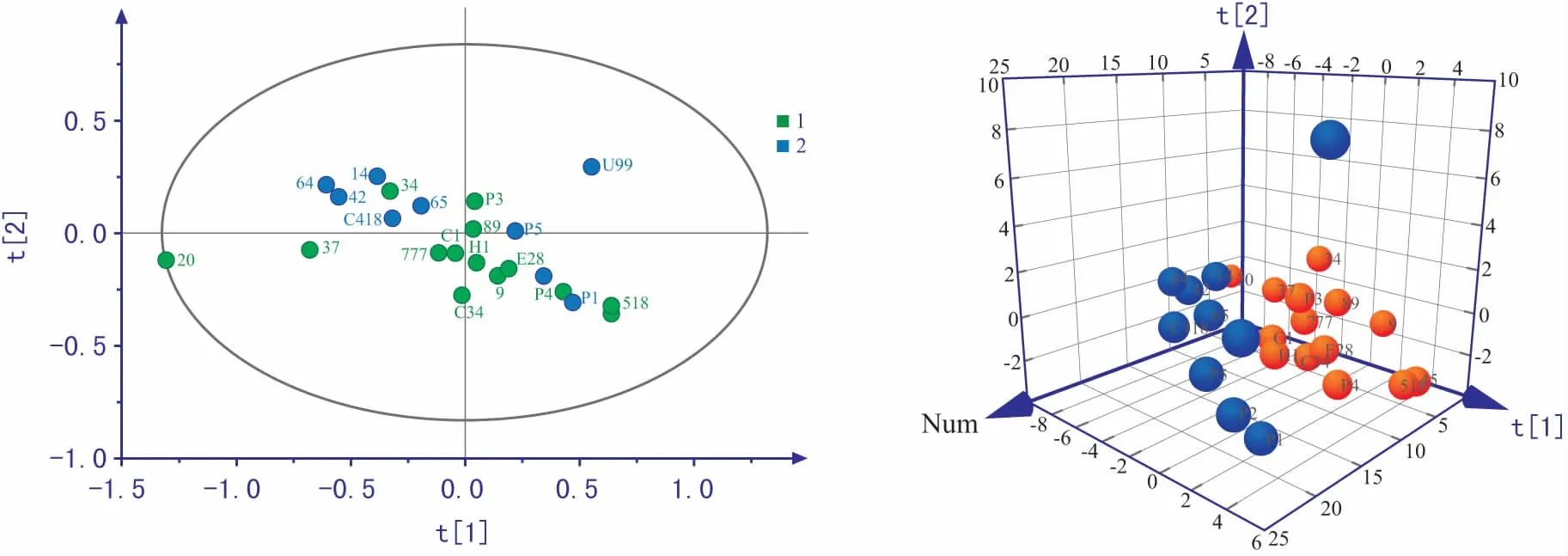
注:图中不同颜色代表不同耐镉水平水稻品种:t 1、与t 2分别代表PC1和PC2。图2 两类水稻种子代谢物主成分二维及三维得分图
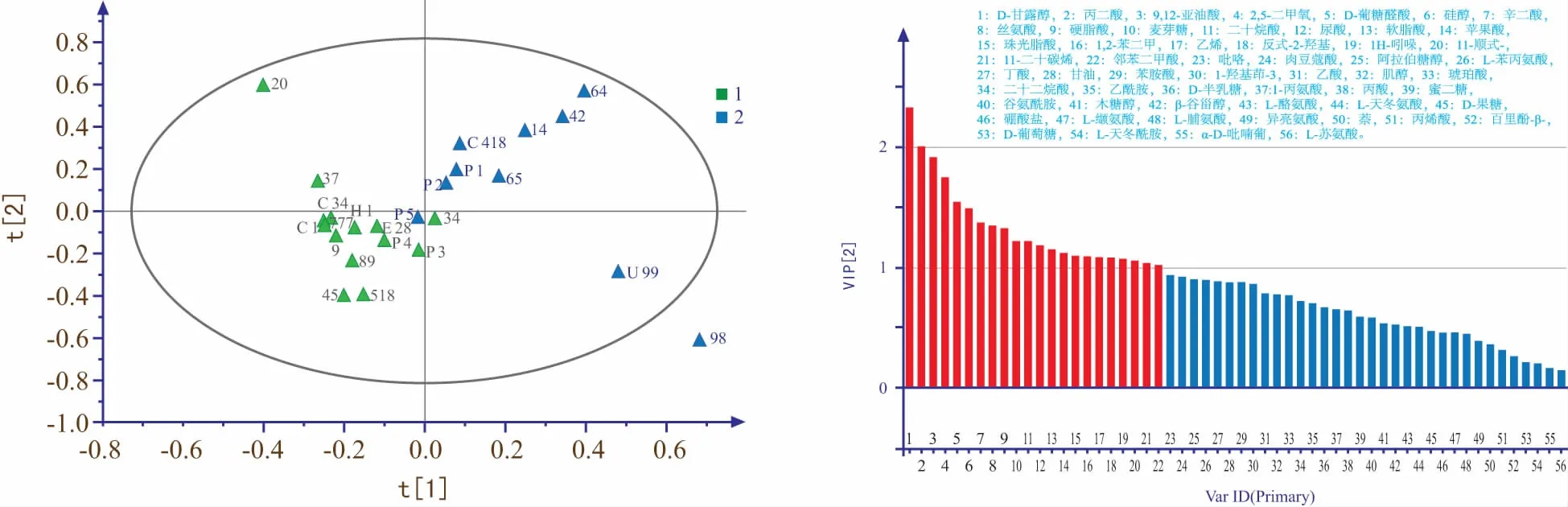
图3 不同水稻种子中代谢物PLS-DA得分图与VIP图
2.2 PCA分析
为了更加清晰的判别不同耐镉程度水稻种子代谢物之间的差异,首先采用了主成分分析(PCA)方法,对不同种水稻种子代谢物采用UV算法得到二维与三维得分图。从图2可以看出,剔除一个异常样品后其他均处于95%置信区间内,第2主成分能将低耐品种与高耐品种区分开但不明显,根据因子分析提取的各代谢物贡献率选取了具有较高载荷系数的代谢物(>0.8)(表1),从图中看出PC2主要反映了2种有机酸(11-顺式-油酸、9,12-亚油酸),说明有机酸在不同耐镉水平的水稻种子中存在差异可能是导致水稻种子耐镉水平不同的原因。但前3个主成分只解释了41.92%的变量,前10主成分累计贡献率才达到80%,无监督分析方法虽能反映数据的原始状态,但不能忽略组内与随机误差,不利于寻找两类水稻种子组间的差异,因此后续将采取有监督分析方法进一步分析差异代谢物。
2.3 PLS-DA分析
通过有监督的偏最小二乘-判别分析对不同水稻品种种子代谢物进行分析,PLS-DA方法可以有效过滤掉与分类无关的信息以更为准确地分析组间差异[17],可以解决PCA分析中对于变量不敏感而不能较好将品种分离的问题。从图3可以看出,通过PLS-DA方法能较好的将两类水稻种子区分开(R2=0.650,Q2=0.147),其中低耐品种分布在置信区间左侧而高耐品种分布在置信区间右侧,且为防止分析结果过拟合对模型进行交叉验证后的点均小于原始R2和Q2,说明PLS-DA相比于PCA模型效果更好,可解释度较高。
2.4 差异代谢物的鉴定
根据PLS-DA分析得到的投影变量权重值(VIP)来反映代谢物对分组的贡献大小,VIP图中柱子越高(图3),贡献越大,差异越显著[18]。再利用SPSS 22.0软件对两类水稻种子中的代谢物进行t检验,根据p值来筛选差异代谢物,当p<0.05时表明差异显著,可筛选出差异标志代谢物[19]。通过结合2种方法(VIP>1,p<0.05)筛选后得出4种差异代谢物(表2),包括3种有机酸(丙二酸、扁桃酸、9,12-亚油酸)和一种醇(D-甘露醇)。其中差异最明显的为D-甘露醇,而其余有机酸类均为高耐品种高于低耐品种且差异倍数在0.12~0.36之间,可见不同耐镉水平的水稻中相同的代谢物的含量也有很大差异,说明水稻的耐镉性在代谢水平上有明显差异,且有机酸与植物的抗逆性紧密相关,是植物体内重要的渗透调节物质[20]。
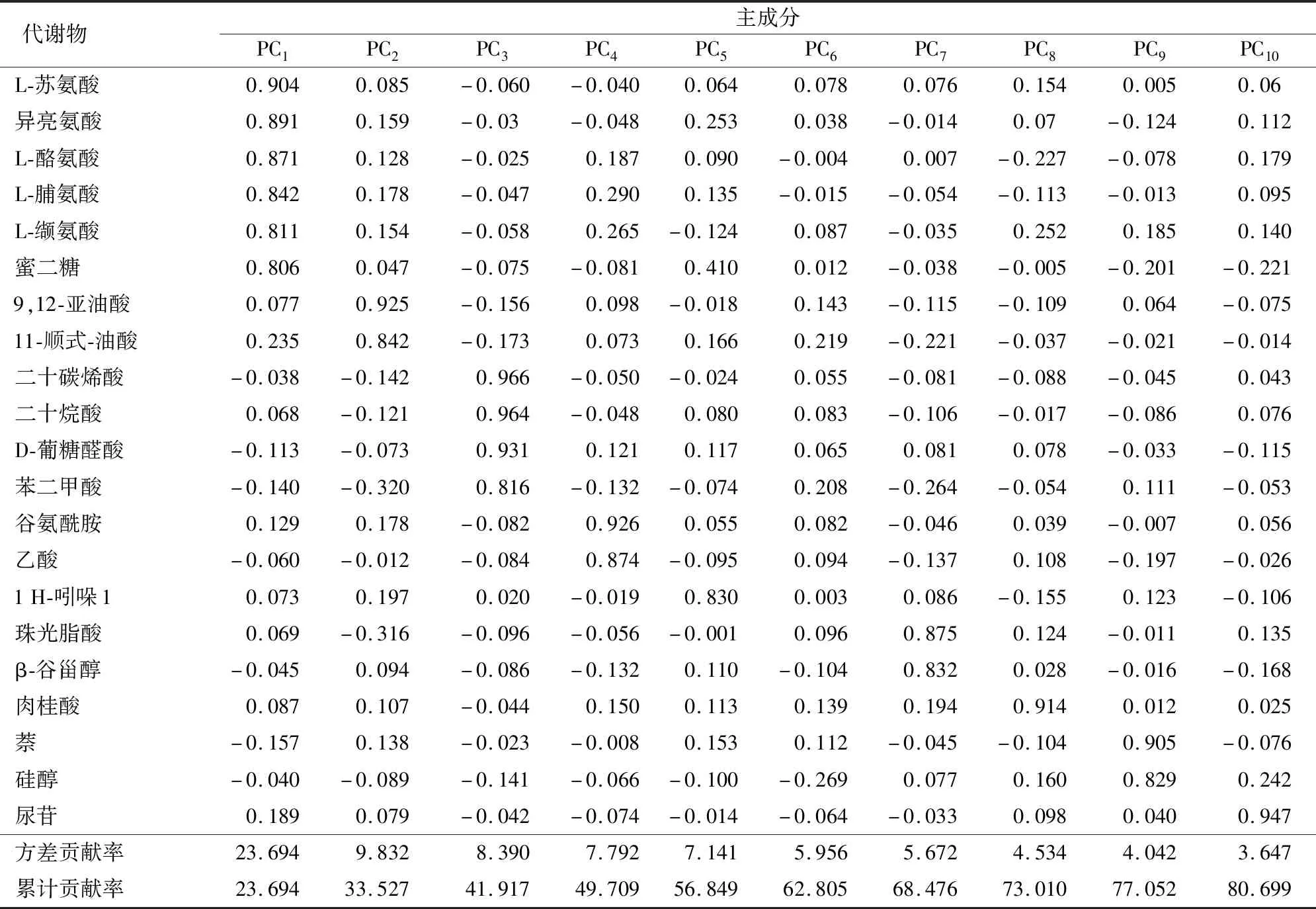
表2 主成分因子载荷旋转矩阵

表3 两类水稻品种差异代谢物
2.5 差异代谢物热图分析
通过Omicshare平台利用pheatmap程序包对寻找到的差异代谢物数据进行归一化处理后进行品种与代谢物的双向聚类(图4),图中横坐标为品种(2为低耐;3为高耐),纵坐标代表差异代谢物,图中颜色由绿到红表示代谢物由低含量向高含量过渡,在图中可以直观的看出,通过4种差异代谢物能较好的将两类水稻种子区分开,同时高耐品种中的丙二酸、扁桃酸、D-甘露醇主要集中在两侧的红色高表达区,而低耐品种集中在中间的绿色低表达区,相反低耐品种中的9,12-亚油酸主要集中在高表达区,高耐品种集中在低表达区,说明低耐与高耐水稻品种在代谢物上有显著差异,可以通过差异代谢物寻找鉴定水稻耐镉水平的方法。同时在图上看出,将扁桃酸与D-甘露醇,9,12-亚油酸与丙二酸聚在了一起,可以说明这些代谢物有相似的功能或参与了相同的代谢途径。
3 讨 论
代谢组学于1997年奥利弗提出至今已被广泛应用于各个领域,如解决植物食品质量问题[20]、研究基因型和环境对植物生长的影响[21],逐渐成为探索内部物质的有力手段[22],Nicholson等[23]研究不同来源的拟南芥发现,不同的生长环境会导致氨基酸和糖的差异,GC-MS在分析植物抗逆性方面也在马铃薯、番茄等作物上取得进展[24]。同时无监督的代谢物分析方法可以更全面的展示植物体代谢物和代谢状态,王希越等[25]应用非靶向即无监督代谢组学研究水稻不同基因型对环境的表型特征。本研究利用气相色谱质谱联用(GC-MS)的方法对两类耐镉程度的水稻种子进行差异代谢物分析,通过无监督的方法筛选出4种差异代谢物,包括3种有机酸(丙二酸、扁桃酸、9,12-亚油酸)和1种醇(D-甘露醇),其中9,12-亚油酸主要存在于低耐品种中。而丙二酸、扁桃酸与D-甘露醇主要存在于高耐品种中,有研究表明,植物的抗逆性与氨基酸和有机酸有关[26],赵秀琴等的研究发现,有机酸含量的高低直接反映植物的生长及代谢活性[24];秦美玲[27]研究表明,有机酸是水稻在镉胁迫下正常生长的原因之一,本研究结果与之一致,说明有机酸是水稻耐镉的重要因素,也可能是水稻耐镉的标志代谢物。但秦美玲[27]的研究结果却是,油酸对植物抵抗镉胁迫的作用较大;Luo等[28]在研究镉胁迫下超积累植物分泌物中发现油酸在缓解镉胁迫起到了重要作用,而本研究结果表明,9,12-亚油酸为低耐水稻种子的标志代谢物,这是由于对于不同的研究对于水稻耐镉的分类不同。
在植物生物学研究发现[29],许多代谢产物同一些重要农艺性状表型相关,Houle[30]在研究中发现这些代谢物是由于基因型的差异及其与环境相互用下产生的,这使得这些代谢物拥有一定的性状预测能力[31]。因此研究水稻种子的耐镉性为后续水稻耐镉品质研究提供重要信息,同时也建立起水稻基因型与表型之间的桥梁,为水稻耐镉品种的选育及耐镉品种的筛选提供理论依据[25]。
