不同壳色合浦珠母贝生产性能评估及其雌雄群体间的差异
2020-04-14魏海军邓正华陈明强邓林琪谭春明李有宁王雨王爱民
魏海军 邓正华 陈明强 邓林琪 谭春明 李有宁 王雨 王爱民

摘要:【目的】綜合评价不同壳色合浦珠母贝(Pinctada fucata)雌雄群体的生产性能,为其良种选育及优质珍珠养殖提供理论指导。【方法】对4种壳色(金壳色、红壳色、黑壳色和白壳色)选育系及常规壳色合浦珠母贝雌雄群体间的数量性状(湿重、壳重、软体重、闭壳肌重、壳长、壳高、壳宽、铰合线长和闭壳肌拉力)和才女虫(Polydora ciliata)感染情况进行系统分析,综合评价其生产性能及雌雄群体间的差异。【结果】除闭壳肌拉力外,4种壳色选育系的数量性状均显著优于常规壳色合浦珠母贝(P<0.05),其中,红壳色选育系可作为以湿重、壳重、软体重、闭壳肌重、壳长、壳高和铰合线长为选育目标的最优选育群体,白壳色选育系可作为以闭壳肌拉力和壳宽为选育目标的最佳选育群体。4种壳色选育系的才女虫感染率均明显高于常规壳色合浦珠母贝,且以红壳色选育系的感染率最高、白壳色选育系的感染率最低。5种壳色合浦珠母贝间的性别分化存在极显著差异(P<0.01),其中,白壳色选育系和金壳色选育系为偏雄群体,红壳色选育系为偏雌群体。雌性合浦珠母贝通常比雄性合浦珠母贝大,但其闭壳肌拉力和抗才女虫感染能力与性别无关。【结论】白壳色选育系和金壳色选育系的综合育种价值较红壳色选育系、黑壳色选育系及常规壳色合浦珠母贝高,建议在今后的合浦珠母贝选育工作中,可根据需求加强对白壳色选育系和金壳色选育系的优化选育,并应用到优质珍珠生产中。
关键词: 合浦珠母贝;壳色;生产性能;才女虫;性别
中图分类号: S968.316.1 文献标志码: A 文章编号:2095-1191(2020)02-0412-09
Evaluation of production performance of Pinctada fucata with different shell colors and its differences between male and female
WEI Hai-jun1,2,3, DENG Zheng-hua2,3, CHEN Ming-qiang2,3, DENG Lin-qi4,
TAN Chun-ming2,3, LI You-ning2,3, WANG Yu2,3*, WANG Ai-min1*
(1College of Marine Science, Hainan University, Haikou 570228, China; 2South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences/Key Laboratory of South China Sea Fishery Resources Exploitation & Utilization, Ministry of Agriculture and Rural Affairs, Guangzhou 510300, China; 3Tropical Aquaculture Research and Development Center, South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Sanya, Hainan 572018, China; 4School of Literature, Hainan Normal University, Haikou 570228, China)
Abstract:【Objective】To reveal the production performance of male and female populations of Pinctada fucata with different shell colors, and to provide theoretical guidance for the breeding of P. fucata and high-quality pearl cultivation. 【Method】Quantitative traits(wet weight, shell weight, tissues weight, adductor muscle weight, shell length, shell height, shell width, hinge length and adductor muscle tension) and the infection status of Polydora ciliata in male and female of P. fucata breeding lines in four shell colors(golden shell color, red shell color, black shell color and white shell color)and normal-shell color were measured for comprehensive evaluation of production performance and differences between male and female populations. 【Result】The results showed that the quantitative traits of the four shell color breeding lines were better than the normal-shell color strains of P. fucata(P<0.05), except for the adductor muscle tension. Among them, the red-shell color breeding line of P. fucata could be used as the most preferred breeding line with wet weight, shell weight, tissues weight, adductor muscle weight, shell length, shell height, and hinge length as the targets of breeding,and the white-shell color breeding line could be used as the most preferred breeding line with adductor muscle tension and shell width as the targets of breeding. The infection ratesof P. ciliate in the four shell color breeding lines were higher than that of the normal-shell color strains of P. fucata, and the infection rate of P. ciliate in the red-shell color breeding line was the highest,the infection rate of P. ciliate in white-shell color breeding line was the lowest. There was extremely significant difference in gender differentiation among five different shell color strains of P. fucata(P<0.01), among which the white-shell color breeding line and golden-shell color breeding line were male-most strains, and the red-shell color breeding line was female-most breeding line. The females were bigger than the males in P. fucata, and the gender had no effect on the adductor muscle tension and the infection rates of P. ciliata. 【Conclusion】The comprehensive breeding values of the white-shell color breeding line and the golden-shell color breeding line are better than those of the red-shell color breeding line, the black-shell color breeding line, and the normal-shell color strains of P. fucata. It is suggested that in the future selection and breeding work, the optimization and selection of white-shell color breeding line and golden-shell color bree-ding line can be strengthened according to demand and applied to high-quality pearl production.
Key words: Pinctada fucata; shell color; production performance; Polydora ciliata; gender
Foundation item: Hainan Natural Science Foundation(319QN338); Special Project of Guangdong Fishing Port Construction and Fishery Development(B201601-Z02); Guangxi Key Research and Development Project (AB18221090); Beihai Scientific Research and Technology Development Project (Beikehe 201995002)
0 引言
【研究意义】合浦珠母贝(Pinctada fucata)又称为马氏珠母贝(P. martensii),是生产海水珍珠的重要贝种之一,主要分布在红海、波斯湾及印度、中国、韩国、日本和西太平洋。近年来,随着珍珠养殖业的持续发展,其养殖环境逐渐恶化,加上近亲繁殖造成种质退化,母贝生长缓慢,死亡率高,育珠质量下降等,严重制约珍珠养殖产业的发展(范嗣刚等,2016)。因此,亟需选育出合浦珠母贝新品种,从根本上改善种质和提高育珠质量。其中,选育生长快的品系可缩短养殖周期,选育壳宽较大的品系可插大核培育大珍珠,选育闭壳肌拉力大且才女虫(Polydora ciliata)感染率低的品系可更好地适应多变的养殖环境而提高成活率,选育雄贝占比高的品系可提高育珠性能,均对珍珠养殖具有重要意义。【前人研究进展】自然界中,大部分動物都是雌雄异体,且雌雄个体在体型及生长方面存在一定差异。在龟鳖类中,黄喉拟水龟(Mauremys mutica)雌、雄个体的生长存在明显差异,表现为雄性个体明显大于雌性个体(朱新平等,2003);平胸龟(Platsternon megacephalun)在1龄前雌、雄性生长无差异,2~3龄时雄性生长快于雌性,但4~5龄时雌性生长快于雄性(郑光明等,2008)。在鱼类中,金钱鱼(Scatophagus argus)雌性个体明显大于雄性个体(吴波等,2013);大菱鲆(Scophthalmus maximus)雌性群体显著大于雄性群体(王新安等,2014);黄颡鱼(Pelteobagrus fluvidraco)雄性个体显著大于雌性个体(马文阁,2016)。在甲壳类水产生物中,中国对虾(Penaeus chinesis)(Browdy,1998)和斑节对虾(Pinaeus mo-nodon)(黄建华等,2013)的雄性个体明显小于雌性个体,中华绒螯蟹(Eriocheir sinensis)雌性个体显著大于雄性个体(何杰等,2016)。在贝类中,目前有关雌雄群体数量性状比较分析的研究相对较少。肖述等(2011)、王冲(2013)在研究香港巨牡蛎(Crassostrea hongkongensis)和栉孔扇贝(Chlamys farreri)时发现雌雄个体间的数量性状不存在显著差异;Iwai等(2015)在研究合浦珠母贝性别对育珠效果的影响时发现雄性个体育珠效果优于雌性个体。贝类的数量性状主要包括湿重、壳长、壳高、壳宽和铰合线长,可在不损伤贝的情况下获取;此外,还有软体部重、壳重、闭壳肌重和闭壳肌拉力等,但这些性状的测量会对贝类造成一定机械损伤(闭壳肌拉力),甚至需要剖解。其中,闭壳肌拉力与贝类的强壮及健康程度密切相关,且已有研究证实闭壳肌拉力与死亡率呈负相关(Poulet et al.,2003;Okamoto et al.,2006)。多毛类凿贝才女虫侵蚀珍珠贝会引起黑壳病和黑心肝病,在合浦珠母贝养殖过程中危害较严重,是造成合浦珠母贝高死亡率的主要原因之一(陈明强等,2015)。王爱民等(2003)研究表明,通过不同地理群体杂交选育可提高合浦珠母贝的抗才女虫侵蚀能力,并证实防治才女虫侵蚀时采用水泥涂覆法较饱和盐水浸泡法、淡水浸泡法和高温海水浸泡法的效果更佳,且操作简单,成本低(王爱民等,2004)。陈明强等(2015)依据后代的才女虫感染情况,从12个全同胞合浦珠母贝家系中筛选出4个抗才女虫的家系。【本研究切入点】自然条件下,无论是野生的还是养殖的合浦珠母贝均存在壳色多样性,壳色分化为金壳色、红壳色、黑壳色和白壳色等。近年来,我国科学家围绕合浦珠母贝4种壳色选育系开展了一系列研究,并取得长足进展,包括4种壳色合浦珠母贝选育系生长速度、数量性状及存活率的比较(邬思荣,2011;王祎哲等,2018),遗传多样性分析(陈静等,2010;朱晓闻等,2012;白成等,2014),插核育珠效果和小片贝选育(符韶等,2012;张文府,2013),壳色相关基因克隆(Zou et al.,2014),以及金属元素含量比较分析(邹柯姝等,2015)等,但关于合浦珠母贝雌、雄群体间数量性状的差异尚无研究报道。【拟解决的关键问题】通过对4种壳色选育系和常规壳色合浦珠母贝的数量性状(湿重、软体重、壳重、闭壳肌重、壳长、壳宽、壳高、铰合线长和闭壳肌拉力)及才女虫感染情况进行系统分析,综合评价其生产性能和雌雄性群体间的差异,为合浦珠母贝的良种选育及优质珍珠养殖提供理论指导。
1 材料与方法
1. 1 试验材料
2017年11月在中国水产科学研究院南海水产研究所热带水产研究开发中心(海南省陵水县新村镇)人工繁育合浦珠母贝4种壳色选育系F7代及常规合浦珠母贝,采用相同的饲养管理方法:在早期浮游阶段(2~5 d)以等鞭金藻(Isochrysis zhanjiangensis)作为主要饵料;在中期浮游阶段(6~15 d)投喂亚心形扁藻(Platymonas subcordiformis)和等鞭金藻的混合物;从后期浮游阶段至附着阶段,主要投喂亚心形扁藻。在育苗期间,每天换育苗池总水量1/3的过滤海水,并维持海水盐度为(32±1)‰,24 h持续充气增氧。待幼苗在附着板上长至2 mm(壳长)左右时,将其从附着板上洗下并放入网袋中移至陵水新村港的贝架进行吊养。在海上吊养期间,需不定期对合浦珠母贝进行清洗和更换网袋,避免因污浊生物附着及密度过大而影响其正常生长和存活。2019年1月随机取14月龄金壳色、红壳色、黑壳色、白壳色选育系及常规壳色合浦珠母贝(图1)各200只,湿重27.3±6.3 g,壳长53.9±5.9 mm。在贝架上清洗干净后移回中国水产科学研究院南海水产研究所热带水产研究开发中心陵水实验基地的水泥池中暂养[盐度(32±1)‰,水温(23±1)℃]以备后续试验。
1. 2 试验方法
先用数显卡尺测量贝的壳长、壳高、壳宽和铰合线长(精确到0.1 mm),然后擦干贝壳表面水分以天平称量湿重(精确到0.1 g);采用数显拉力机测量闭壳肌拉力(精确到0.1 N);解剖贝分辨其性别,再以天平分别称量壳重、软体重和闭壳肌重;最后观察其左壳和右壳的才女虫感染情况。其中,形态性状和质量性状的测量参照刘志刚等(2007)、陈飞飞等(2012)的方法;闭壳肌拉力测量参照Aoki等(2010)的方法,具体操作:将合浦珠母贝放入淡水浸没10 min使其因应激反应而双壳紧闭,然后用数显拉力机(ZQ-21B-1,广东东莞智取精密仪器有限公司)挤压开壳器使贝开口10 mm,并读取最大拉力值即为闭壳肌拉力。合浦珠母贝雌雄辨别采用镜检法:取性腺内容物置于滴有海水的载玻片上,显微镜观察样品形态,如性腺内容物在海水中发散且能看到圆形或梨形卵子即鉴定为雌性个体;若性腺内容物在海水中呈烟雾状即鉴定为雄性个体。才女虫感染情况数据收集参考陈明强等(2015)的方法,清洗贝壳珍珠层后进行观察,若贝壳珍珠层光滑有珍珠光泽,无虫管症状等异常,说明此贝未感染才女虫;若贝壳珍珠层上有小斑点、脓包、溃烂性病灶或虫管等症状(石耀华等,2004),则表明此贝已感染才女虫。
1. 3 统计分析
试验数据使用Excel 2010进行整理,采用SPSS 18.0进行统计分析,以Origin 9.0制图。其中,以Tukey检验法(方差不齐时以Games-Howell检验法)比较不同壳色合浦珠母贝间主要性状(湿重、壳重、软体重、闭壳肌重、闭壳肌拉力、壳长、壳高、壳宽和铰合线长)的差异;以卡方检验分析不同壳色合浦珠母贝的性别分化情况;以t 检验比较不同壳色合浦珠母贝雌、雄群体间主要性状的差异。同时,统计不同壳色合浦珠母贝群体及其雌、雄群体的左壳感染率、右壳感染率、左右壳感染率和总感染率,计算公式如下:
左壳感染率(%):Plij=Σnlij/N×100
右壳感染率(%):Prij=Σnrij/N×100
左右壳感染率(%):Pbij=Σnbij/N×100
总感染率(%):Ptij=(Σnlij+Σnrij-Σnbij)/N×100
式中,Plji表示第ij组合浦珠母贝左壳感染率,nlij表示第ij组合浦珠母贝左壳感染个体数,Prij表示第ij组合浦珠母贝右壳感染率,nrij表示第ij组合浦珠母贝右壳感染个体数,Pbij表示第ij组合浦珠母贝左右壳感染率,nbij表示第ij组合浦珠母贝左壳和右壳同时感染个体数,Ptij表示第ij组合浦珠母贝总感染率,i取值范围为1~5(4种壳色选育系和常规壳色合浦珠母贝各为一组),j取值范围为1~3(不分雌雄群体、雌性群体和雄性群体各为一组),N为每组合浦珠母贝总个体数。
2 结果与分析
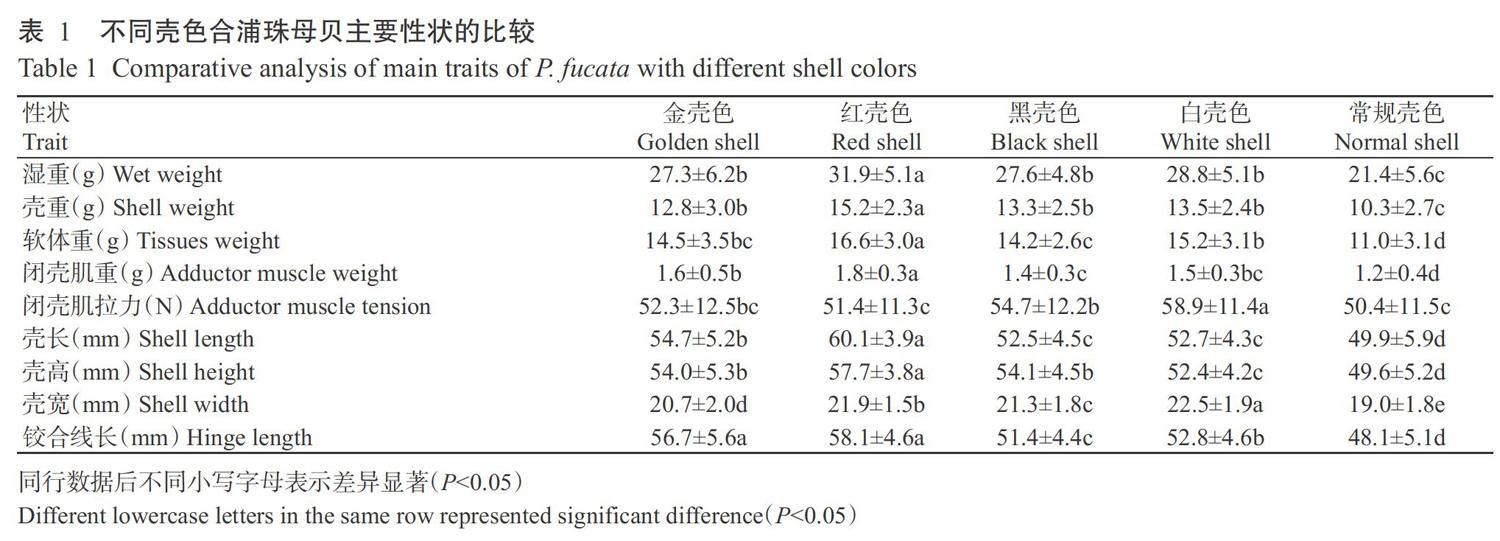
2. 1 不同壳色合浦珠母贝主要性状的比较
5种壳色合浦珠母贝主要性状的比较分析结果如表1所示。金壳色选育系、黑壳色选育系和白壳色选育系间的湿重和壳重差异不显著(P>0.05,下同),金壳色选育系和红壳色选育系间的闭壳肌拉力和铰合线长差异不显著,金壳色选育系和黑壳色选育系间的软体重、闭壳肌拉力和壳高差异不显著,金壳色选育系和白壳色选育系间的软体重和闭壳肌重差异不显著,金壳色选育系、红壳色选育系和常规壳色合浦珠母贝间的闭壳肌拉力差异不显著,黑壳色选育系和白壳色选育系间的闭壳肌重和壳长差异也不显著。综合表1中的各项数量性状指标可知,除闭壳肌拉力外,4种壳色选育系的数量性状均显著优于常规壳色合浦珠母贝(P<0.05,下同),其中,红壳色选育系可作为以湿重、壳重、软体重、闭壳肌重、壳长、壳高和铰合线长为选育目标的最优选育群体,白壳色选育系可作为以闭壳肌拉力和壳宽为选育目标的最佳选育群体。
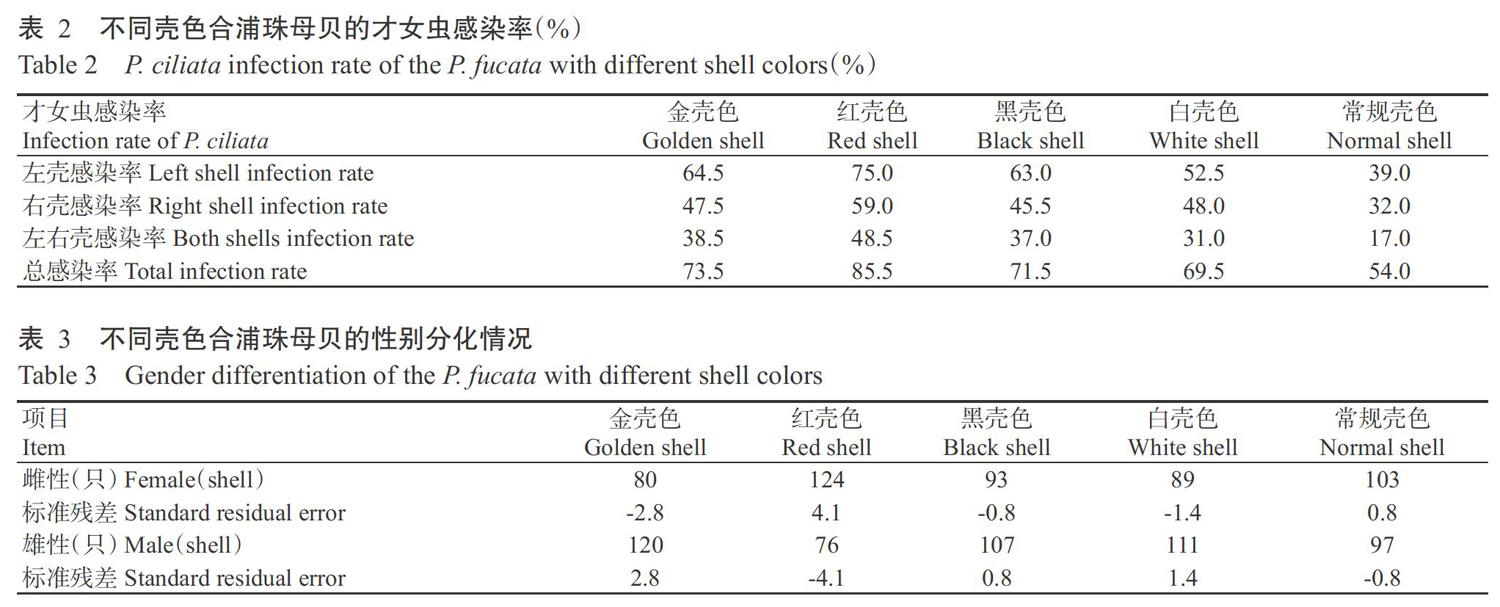
由表2可看出,合浦珠母贝的左壳更易被才女虫感染,且4种壳色选育系的左壳感染率、右壳感染率、左右壳感染率和总感染率均明显高于常规壳色合浦珠母贝。在4种壳色选育系中,红壳色选育系的才女虫感染率最高:左壳感染率为75.0%、右壳感染率为59.0%、左右壳感染率為48.5%、总感染率为85.5%;白壳色选育系的才女虫感染率最低:左壳感染率为52.5%、右壳感染率为48.0%、左右壳感染率为31.0%、总感染率为69.5%。
5种壳色合浦珠母贝的雌性期望计数为97.8只,雄性期望计数为102.2只。卡方检验结果显示,卡方值为22.627(P<0.001),似然比为22.789(P<0.001),说明5种壳色合浦珠母贝间的性别分化存在极显著差异(P<0.01,下同)。由表3(雌雄个体数及其标准残差)可知:红壳色选育系的雌性个体数最多,为124只(标准残差为4.1),而雄性个体数最少,为76只(标准残差为-4.1);金壳色选育系的雌性个体数最少,为80只(标准残差为-2.8),而雄性个体数最多,为120只(标准残差为2.8)。
2. 2 不同壳色合浦珠母贝雌雄性群体间主要性状的比较
以t 检验比较不同壳色合浦珠母贝雌雄性群体间主要性状的差异,结果如图2所示。金壳色选育系的软体重表现为雌性群体显著大于雄性群体,壳宽表现为雌性群体极显著大于雄性群体;红壳色选育系的湿重、壳重、软体重、壳长和壳高均表现为雌性群体极显著大于雄性群体,铰合线长表现为雌性群体显著大于雄性群体;黑壳色选育系的壳长表现为雌性群体显著大于雄性群体;常规壳色合浦珠母贝的湿重、壳重、软体重、壳长、壳高、壳宽和铰合线长均表现为雌性群体极显著大于雄性群体,闭壳肌重表现为雌性群体显著大于雄性群体;白壳色选育系的各项数量性状指标在雌雄群体间均无显著差异。此外,对于闭壳肌拉力而言,5种壳色合浦珠母贝雌雄性群体间均无显著差异,说明闭壳肌拉力与其性别无关。
2. 3 不同壳色合浦珠母贝雌雄性群体间才女虫感染情况的比较
在5种壳色合浦珠母贝中,不论是雌性群体还是雄性群体,均表现为左壳较右壳更易感染才女虫(图3-A和图3-B)。从图3-D可看出,常规壳色合浦珠母贝的才女虫感染率最低,而在4种壳色选育系中以红壳色选育系的感染率最高、白壳色选育系的感染率最低。此外,金壳色选育系和常规壳色合浦珠母贝的雄性群体较雌性群体更易感染才女虫,而红壳色选育系和黑壳色选育系表现为雌性群体较雄性群体更易感染才女虫。
3 讨论
本研究结果表明,不论雌性群体还是雄性群体,4种壳色选育系的生长性能均优于常规壳色合浦珠母贝,但其抗才女虫感染能力明显下降,可能是由于连续以生长性能为选育目标进行闭锁群体选育,而导致其遗传多样性降低及抗才女虫感染能力下降。因此,建议今后繁育这4种壳色选育系时,可挑选野生的4种壳色合浦珠母贝与人工繁育的4种壳色合浦珠母贝选育系进行杂交,且同时以生长性能和抗才女虫感染为选育目标。4种壳色选育系繁育至今已是F7代,综合本研究结果可知,F7代白壳色选育系可作为闭壳肌拉力、壳宽和抗才女虫感染复合选育的最佳育种群体,F7代红壳色选育系则可作为湿重、壳重、软体重、闭壳肌重、壳长、壳高和铰合线长复合选育的最佳育种群体。这与王庆恒等(2008)、陈明强等(2016)研究表明金壳色选育系和黑壳色选育系生长性能最佳的结果有所不同,究其原因可能是选育技术路线、养殖环境和选育基础群体亲本来源不同,以及4种壳色合浦珠母贝的季节性生长差异所致。
本研究还发现,金壳色选育系为偏雄群体(60%雄性),而红壳色选育系为偏雌群体(62%雌性)。赵永超(2013)研究表明雄性三角帆蚌的产珠性能显著优于雌性;Iwai等(2015)针对合浦珠母贝的育珠效果进行研究,也发现雄性合浦珠母贝的育珠效果优于雌性合浦珠母贝。由此可确定,金壳色选育系可作为偏雄群体进行优质珍珠生产。动物雌雄性群体间的生长差异与其消化、生长相关基因表达、性激素水平及生长繁殖能量配置等均有关联(马细兰等,2009)。本研究中,金壳色选育系、红壳色选育系、黑壳色选育系及常规壳色合浦珠母贝雌雄性群体间的数量性状(除闭壳肌拉力外)均表现为雌性群体大于雄性群体,且部分性状指标存在极显著或显著差异,说明雌性合浦珠母贝一般比雄性合浦珠母贝大。这与香港巨牡蛎(肖述等,2011)和栉孔扇贝(王冲,2013)的研究结果不同,可能是繁殖习性及物种遗传本质等导致的物种间差异(朱新平等,2003)。在5种壳色合浦珠母贝中,雌雄性群体间的闭壳肌拉力均无显著差异,说明闭壳肌拉力与合浦珠母贝的性别无关。Okamoto等(2006)研究发现,闭壳肌拉力与贝类死亡率呈负相关、与其滤水能力及闭壳肌糖原含量呈正相关,因此闭壳肌拉力可作为评估贝类强壮程度、健康程度、营养状况及抗逆性的主要指标之一。此外,贝类闭壳肌拉力性状可遗传(Ishikawa et al.,2009)。牛志凯等(2015)通过杂交3个地理群体的合浦珠母贝选育出生长性状和闭壳肌拉力均较优的YNSY群体(越南♂×海南三亚♀)和YNSZ群体(越南♂×广东深圳♀)。本研究在解剖合浦珠母贝时发现,才女虫感染情况严重(有大面积脓包、大面积溃烂性病灶或多处虫管)或病灶集中在闭壳肌处的合浦珠母贝,其闭壳肌拉力较小;才女虫感染情况较轻(感染面积小,珍珠质有星点凸起,尚光滑)或病灶不在闭壳肌处的合浦珠母贝,其闭壳肌拉力较大。这在一定程度上佐证了Okamoto等(2006)的研究结果。本研究在采集才女虫感染率数据时,无差别地对待才女虫感染情况,尚未区分才女虫感染的严重程度及其病灶的具体部位,而简单将有病灶的合浦珠母贝定义为感染、无病灶的合浦珠母贝定义为未感染。因此,今后应系统研究合浦珠母贝闭壳肌拉力与抗才女虫感染能力间的关联性。
本研究对4种壳色选育系及常规壳色合浦珠母贝雌雄性群体的主要性状和才女虫感染情况进行比较,发现合浦珠母贝的雌性个体比雄性个体大,但其性别对闭壳肌拉力和抗才女虫感染的影响不明显。综合评价可知,红壳色选育系的抗才女虫感染能力太弱,常规壳色合浦珠母贝的生长性能太低,且二者均为非偏雄群体,故用于育珠的价值不及白壳色选育系和金壳色选育系。
4 结论
雌性合浦珠母贝通常比雄性合浦珠母贝大,但闭壳肌拉力和抗才女虫感染能力与其性别无关。白壳色选育系和金壳色选育系的综合育种价值较红壳色选育系、黑壳色选育系及常规壳色合浦珠母贝高,建议在今后的选育工作中,可根据需求加强对白壳色选育系和金壳色选育系的优化选育,并应用到优质珍珠生产中。
参考文献:
白成,刘志刚,吴灶和,孙宗红,刘付少梅. 2014. 马氏珠母贝4种壳色家系遗传差异的SSR分析[J]. 广东海洋大学学报,34(1):48-53. [Bai C,Liu Z G,Wu Z H,Sun Z H,Liufu S M. 2014. Genetic diversity of the four shell co-lor families of Pinctada martensii by SSR marker system[J]. Journal of Guangdong Ocean University,34(1):48-53.]
陈飞飞,黄桂菊,陈明强,油九菊,李有寧,吴开畅,邢孔武,喻达辉. 2012. 合浦珠母贝三亚养殖群体生长性状的相关与通径分析[J]. 广东农业科学,39(9):122-125. [Chen F F,Huang G J,Chen M Q,You J J,Li Y N,Wu K C,Xing K W,Yu D H. 2012. Correlation and path analysis of growth-realted traits in Sanya cultured population of pearl oyster Pinctada fucata[J]. Guangdong Agricultural Science,39(9):122-125.]
陈静,刘志刚,孙小真,王辉,杜晓东. 2010. 马氏珠母贝4种壳色选育系F3的生长特性和遗传多样性比较[J]. 上海海洋大学学报,19(5):588-595. [Chen J,Liu Z G,Sun X Z,Wang H,Du X D. 2010. Analysis on growth characteristic and genetic diversity in the selected lines(F3) of four shell colors of Pinctada martensii[J]. Journal of Shanghai Ocean University,19(5):588-595.]
陈明强,刘宝锁,谭才钢,吴开畅,严俊贤,李有宁. 2015. 合浦珠母贝家系间生长性状和感染多毛类寄生病的比较分析[J]. 广东农业科学,42(5):87-91. [Chen M Q,Liu B S,Tan C G,Wu K C,Yan J X,Li Y N. 2015. Comparative analysis on growth traits polychaetes infection in di-fferent Pinctada fucata families[J]. Guangdong Agricultural Science,42(5):87-91.]
陳明强,刘宝锁,严俊贤,郭华阳,吴开畅,李有宁. 2016. 合浦珠母贝4种壳色选育系主要性状的比较分析[J]. 南方水产科学,12(5):118-122. [Chen M Q,Liu B S,Yan J X,Guo H Y,Wu K C,Li Y N. 2016. Comparison analysis of major traits among four shell color shell lines of pearl oyster (Pinctada fucata)[J]. South China Fisheries Science,12(5):118-122.]
范嗣刚,王婧璇,黄桂菊,刘宝锁,郭奕惠,喻达辉. 2016. 合浦珠母贝选育家系的遗传多样性分析[J]. 南方水产科学,12(5):90-96. [Fan S G,Wang J X,Huang G J,Liu B S,Guo Y H,Yu D H. 2016. Analysis of genetic diversity of nine families of Pinctada fucata[J]. South China Fishe-ries Science,12(5):90-96.]
符韶,谢绍河,邓岳文,梁飞龙. 2012. 利用马氏珠母贝4个壳色系F3培育厚层优质珍珠[J]. 水产学报,36(9):1418-1424. [Fu S,Xie S H,Deng Y W,Liang F L. 2012. A study on thick pearls produced by the third generation shell color lines of pearl oyster(Pinctada martensii)[J]. Journal of Fisheries of China,36(9):1418-1424.]
何杰,吴旭干,赵恒亮,姜晓东,葛永春,王幼鹏,成永旭. 2016. 全程投喂配合饲料条件下池养中华绒螯蟹的生长性能及其性腺发育[J]. 中国水产科学,23(3):606-618. [He J,Wu X G,Zhao H L,Jiang X D,Ge Y C,Wang Y P,Cheng Y X. 2016. Growth performance and gonadal development of pond-reared Chinese mitten crab(Eriocheir sinensis) fed formulated diets during the whole culture process[J]. Journal of Fishery Sciences of China,23(3):606-618.]
黄建华,杨其彬,马之明,陈旭,周发林,温伟庚,江世贵. 2013. 池塘养殖斑节对虾的生长、发育与性成熟[J]. 水产学报,37(3):397-406. [Huang J H,Yang Q B,Ma Z M,Chen X,Zhou F L,Wen W G,Jiang S G. 2013. The growth,development and sexual maturity of pond-reared Penaeus monodon[J]. Journal of Fisheries of China,37(3):397-406.]
刘志刚,王辉,孙小真,杜晓东,李雷斌. 2007. 马氏珠母贝经济性状对体重决定效应分析[J]. 广东海洋大学学报,27(4):15-20. [Liu Z G,Wang H,Sun X Z,Du X D,Li L B. 2007. Analysis of determination effects of economic traits on body weight of Pinctada martensii(Dunker)[J]. Journal of Guangdong Ocean University,27(4):15-20.]
马文阁. 2016. 黄颡鱼GH/IGF生长轴基因的序列特征和两性表达差异分析[D]. 武汉:华中农业大学. [Ma W G. 2016. Characterization and sex-dependent expression of GH/IGF axis genes in yellow catfish[D]. Wuhan:Huazhong Agricultural University.]
马细兰,张勇,周立斌,刘晓春,林浩然. 2009. 脊椎动物雌雄生长差异的研究进展[J]. 动物学杂志,44(2):141-146. [Ma X L,Zhang Y,Zhou L B,Liu X C,Lin H R. 2009. Studies of growth sexual dimorphism in vertebrate[J]. Chinese Journal of Zoology,44(2):141-146.]
牛志凯,刘宝锁,张东玲,谭才钢,张博,陈明强,范嗣刚,姜松,黄桂菊,李有宁,喻达辉. 2015. 合浦珠母贝3个地理群体杂交后代生长性状和闭壳肌拉力的比较分析[J]. 南方水产科学,11(1):26-32. [Niu Z K,Liu B S,Zhang D L,Tan C G,Zhang B,Chen M Q,Fan S G,Jiang S,Huang G J,Li Y N,Yu D H. 2015. Comparative analysis of growth traits and shell-closing strength among hybrid populations from three geographical groups of pearl oyster (Pinctada fucata)[J]. South China Fisheries Science,11(1):26-32.]
石耀华,王爱民,吴星. 2004. 中国养殖马氏珠母贝多毛类寄生虫病的调查[J]. 海洋科学,28(7):13-18. [Shi Y H,Wang A M,Wu X. 2004. Survey on polychaete infestation of farmed pearl oyster,Pinctada martensii in China[J]. Marine Science,28(7):13-18.]
王愛民,石耀华,吴星. 2004. 4种防治马氏珠母贝多毛类寄生病方法的效果比较[J]. 海洋水产研究,25(2):41-46. [Wang A M,Shi Y H,Wu X. 2004. The comparison among effects of four treatments to reduce polychaete infestation in pearl oyster,Pinctada martensii[J]. Marine Fisheries Research,25(2):41-46.]
王爱民,阎冰,叶力,兰国宝. 2003. 马氏珠母贝不同地理种群内自繁和种群间杂交子一代感染多毛类寄生病的分析[J]. 海洋水产研究,24(3):38-44. [Wang A M,Yan B,Ye L,Lan G B. 2003. Analyses on polychaetes infestation of the first filial generation from matings and cro-sses of different geographical populations in Pinctada martensii(Dunker)[J]. Marine Fisheries Research,24(3):38-44.]
王冲. 2013. 栉孔扇贝不同性别间重要经济性状比较及通径分析[J]. 水产科学,32(8):441-446. [Wang C. 2013. Comparison and path analysis of important economic traits between genders of scallop Chlamys farreri[J]. Fisheries Science,32(8):441-446.]
王庆恒,邓岳文,杜晓东,符韶. 2008. 马氏珠母贝4个壳色选系F1幼虫的生长比较[J]. 中国水产科学,15(3):488-492. [Wang Q H,Deng Y W,Du X D,Fu S. 2008. Growth comparison of F1 larvae between four shell color lines of pearl oyster Pinctada martensii[J]. Journal of Fishery Sciences of China,15(3):488-492.]
王新安,马爱军,黄智慧,马得友,王婷,田岳强,王广宁,曲江波. 2014. 大菱鲆选育家系雌、雄群体的生长发育差异[J]. 水产学报, 38(4):464-469. [Wang X A,Ma A J,Huang Z H,Ma D Y,Wang T,Tian Y Q,Wang G N,Qu J B. 2014. Developmental differences between female and male groups in turbot(Scophthalmus maximus) bree-ding families[J]. Journal of Fisheries of China,38(4):464-469.]
王祎哲,邓正华,王雨,李有宁,于刚,黄桂菊,陈明强. 2018. 合浦珠母贝不同壳色选育系F6数量性状的相关性和通径分析[J]. 中国水产科学,25(5):988-997. [Wang Y Z,Deng Z H,Wang Y,Li Y N,Yu G,Huang G J,Chen M Q. 2018. Correlation and path analyses of quantitative traits in different shell color F6 lines of pearl oyster (Pinctada fucata)[J]. Journal of Fishery Sciences of China,25(5):988-997.]
邬思荣. 2011. 湛江北部湾深水海域马氏珠母贝四种壳色选育系F5的生长速度、生长模型及其遗传多样性的SSR分析[D]. 湛江:广东海洋大学. [Wu S R. 2011. Growth rate and growth model of F5 selected lines for four colors of Pinctada martensii (Dunker) cultured in deepwater area of Beibu bay in Zhanjiang and their genetic diversity analysis using SSR molecular markers[D]. Zhanjiang:Guangdong Ocean University.]
吴波,叶满,陈娈娈,邓思平,朱春华,师尚丽,李广丽. 2013. 不同性别养殖金钱鱼生长性能及消化酶活性的比较[J]. 上海海洋大学学报,22(4):545-551. [Wu B,Ye M,Chen L L,Deng S P,Zhu C H,Shi S L,Li G L. 2013. Growth performance and digestive enzyme activity between male and female Scatophagus argus[J]. Journal of Shanghai Ocean University,22(4):545-551.]
肖述,符政君,喻子牛. 2011. 香港巨牡蛎雌雄群体的数量性状通径分析[J]. 南方水产科学,7(4):1-9. [Xiao S,Fu Z J,Yu Z N. 2011. Path analysis of quantitative traits of male and females Hong Kong oyster Crassostrea hongkonggensis[J]. South China Fisheries Science,7(4):1-9.]
张文府. 2013. 马氏珠母贝四种壳色选育系中育珠贝家系和小片贝家系的选育研究[D]. 湛江:广东海洋大学. [Zhang W F. 2013. Study on the families selection of the four shell colors Pinctada martensii as culturing pearl and mantle tissue donors[D]. Zhanjiang:Guangdong Ocean University.]
赵永超. 2013. 三角帆蚌雌雄间生长和产珠性能差异研究[D]. 上海:上海海洋大学. [Zhao Y C. 2013. Comparison of growth and pearl production between female and male Hyriopsis cumingii[D]. Shanghai:Shanghai Ocean University.]
郑光明,马丽莎,朱新平,刘毅辉,陈永乐,罗建仁. 2008. 平胸龟年龄与生长及雌雄差异的比较[J]. 华南农业大学学报,29(3):66-70. [Zheng G M,Ma L S,Zhu X P,Liu Y H,Chen Y L,Luo J R. 2008. Age and growth of Platystemon megacephalum (Gay) and difference in growth and morphology between males and females[J]. Journal of South China Agricultural University,29(3):66-70.]
朱晓闻,刘志刚,王辉,邬思荣. 2012. 马氏珠母贝(Pinctada martensii)四种壳色选育系F5的生长及遗传多样性分析[J]. 海洋与湖沼,43(1):160-165. [Zhu X W,Liu Z G,Wang H,Wu S R. 2012. Growth and genetic diversity for four shell color lines(F5) of the pearl oyster Pinctada martensii[J]. Oceanologia et Limnologia Sinica,43(1):160-165.]
朱新平,陳永乐,魏成清,刘毅辉. 2003. 黄喉拟水龟雌、雄生长及形态差异[J]. 中国水产科学,10(5):434-436. [Zhu X P,Chen Y L,Wei C Q,Liu Y H. 2003. Diversity of male and female Mauremys mutica in growth and morphology[J]. Journal of Fishery Science of China,10(5):434-436.]
邹柯姝,张殿昌,郭华阳,张楠,江世贵. 2015. 4种壳色合浦珠母贝贝壳棱柱层和珍珠质层7种金属元素质量分数的比较分析[J]. 南方水产科学,11(3):74-79. [Zou K S,Zhang D C,Guo H Y,Zhang N,Jiang S G. 2015. Comparison of seven metallic elements in prismatic layer and na-creous layer of shells from four shell color strains in pearl oyster(Pinctada fucata)[J]. South China Fisheries Scien-ce,11(3):74-79.]
Aoki H,Ishikawa T,Fujiwara T,Atsurni T,Nishikawa H,Okamoto C,Komaru A. 2010. Utility of shell closing strength as the indicator of good health in breeding and culture management of Japanese pearl oyster Pinctada fucata[J]. Aquaculture,308(S1):S115-S118.
Browdy C L. 1998. Recent developments in penaeid broodstock and seed production technologies:Improving the outlook for superior captive stocks[J]. Aquaculture,164(1-4):3-21.
Ishikawa T,Okamoto C,Hayash M,Aoki H,Isowa K,Komaru A. 2009. The heritability of the shell-closing strength in the Japanese pearl oyster,Pinctada fucata martensii[J]. Aquaculture Science,57(1):77-82.
Iwai T,Takahashi M,Ido A,Miura C,Miura T. 2015. Effect of gender on Akoya pearl quality[J]. Aquaculture,437:333-338.
Okamoto C,Komaru A,Hayashi M,Isowa K. 2006. Shell-closing strength of pearl oyster,Pinctada fucata martensii and its relationship to mortality and various parts weight of oyster[J]. Aquaculture Science,54(3):293-299.
Poulet S A,Lennon J F,Plouvebez F,Jalabert F,Correc G,Cueff A,Lacoste A. 2003. A nondestructive tool for the measurement of muscle strength in juvenile oysters Cra-ssostrea gigas[J]. Aquaculture,217(1-4):49-60.
Zou K S,Zhang D C,Guo H Y,Zhu C Y,Li M,Jiang S G. 2014. A preliminary study for identification of candidate AFLP markers under artificial selection for shell color in pearl oyster Pinctada fucata[J]. Gene,542(1):8-15.
