可溶性高尿酸对人脐静脉内皮细胞NLRP3炎性体途径的影响
2020-03-30苏毕阿吉尹洪超
苏毕·阿吉 尹洪超
尿酸(uric acid,UA)作为体内嘌呤代谢产物,是血浆中主要的抗氧化物之一,能有效抵抗氧自由基导致的氧化应激[1]。然而在人类进化中,由于丢失了尿酸酶这一基因,导致人体血液中的UA比其他哺乳动物体内高出10倍左右。当人体血尿UA浓度高于6.8mg/dl时被定义为高尿酸血症。近年来的流行病研究结果显示,高尿酸血症与多种心血管疾病的病理过程相关,如高血压、冠心病、动脉粥样硬化等,然而UA在其中的病理生理学作用仍有待于进一步研究[2~4]。内皮细胞作为血管的第1层屏障和内分泌细胞,通过旁分泌作用分泌多种血管活性物质和炎性细胞因子在调节血管功能方面起着重要作用[5]。内皮细胞活化被Pober等[6]研究者定义为,内皮细胞在损伤因素的刺激下改变了蛋白质的合成,并以细胞因子的形式传递细胞对刺激的反应。当内皮细胞发生活化时,选择素、黏附分子、趋化分子等炎性介质表达增加,从而参与心血管等疾病的使动环节[7]。因此,内皮细胞在心血管疾病发生、发展中起重要作用。
NOD样受体(NOD-like receptor,NLR)家族蛋白3(NLRP3)炎性体是由NLRP3、凋亡相关微粒蛋白(apoptosis-associated speck-like protein containing CARD,ASC)和胱冬肽酶-1前体(pro-caspase-1)组成的异源三聚体。被激活时,NLRP3蛋白募集衔接蛋白ASC,ASC募集下游失活状态的pro-caspase-1[8]。该炎性体复合物的聚合会使pro-caspase-1自我催化形成活化状态的caspase-1。活化的caspase-1切割游离的IL-1β前体(pro-IL-1β)形成成熟的IL-1β并释放于胞外。普遍认为,NLRP3炎性体通路需要两步激活[9]。第一步需要炎性体启动(inflammasome priming)信号,负责诱导NLRP3和pro-IL-β的转录翻译,使其表达量上调,为转录水平调控;第二步为炎性体活化(inflammasome activation)信号,负责诱导NLRP3炎性体复合物的募集,如形成NLRP3-ASC复合物等,为转录后水平调控。多种配体可作为NLRP3炎性体的启动信号刺激物,包括作用于Toll样受体4(TLR4)的脂多糖(LPS)和作用于TLR2的Pam3CSK4均激活NF-κB通路从而促进NLRP3和pro-IL-1β的转录[10]。多种刺激物可作为NLRP3的活化信号,包括UA晶体、ATP等,他们共同的作用机制可能通过ROS生成、溶酶体降解或阳离子外流等途径[11~13]。
NLRP3炎性体是固有免疫的重要组成部分,其主要作用是识别非微生物危险信号并诱发无菌性炎性反应参与各种炎性慢性疾病的发生、发展[14]。近期许多研究结果显示,UA作为一种内源性危险信号,不仅单钠尿酸盐(MSU)能激活NLRP3炎性体,溶解性UA也能激活NLRP3,如在小鼠骨髓源巨噬细胞、大鼠心肌细胞、人单核细胞、平滑肌细胞和肾小管上皮细胞等[15~19]。目前并不了解溶解性UA是否能激活人内皮细胞NLRP3炎性体通路并影响其功能。已有研究报道,200μg/ml的可溶性高浓度UA作用于内皮细胞,并通过HMGB1/RAGE通路诱导内皮细胞功能紊乱[20]。且200μg/ml以上的UA在低温下可能会析出[21]。因此,本研究用高浓度(200μg/ml)可溶性UA明确高浓度UA是否对HUVECs NLRP3炎性体通路有显著影响。
材料与方法
1.材料:原代人脐静脉内皮细胞(human umbilical vein endothelial cell, HUVEC)和血管内皮细胞培养基(endothelial cell medium, ECM)购自美国ScienCell公司;尿酸(uric acid, UA)购自美国Sigma公司;TB GreenTMPremix Ex TaqTMⅡ(RR820A)购自宝日医生物技术(北京)有限公司;蛋白质定量试剂盒(BCA法)(P1511)、抗β-tubulin单克隆抗体(C1340)、HRP标羊抗小鼠二抗(C1308)、HRP标羊抗兔二抗(C1309)和活性氧检测试剂盒(S0033)购自碧云天生物技术公司;NLRP3一抗(ab214185,ab4207)、IL-1β一抗(ab9722)、ASC一抗(ab155970)、羊抗兔荧光标记IgG(ab96899)和驴抗羊荧光标记IgG(ab150131)购自英国Abcam公司;P65一抗(D14E12)购自美国CST公司;TRIzol(15596026)和驴抗兔荧光标记IgG(A21206)购自美国赛默飞公司。
2.细胞培养方法:HUVECs用含5%胎牛血清、1%内皮细胞生长添加剂 (ECGS,Cat.No.1052)的ECM培养基于37℃、5%CO2孵箱中常规培养。细胞隔天换液,融合至80%~90%时胰酶按1∶3传代,取4~7代细胞用于后续实验。
3.UA溶液配制:参考如前所述的方法配制200μg/ml的UA溶液[21]:将称取好的UA粉末直接添加至ECM,37℃孵育40min,并每隔10min上下颠倒混匀。待UA粉末完全溶解,用0.22μm滤器过滤至无菌容器中。尿酸酶法检测UA浓度,并根据检测结果进行矫正,再次确认浓度,4℃保存,并1周内用完。
4.RNA提取和qPCR检测:取第4~5 代细胞,前1天将细胞传代至6cm皿,次日更换200μg/ml的UA培养液并培养24h。用TRIzol(1毫升/皿)收集各组细胞,在预处理过的无RNase环境抽提细胞RNA。用微量分光光度计测RNA浓度,并按RT-PCR试剂盒(北京全式金生物技术有限公司,AT311)流程进行反转录。根据qPCR试剂盒(RR820A)说明进行qPCR反应。β-actin作内参,以2-ΔΔCt值计算各基因的相对表达量。PCR反应引物序列及扩增片段长度见表1。

表1 PCR反应引物序列及扩增片段长度
5.Western blot法检测蛋白表达:细胞处理同qPCR。其中LPS(500ng/ml,12h)作阳性对照。用Western blot法及IP细胞裂解液(碧云天生物技术公司,P0013)提取细胞总蛋白,并用BCA法测定蛋白浓度。调整蛋白浓度后95℃金属浴10min,并取10μg蛋白进行SDS-PAGE胶电泳,将蛋白电泳产物转印至NC膜上。用5% BSA室温封闭1h,分别加入β-tubulin一抗(1∶5000)、NLRP3一抗(1∶200)及IL-1β一抗(1∶1000),4℃孵育过夜。次日,TBST洗5min×4次,加入相应二抗(1∶5000),室温孵育1h。TBST洗5min×8次,ECL发光、显影。采用Image J图像处理软件分析各蛋白灰度值。以tubulin为内参,目的蛋白灰度值/内参蛋白灰度值算出目的蛋白相对表达量。
6.免疫荧光染色:将HUVEC以密度1.5×105接种于24孔板爬片过夜培养,并用200μg/ml UA处理细胞24h或500ng/ml LPS处理12h(阳性对照)。4%多聚甲醛室温固定15min,PBS洗3次,含0.3% Triton X-100的5%BSA室温封闭60min。弃上清加入P65一抗(1∶400)4℃过夜。次日,PBS洗3次加入羊抗兔二抗(1∶250)室温避光孵育1h,PBS洗3次,含DAPI甘油封片,并用共聚焦显微镜拍照。至于NLRP3和ASC双染实验采用依次孵育的方法,即先孵育NLRP3一抗(1∶200)4℃过夜,之后室温避光孵育1h驴抗羊红色荧光二抗(1∶500),再孵育ASC一抗(1∶250)4℃避光过夜,最后室温避光孵育1h驴抗兔绿色荧光二抗(1∶500),含DAPI甘油封片并拍照。采用对照组调整背景荧光强度后固定条件,每个实验组拍摄5~10个视野。
7.ROS检测:采用DCFH-DA法。细胞处理同免疫荧光染色。各组在培养6h时分别加入1∶1000稀释的 DCFH-DA(终浓度10μmol/L),继续37℃避光孵育45min。遇冷PBS洗3次,1%多聚甲醛室温固定15min,PBS洗3次,含DAPI甘油封片,及时用共聚焦显微镜观察并拍照,并采用Image J图像处理软件统计各组照片中平均细胞光密度(IOD)值。
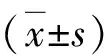
结 果
1.UA诱导表达HUVECs黏附分子和趋化因子mRNA:200μg/ml的UA诱导HUVECs的黏附分子ICAM-1和E-选择素mRNA水平上调(图1中A、B)。其中ICAM-1表达量上调约5倍(P=0.000),E-选择素表达量上调约2倍(P<0.01)。此外,200μg/ml的UA诱导HUVECs的趋化因子CXCL10和CCL26 mRNA表达量上调(图1中C、D),且CXCL10和CCL26 mRNA均上调5倍左右(P=0.000,P<0.01)。
2.UA降低HUVECs内皮型一氧化氮合酶(eNOS)mRNA水平:由图2可见,与对照组比较,200μg/ml UA组eNOS mRNA表达量下调约2.4倍,提示溶解性高UA下调eNOS表达量。
3.UA对HUVECs NLRP3炎性体核酸和蛋白水平的影响:用LPS作阳性对照显示,细胞NLRP3和pro-IL-1β蛋白水平均显著上调(图3中C、D),且切割后成熟IL-1β水平也增加到约2倍。然而,200μg/ml的可溶性UA对 HUVECs内NLRP3、pro-IL-1β的mRNA(图3中A、B)和蛋白水平没有显著影响(图3中C、D),且未切割形成成熟的IL-1β(图3中C、D)。可溶性高UA对HUVECs NLRP3和pro-IL-β转录水平无显著影响。由图4可见,200μg/ml UA刺激HUVECs 24h后,未检测到NF-κB核移位。而阳性组LPS明显上调了HUVECs的NF-κB表达并激活入核。可溶性高UA对HUVECs NF-κB通路无明显的刺激作用。
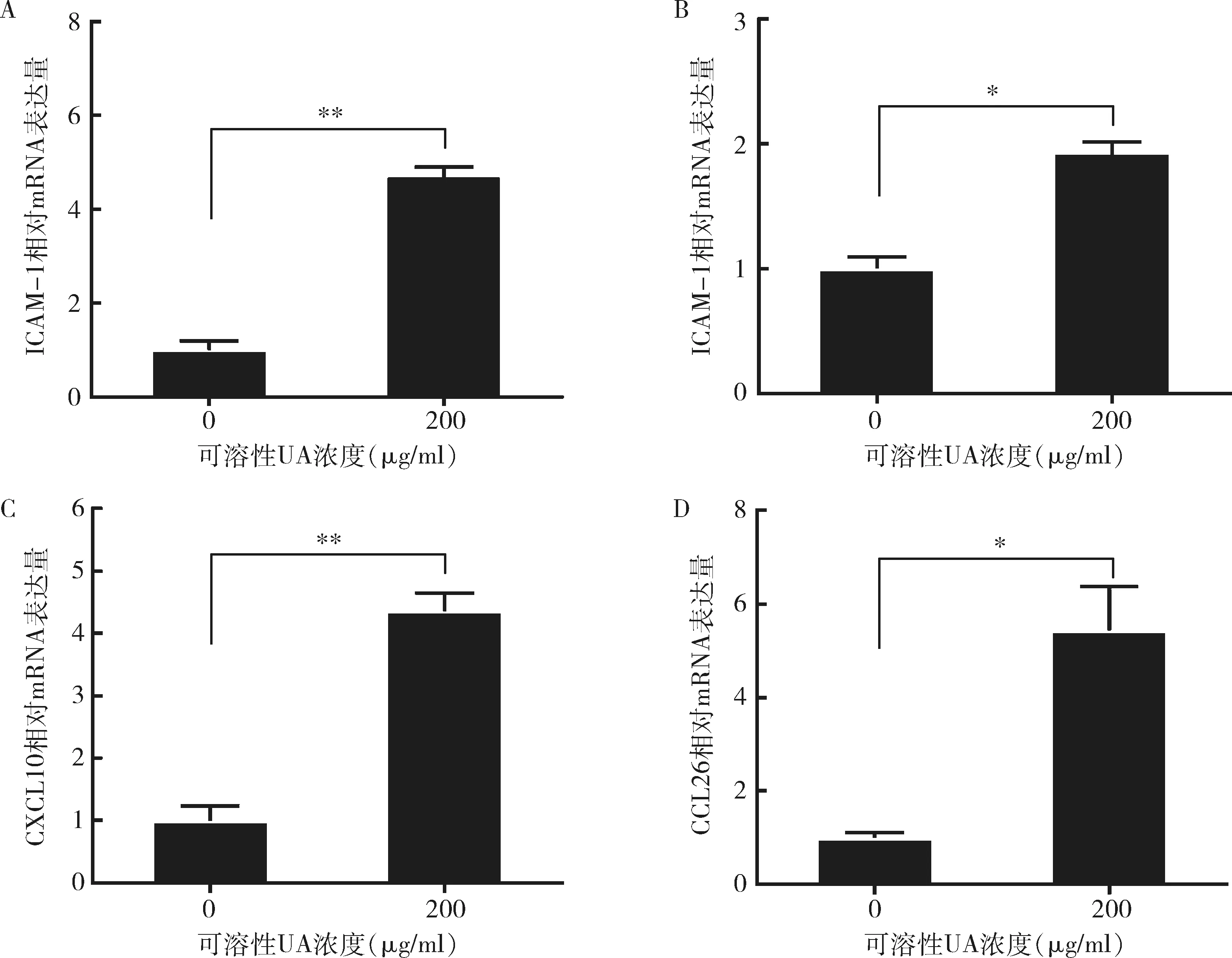
图1 UA诱导HUVECs黏附分子和趋化因子mRNA表达用200μg/ml的可溶性UA刺激HUVEC 24h,并通过qPCR检测黏附分子ICAM-1、E-选择素和趋化因子CXCL10、CCL26 mRNA水平。数据来自3个独立实验,*P<0.01, **P=0.000

图2 qPCR检测eNOS mRNA表达量用200μg/ml的可溶性UA刺激HUVEC 24h,并通过qPCR检测eNOS mRNA水平。数据来自3个独立实验,*P<0.01
4.UA诱导HUVECs内NLRP3-ASC复合体形成:200μg/ml UA诱导NLRP3-ASC共定位,可溶性高UA促进了HUVECs 内NLRP3-ASC复合体形成(图5)。
5.UA上调HUVECs ROS水平:HUVECs胞质ROS水平显著增高(图6中A、B,P=0.000)。
讨 论
心血管疾病是世界上高发生率和高病死率的疾病之一。大规模的流行病学研究提示,高尿酸血症与心血管疾病的病程进展密切相关。很多研究者倾向于认为,UA可能是心血管疾病发生率和病死率独立的特异性标志物之一[2]。然而其相关机制尚需进一步研究。NLRP3炎性体作为固有免疫的重要组成部分,识别非微生物危险信号并诱发无菌性炎性反应参与心血管疾病等慢性炎性疾病的发生、发展[14]。因此,笔者通过高浓度的UA体外处理HUVECs,探究可溶性高UA是否诱导HUVECs NLRP3炎性体通路来参与这一过程。

图3 UA对HUVECs NLRP3炎性体启动无显著影响A、B.200μg/ml的可溶性UA刺激HUVECs 24h或500ng/ml LPS处理12h后 Western blot法检测NLRP3、pro-IL-1β和IL-1β的蛋白表达量;C.用qPCR检测NLRP3相应mRNA水平;D.用qPCR检测IL-1β相应mRNA水平。数据来自3个独立实验,*P<0.01

图4 UA对HUVECs NF-κB无明显刺激作用200μg/ml UA刺激HUVECs 24h或500ng/ml LPS处理12h后,共聚焦显微镜观察NF-κB的表达和核移位情况(免疫荧光染色)
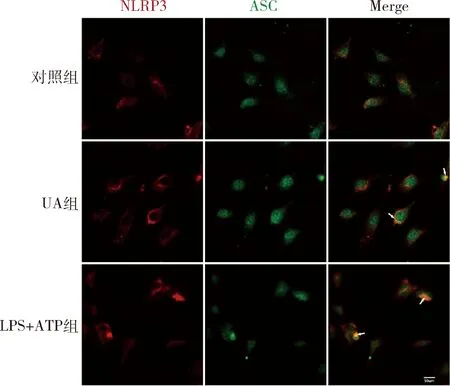
图5 UA诱导HUVECs 内NLRP3-ASC募集200μg/ml UA刺激HUVECs 24h或500ng/ml LPS处理12h+5mmol/L ATP处理30min后,共聚焦显微镜检测NLRP3和ASC共定位情况(免疫荧光染色)

图6 UA诱导HUVECs 产生ROSA.200μg/ml的可溶UA处理HUVEC 24h后,用DCFH-DA荧光探针检测胞质ROS水平;B.通过ImageJ图像处理软件统计ROS相对荧光强度。柱状图代表至少5个单独视野中单个细胞总荧光强度(免疫荧光染色)
eNOS通过产生一氧化氮(NO)发挥舒血管作用,在内皮细胞功能调节中起标志性重要作用[22]。eNOS的转录水平调控和转录后调控以及蛋白之间的相互作用均会影响NO的合成和释放,从而影响内皮细胞功能。由于NLRP3炎性体启动过程需要NF-κB通路的参与,因此笔者用UA处理HUVECs之后荧光标记NF-κB确定UA是否能激活NF-κB途径[10]。ASC作为NLRP3通路的衔接分子,一旦与NLRP3产生复合体,募集下游的效应器分子并最终切割pro-IL-1β形成有生物学活性的IL-1β。因此,NLRP3-ASC共定位可作为NLRP3炎性体活化的标志。本研究结果显示,可溶性高UA诱导HUVECs上调黏附分子ICAM-1、E-选择素和趋化分子CXCL10、CCL-26,提示UA诱导HUVECs活化。同时可溶性高UA下调HUVECs eNOS mRNA水平,提示UA在转录水平影响eNOS表达,从而可能影响HUVECs NO合成功能。
QPCR和Western blot法检测结果显示,可溶性高UA对NLRP3和pro-IL-1β mRNA和蛋白水平均无显著影响,提示可溶性高UA并未激活HUVECs NLRP3炎性体的启动信号。虽然UA诱导HUVECs形成了NLRP3-ASC复合物,但可能由于NLRP3炎性体复合物形成较少且细胞内没有上调的pro-IL-1β,并未在细胞水平检测到切割成熟的IL-1β。已有研究报道用LPS+UA的共刺激作用于小鼠骨髓源巨噬细胞等检测到IL-1β的释放[15,23]。因此,UA与其他危险因素如高脂、高糖等共同存在的条件下,UA与这些因子是否有协同作用激活NLRP3炎性体途径并加剧内皮细胞功能不良有待做深入研究。
本研究结果显示,可溶性高UA未诱导HUVECs NF-κB活化。有多项研究报道,在人外周血单核细胞(PBMCs),UA通过NF-κB通路诱导NLRP3炎性体启动,显著上调 NLRP3和pro-IL-1β,为NLRP3炎性体活化提供分子基础[10]。但本实验结果提示,UA在HUVECs不通过激活NF-κB通路诱导NLRP3和pro-IL-1β的转录表达。
本实验也观察到UA诱导ROS在HUVECs内累积。UA被认为具有抗氧化性,但之后被报道UA的(抗)氧化能力取决于它所处的环境,UA在细胞外基质和血浆中主要承担抗氧化作用,一旦进入细胞就发挥促氧化作用[1]。关于HUVECs的一项研究表明,高UA能正反馈进入细胞中,并通过下调UA外流通道抑制UA流出,导致UA在内皮细胞内大量积累并产生ROS[24]。另外,曾在单核细胞、人THP-1细胞等其他细胞系中多次被报道ROS可激活NLRP3炎性体[13]。但在本实验中,ROS对HUVECs 内IL-1β的产生没有显著影响,提示ROS在HUVECs对NLRP3炎性体途径的激活无直接刺激作用。
综上所述,本研究首次用原代人脐静脉内皮细胞探讨了UA对NLRP3炎性体通路的作用。结果显示,可溶性高UA能使HUVECs活化。虽然UA促进NLRP3-ASC复合体形成,但没有诱导IL-1β的产生。虽然UA上调HUVECs ROS水平,但跟NLRP3通路的激活不直接相关。因此可溶性高UA不激活HUVECs NLRP3炎性体途径从而影响HUVECs相关功能。实验结果也显示,细胞内NLRP3-ASC复合物显著增加,因此不能排除UA在其他因素协同作用下诱导NLRP3炎性体的激活并促进内皮细胞功能紊乱,从而参与心血管疾病等慢性炎性疾病的发生、发展。
