套尔河贝类增养殖区放流活动前后浮游植物群落结构变化
2020-03-26吕其明张晶晶高彦洁吕振波王田田于君宝王庆孝
吕其明,张晶晶,高彦洁,吕振波,王田田,于君宝,王庆孝
(1.鲁东大学滨海生态高等研究院,山东省高校重点实验室“滨海湿地保育与生态修复重点实验室”,山东烟台 264025;2.烟台市水产研究所,山东烟台 264003)
浮游植物作为海洋生态系统中最主要的初级生产者,也是重要的饵料生物,其群落结构和丰度会直接影响上层食物网[1]。贝类是重要的滤食性低营养级生物,浮游植物供给不足会导致贝类个体生长缓慢、发病率高。此外,贝类对浮游植物饵料具有选择性,偏好摄食含有特种脂肪酸的浮游植物,不同大小和种类的贝类摄食浮游植物偏好也不同,因此浮游植物群落结构会直接影响贝类的生长状态[2]。环境因子是影响浮游植物群落结构和丰度的重要因素,诸如温度、盐度、光照及营养盐浓度等因素都会强烈影响浮游植物群落结构[3-4]。贝类增养殖区环境因子不仅受自然条件(例如降雨、淡水输入、海流)的影响,还受到贝类增养殖活动的影响[5]。适宜的贝类放流密度有利于降低水体营养盐浓度,改善近海富营养化现象[6];过高的放流密度则会形成大量的富含营养物质的浅层沉积碎屑,这些富含氨基酸和有机氮的碎屑会随着水流再次输送至水体,对浅海生态环境造成强烈的影响[7]。因此,系统研究贝类增养殖区环境因子的变化特征及其对浮游植物群落结构分布的影响,是确定合适的贝类增养殖种类和增养殖密度、维持贝类增养殖区域生态环境可持续发展的前提。
黄河口邻近海域是典型的海陆交互作用地带,受河流输入影响,环境季节性变化明显,生态系统功能复杂,同时该区域海岸线长,滩涂面积广阔,海水营养丰富、饵料充足,贝类资源丰富[8]。套尔河河口位于黄河口西侧,是重要的贝类产区,同时,该海域也是经济发展的重点区域,港口工程的建设、人类对生物资源的掠夺性开发、不经处理排放废弃物及不科学的渔业养殖等,造成该海区生物资源尤其是贝类资源不断衰减[9]。为了恢复贝类资源,2006年以来,山东省加强了黄河口附近海域的贝类增殖放流活动,增殖主要以文蛤(Mercenariamercenaria)、杂色蛤(Venerupisvariegate)、竹蛏(Solenstrictus)、泥螺(Bullactaexarate)等地方种类为主,本区域主要放流品种是文蛤(规格6 000粒·kg-1),且投放量逐年增多,2015年4 000万粒,2016年6 000万粒,2017年放流2亿粒,2018年5月8日进行贝类放流活动,投放量达4亿粒。对恢复及增加贝类种群生物量、改善水质取得一定效果[10]。但是,目前针对该海域增养殖前后环境因子变化特征及其对浮游植物群落结构影响的研究较少,贝类增养殖活动缺乏理论依据。
本研究通过对2018年5月、8月黄河口西侧套尔河贝类增养殖海区贝类放流前后环境因子、叶绿素a和浮游植物群落结构进行系统调查,分析了增养殖区环境因子、叶绿素a和浮游植物群落的空间分布特征,并结合皮尔森(Person)相关分析和冗余分析(RDA),研究了放流前后叶绿素a、浮游植物群落结构与环境因子之间的相关关系,旨在揭示贝类增养殖区影响浮游植物群落结构及丰度的关键环境因子及其响应关系,为深入分析贝类增养殖对河口地带生态环境的影响提供重要的基础资料。
1 材料与方法
1.1 采样站位
2018年5月(贝类增殖放流前)、8月(贝类增殖放流后),在套尔河贝类增养殖区附近设置20个站点(图1),分别开展了生态环境综合调查,取表层水样进行检测,项目包括温度、盐度、营养盐、叶绿素a;同时分别在河口、泥滩浅海交界处和离岸海域选取6个代表站位(A1、A8、A6、A13、A14、A19)采集浮游植物样品,鉴定其群落结构。
1.2 样品采集及分析
样品的采集分析和浮游植物鉴定严格按照《海洋调查规范》(GB/T 12763)进行,用5 L的卡盖式采水器采集表层水样分别用于营养盐、叶绿素a和浮游植物分析。取500 mL海水装入聚乙烯瓶,并立即加入甲醛溶液固定保存(终浓度为3% ~5%),取 300 mL水样经 GF/F滤膜(Whatman)过滤后,滤膜于-20℃下避光并冷冻保存,用于叶绿素a浓度的测定,采用分光光度法进行叶绿素a的测定分析。每个采样点取3个平行样,结果取其平均值。另取250 mL水样用0.45μm醋酸纤维滤膜过滤,滤液用于营养盐浓度的测定。
浮游植物计数:浮游植物样品带回实验室,将水样摇匀,取200 mL放入分液漏斗,静置24 h,将最下面1~2 mL浓缩液体接入小瓶用于浮游植物计数,在OLYMPUSCKX 53光学显微镜下(10×40倍)进行浮游植物的计数,取0.1 mL样品放入计数框中,观察10个计数格,每个样品计数2片,取其平均值做最终结果,若2片计数结果相差超过15%,则进行第3片计数,并取其中个数相近的2片的平均值;换算成浮游植物密度即每升水样中藻类的细胞个数,单位为个·L-1。
温度、盐度与水深使用船载 CTD(Seabird 911)现场测定。无机氮(DIN,为 NO3-N、NO2-N与NH4-N之和)、磷酸盐(DIP)使用营养盐自动分析仪(QuAAtro AutoAnalyzer 39)进行测定,其中,硝酸盐(NO3-N)采用镉-铜还原后再用盐酸-萘乙二胺比色法测定;亚硝酸盐(NO2-N)采用盐酸-萘乙二胺比色法测定;氨氮(NH4-N)采用水杨酸钠法测定;磷酸盐(PO4-P)采用磷钼蓝法。NO3-N、NO2-N、NH4-N和PO4-P的检出限分别是0.000 3、0.000 1、0.000 5和 0.000 2 mg·L-1。

图1 2018年5月、8月采样站位Fig.1 Sampling sites in May and August 2018
1.3 数据处理
通过应用物种优势度、香农-威纳指数、单纯度、均匀度及丰富度指数对浮游植物群落结构进行分析。公式如下:
通过物种优势度指数(Y)指示某物种在种群中的优势地位,当Y>0.02时,即认为该种为优势种[11]:

香 农-威 纳 指 数 (H, Shannon-Wiener index)[12]:
H=-∑Pilog2Pi
单纯度(C):
C=∑(Pi)2
均匀度(J)使用 Pielou指数[13]:
J=H/log2S
丰富度指数(d)使用 Margalef指数[14]:
d=(S-1)/log2N
以上各式中,ni为第i种的总个体数;N为所有站位上采集的生物总个体数;fi为物种在所有站位上出现频率;Pi为该站位上第i种个体数占总个体数的比率;S为各站种类数。
1.4 统计分析
环境因子数据与浮游植物生物量数据采用Excel 2010软件进行统计;数据的正态性检验、叶绿素a与环境因子的相关性使用SPSS 22.0进行分析;将浮游植物与环境因子数据进行lg(x+1)转换,对转换后的数据先进行除趋势对应分析(DCA),结果显示4个轴最大梯度长度小于3,因而选用线性模型的RDA分析环境因子和浮游植物群落结构的相关性。温度、盐度、营养盐、叶绿素a及浮游植物群落结构分布图采用ArcGIS 10.3软件进行绘制;浮游植物群落结构与环境因子的主成分分析和冗余分析的排序图使用Canoco for Windows 4.5软件绘制。
2 结果与分析
2.1 调查海域温度和盐度的时空分布
5月调查水域平均温度18.1℃,整体呈现近岸—离岸逐渐递减的趋势,高值区出现在套尔河河口,最高温度为20.5℃;低值区出现在离岸海域,温度为16.5℃。盐度呈现与温度类似的近岸—离岸逐渐递减趋势,平均盐度29.8,高值区在套尔河河口,盐度最高值为31.8;低值区出现在东部离岸海域,盐度为28.7。8月调查区域温度明显升高,平均温度为32.0℃,温度整体趋势与5月份基本一致,呈现近岸—离岸逐渐递减的趋势,高值区也出现在套尔河河口,最高温度为34.1℃;低值区出现在离岸海域,温度为30.3℃。8月盐度明显低于5月份,平均盐度28.1,盐度变化趋势与5月份相反,高值区出现在离岸海域,呈现明显的河口—离岸逐渐递增的趋势,高值区出现在离岸区域,盐度为29.9,低值区在套尔河河口,盐度为20.2。
2.2 调查海域营养盐的时空分布特征
5月调查海域营养盐整体呈现自河口至离岸逐渐递减的趋势(图3),高值区均出现在套尔河河口,硝酸盐的平均浓度是0.367 mg·L-1,变化范围为0.272~0.592 mg·L-1(图 3-A);亚硝酸盐的平均浓度是0.022 mg·L-1,变化范围为0.006~0.080 mg·L-1(图3-B);氨氮平均浓度是0.021 mg·L-1,变化范围为0.003~0.195 mg·L-1(图3-C);磷酸盐的平均浓度0.005 mg·L-1,变化范围为0.003~0.013 mg·L-1(图3-D)。根据浮游植物营养限制性法则,DIN、DIP浓度分别低于1μmol·L-1、0.1μmol·L-1时,对浮游植物生长造成绝对限制。5月所有调查站位均不存在氮限制,有3个站位(A3、A5和A14)DIP略低于 0.1μmol·L-1,存在绝对磷限制,N/P范围为43.6~126.6,平均值为86.1。
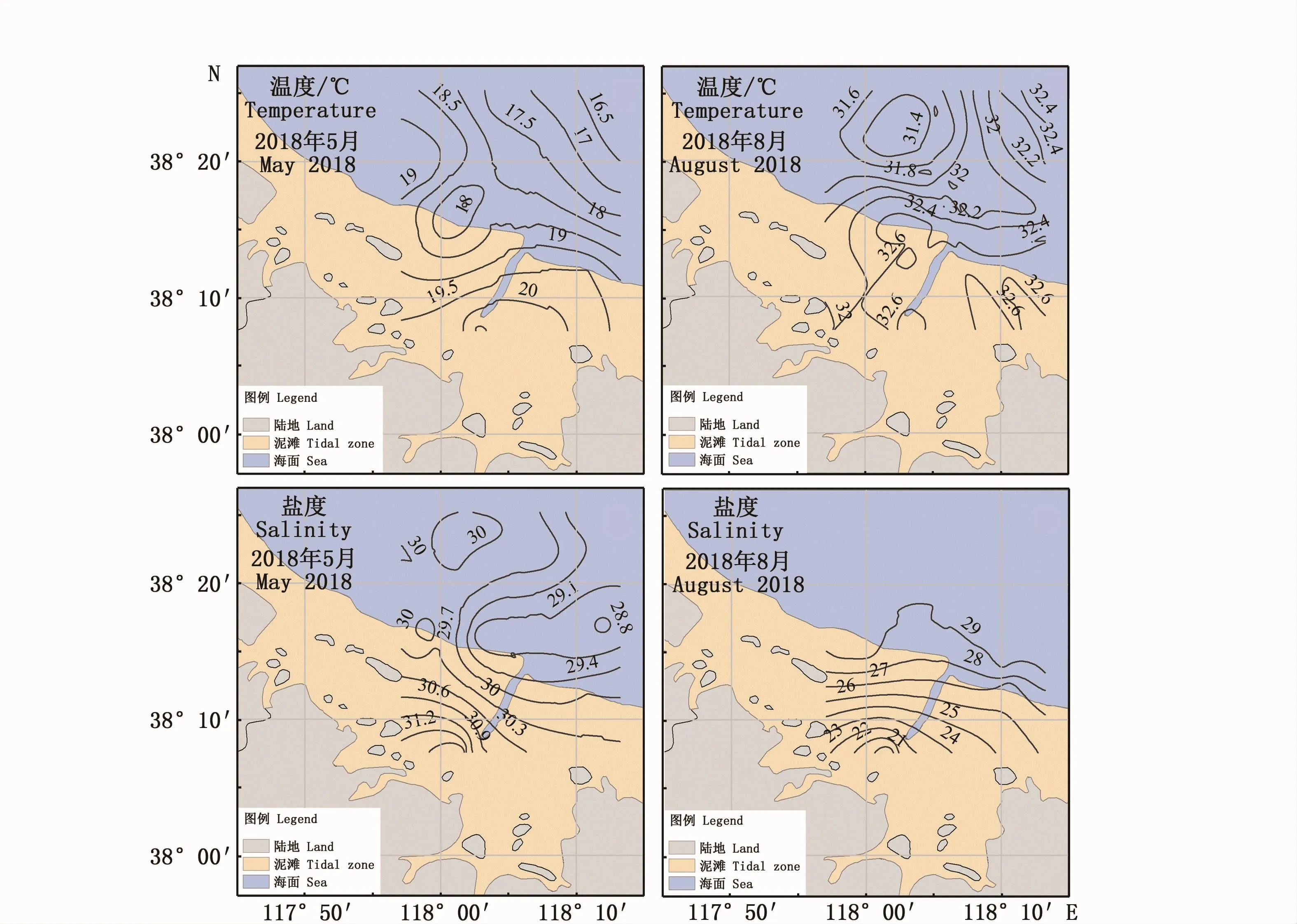
图2 调查海域温度、盐度时空分布图Fig.2 Spatial distribution of sea surface temperature and salinity during the surveys

图3 2018年5月调查海域营养盐分布图Fig.3 Spatial distribution of nutrient concentrations during the survey in May
8月,在整个调查区域硝酸盐、亚硝酸盐、氨氮整体趋势与5月份相似(图4-A~C),高值区均出现在套尔河河口区域,说明陆源排放依然是调查海域氮营养盐的主要来源。8月,硝酸盐的平均浓度是0.134 mg·L-1,变化范围为0.015~0.419 mg·L-1(图4-A);亚硝酸盐和氨氮的平均浓度分别为0.106 mg·L-1和0.043 mg·L-1,变化范围分别为0.016~0.670 mg·L-1和0.015~0.419 mg·L-1(图 4-B,C)。磷酸盐平均浓度为 0.004 mg·L-1(图 4-D),浓度范围为0.002~0.010 mg·L-1,磷酸盐空间分布与其他营养盐略有不同,高值出现在套尔河河口东侧潮间带、浅海交汇区。与5月份相比,可溶性无机氮(DIN)和磷酸盐(DIP)整体呈现降低的趋势。氮营养盐结构发生明显变化,硝酸盐的浓度明显降低,亚硝酸盐和氨氮的含量明显升高。根据浮游植物营养限制性法则,8月所有调查站位不存在氮限制,但存在磷限制的站位增多至5个(A3、A4、A7、A8和 A9),N/P范围为 14.5~331.5,平均值为62.8。
2.3 叶绿素a的时空分布特征及环境因子相关分析
5月调查水域径流量较小,叶绿素a的平均浓度为4.00μg·L-1,叶绿素a整体呈现近岸—离岸逐渐递减的趋势(图5),高值区出现在套尔河河口东侧,与营养盐高值区基本重合,叶绿素a浓度最高值为5.73μg·L-1;低值区出现在离岸海域,叶绿素a浓度为2.99μg·L-1。8月调查水域叶绿素a的浓度略高于5月,平均浓度为4.77μg·L-1;叶绿素a分布趋势与5月份有明显的不同,高值区出现在调查区域东侧离岸区域,两个高值出现在A10和A16站位,叶绿素a浓度分别为6.16和6.22μg·L-1,河口区域叶绿素a浓度相对偏低。
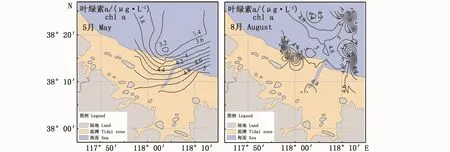
图5 2018年调查海域叶绿素a时空分布图Fig.5 Spatial distribution of chlorophyll a during the surveys
为了进一步研究环境因子对调查海域叶绿素a的影响,本文分别对5月、8月叶绿素a与环境因子进行了相关性分析。结果如表1所示,数据的正态性检验P值=0.2>0.05,表明数据服从正态分布,5月叶绿素a浓度与温度、盐度、氮营养盐浓度呈现正相关关系,与磷酸盐呈现负相关关系,显著性检验显示叶绿素a与氨氮和盐度的P值小于0.05,呈现显著的正相关关系。8月,叶绿素a浓度与营养盐浓度呈现正相关关系,与温度、盐度呈现负相关关系,但相关性不显著。
2.4 调查海域浮游植物群落结构时空分布特征
5月调查海域共采集到浮游植物3门29种,其中,硅藻23种,是主要浮游植物种类,占总物种数的79.3%;其次为甲藻门5种,金藻门1种。浮游植物生物量整体呈现河口近岸高、离岸低的特点,高值区出现在A13、A19站位,以布氏双尾藻 (Ditylumbrightwellii)、威 利 圆 筛 藻(Coscinodiscuswailesii)和劳氏角毛藻(Chaetoceros lorenzianus)为主(图6-A);低值区出现在离岸A8站位,夜光藻(Noctilucascientillans)和斯氏几内亚藻(Guinardiastriata)占比明显增加。调查海域优势种类(Y≥0.02)共4种,优势种为布氏双尾藻、威利圆筛藻、羽纹藻(Pinnulariasp.)和夜光藻,优势度分别为 0.43、0.10、0.04和 0.03。物种多样性指数变化范围为2.36~3.14,平均值为2.78;单纯度指数变化范围为0.15~0.35,平均为0.22;均匀度指数变化范围为0.58~0.85,平均为0.77,物种丰富度指数变化范围为0.54~0.83,平均为0.67(表2)。
8月调查海域共采集到浮游植物3门34种,其中硅藻29种,甲藻4种,金藻1种,硅藻依然是最主要的浮游植物种类,占物种总数的85.3%。浮游植物生物量变化趋势与5月份类似,高值区出现在潮间带与浅海交汇处,以旋链角毛藻(Chaetoceroscurvisetus)、 丹 麦 细 柱 藻(Leptocylindrusdanicus)和菱形海线藻(Thalassionemanitzschioides)为主,低值区出现在离岸海区,以菱形海线藻和旋链角毛藻为主(图6-B)。8月优势种更替明显,调查海域优势种共9种,分别是中肋骨条藻(Skeletonemacostatum)(Y=0.13)、旋链角毛藻(Y=0.11)、派格棍形藻(Bacillariapaxillifera)(Y=0.05)、丹麦细柱藻(Y=0.05)、海洋斜纹藻(Pleurosigmapelagicum)(Y=0.04)、伏氏海线藻(Thalassionemafrauenfeldii)(Y=0.03)、柔弱角毛藻(Chaetocerosdebilis)(Y=0.02)和尖刺伪菱形藻(Pseudonitzschiapungens)(Y=0.02)。物种多样性指数变化范围为0.96~5.44,平均值为2.77;单纯度指数变化范围为0.004~1.91,平均为0.70;均匀度指数变化范围为0.27~1.28,平均为0.67;物种丰富度指数变化范围为0.47~1.17,平均为0.84(表2)。

表1 叶绿素a与环境因子之间的相关性分析Tab.1 Correlation analysis between chlorophyll a and environmental factors

表2 浮游植物生物多样性统计Tab.2 Distribution of abundance diversity index(H)and simplex index(C)and evenness(J)and Margalef index(d)of phytoplankton
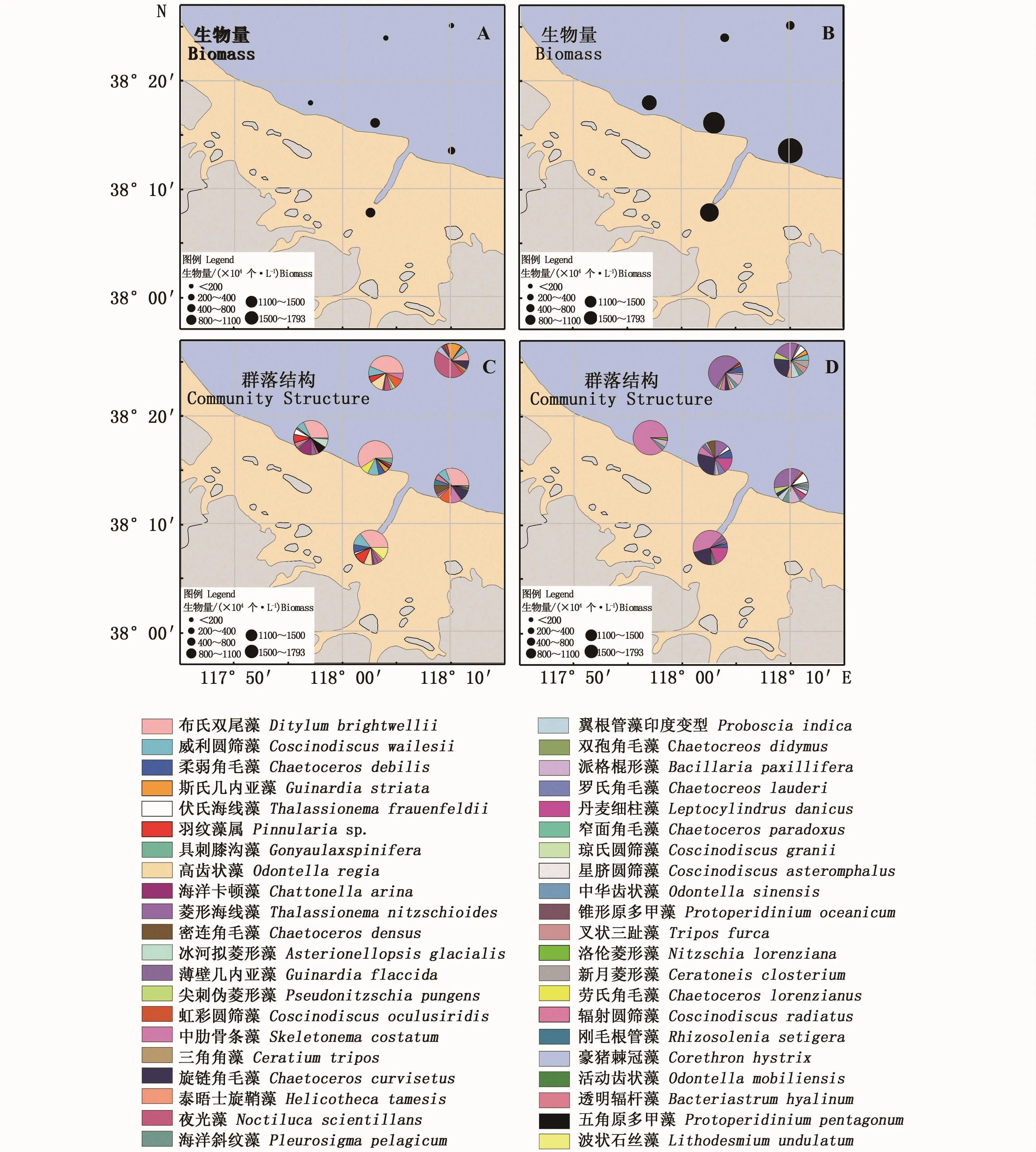
图6 调查海域浮游植物生物量和群落结构分布(A、C:5月,B、D:8月)Fig.6 Spatial distribution of phytoplankton abundance and community structure during the surveys(A,C:May,B,D:August)
2.5 浮游植物群落结构及丰度与环境因子的相关性分析
为了进一步研究增养殖区环境因子对浮游植物群落结构的影响,本研究分别对5月、8月优势种与环境因子进行了RDA分析,结果如图7所示。冗余分析表明,5月RDA的前两个排序轴分别可解释83.3%和14.9%的数据变化。排序轴1主要受氮营养盐浓度和盐度影响,第1轴与NO2-N、NH4-N、DIN、NO3-N和盐度呈正相关关系;排序轴2主要受温度和DIP的影响,与温度和DIP呈现负相关关系。羽纹藻与盐度、氮营养盐浓度呈现负相关关系;夜光藻与温度、DIP呈现负相关关系;威利圆筛藻与布氏双尾藻类似,与DIP和温度有明显的正相关关系。8月RDA的前两个排序轴分别可以解释44.7%和32.7%的数据变化,影响排序轴1的主要环境因子是盐度、温度和DIP,第1轴与盐度呈现正相关关系,与温度和DIP呈负相关关系;影响排序轴2的主要环境因子是盐度和NH4-N,第2轴与盐度呈正相关关系,与NH4-N呈负相关关系。优势种与环境因子的关系大体可以分为3类:1)伏氏海线藻、海洋斜纹藻、菱形海线藻、旋链角毛藻和丹麦细柱藻,它们与盐度呈负相关关系,与磷酸盐浓度和温度呈正相关关系;2)柔弱角毛藻、派格棍形藻和中肋骨条藻,它们与氮营养盐浓度呈现明显的正相关关系,与盐度呈现负相关关系;3)尖刺伪菱形藻与第2类相反,它与盐度呈现正相关关系,与氮营养盐呈现负相关关系。
3 讨论
3.1 环境因子的时空变化特征及其影响因素
环境因子是影响浮游植物群落结构的重要因素。本次调查海域5、8月温度均呈现自近岸至离岸递减的趋势,这与纪大伟[15]在黄河口邻近海域温度调查结果5、8月温度变化趋势相同;温度的平均值偏高,特别是8月份平均温度偏高了4.3℃,这可能与采样时间和年份差异有关。同时本次调查站位布设更靠近陆地,也可能是温度偏高的原因之一。5、8月盐度分布呈现截然相反的趋势,5月河口区盐度高于离岸区域,8月盐度明显呈现自河口至离岸递增的趋势,盐度分布与河口区域丰水期盐度呈羽状分布的研究结果相符[16],说明夏季河流淡水输入对河口区盐度有明显的稀释作用。河口潮间带与浅海交汇处外侧盐度变化不明显,说明套尔河河流输入对调查海域盐度整体影响有限。
本次调查营养盐的时空分布特征与以往调查相似[17-20],营养盐高值区均在河口处,说明陆源排放可能是调查海域氮、磷营养盐的主要来源,这与陈沛沛[21]在套尔河营养盐高值区出现在下游河口处的调查结果类似。与5月份相比,8月氮营养盐结构发生明显变化,硝酸盐的浓度明显降低,亚硝酸盐和氨氮的含量明显升高,这与刘霜等[22]在黄河口附近海域的调查营养盐季节性变化结果基本一致。调查海域5、8月均不存在绝对氮限制,但是部分站位存在绝对磷限制,说明磷限制可能是调查海域重要的限制因子;N/P范围与国内海岸河口N/P范围相似[23],即浮游植物可利用氮含量远大于磷,浮游植物生长可能还存在相对磷限制,这与河口邻近海域普遍存在磷限制的研究结果大致相同[24-26]。放流后(8月)调查海域可溶性无机氮(DIN)和磷酸盐(DIP)与放流前(5月)相比整体呈现降低的趋势,在一定程度上反映出贝类增殖放流活动起到了移除氮磷、净化水质的效果,这与熊永英[27]关于鲢鳙增殖放流活动可以有效缓解水体富营养化、净化水质的研究结果类似。

图7 调查海域浮游优势种与环境因子RDA排序图Fig.7 RDA ordination plot of correlation analysis between phytoplankton dominant species and environmental factors
3.2 环境因子对浮游植物群落结构及丰度的影响
浮游植物群落结构及丰度与水动力过程营养环境有着密切联系[28-29]。5月叶绿素a呈现河口区高于离岸区域的趋势,叶绿素a与环境因子的相关性分析显示,氨氮浓度和盐度是影响春季贝类增养殖区浮游植物生长的主要因素,这也说明春季温度回暖、河口区营养盐浓度高、盐度适宜,有利于浮游植物生长,这与张莹[19]春季的调查结果相似。8月调查海域叶绿素a浓度略高于5月,但变化趋势与5月明显不同,8月河口区叶绿素a低于离岸区域,这与范占伟[30]在东营河口贝类保护区的调查结果相同,叶绿素a与环境因子的相关性分析显示,叶绿素a与温度、盐度、营养盐相关性不显著,这可能是由于夏季径流量增大,水流作用使河口区域水体浑浊度增加,影响了浮游植物光合作用,也可能是由于放流后贝类摄食的作用影响了叶绿素a的分布。
6个典型站位浮游植物鉴定的结果显示,8月浮游植物生物量明显高于5月(图6),但叶绿素a浓度变化并不明显,这可能与夏季浮游植物群落结构组成变化、小粒径的浮游植物占比明显升高有关,例如:8月份优势度较高的物种是旋链角毛藻,粒径通常在3μm左右;5月份优势种主要是布氏双尾藻,粒径通常大于10μm。对5月和8月优势种分析结果显示,优势种季节更替明显,这可能是因为8月份径流量加大,同时水温升高,影响了调查区域的温度、盐度,从而影响浮游植物群落结构组成[31-32]。RDA分析结果显示,环境因子对浮游植物优势种的影响明显。5月,主要优势种布氏双尾藻(占总生物量的43.0%)和威利圆筛藻(占总生物量9.9%)与DIP浓度和温度呈现明显的正相关关系,说明磷酸盐和温度是影响5月份浮游植物组成的最重要的环境因子,它们的升高有利于主要优势种的生长繁殖[33-34]。其次,盐度也是影响优势种结构的因素,羽纹藻与盐度呈负相关关系,说明径流输入明显、盐度较低、水动力复杂的河口区,可能更有利于羽纹藻的生长[35]。8月,磷酸盐依然是影响优势种的关键因子,多种优势浮游植物(如旋链角毛藻、丹麦细柱藻、海洋斜纹藻、伏氏海线藻和菱形海线藻等)与磷酸盐浓度呈正相关关系,磷酸盐浓度的降低抑制了优势种的生长,这与8月调查海域存在明显磷限制的结果吻合。此外,氮营养盐和盐度也是影响优势种的重要因素,柔弱角毛藻、派格棍形藻和中肋骨条藻与氮营养盐浓度呈现明显的正相关关系,与盐度呈现负相关关系,这与此类浮游植物对氮营养盐的需求量高[36]及广温广盐的特性可能有关。
3.3 贝类增养殖对浮游植物群落结构的影响
贝类摄食作用是影响浮游植物群落结构及丰度的另一个重要因素[37-38]。贝类具有摄食偏好[37],倾向于摄食直径大于3μm的颗粒,会使得浮游植物优势种向微型浮游植物转移[39],本次调查海域也显示出类似的趋势,放流后(8月)大粒径浮游植物减少,小粒径浮游植物增多。浮游植物仍以硅藻为主,且占比略有提高,而甲藻的丰度比放流前要小的多,这可能是由于甲藻脂肪酸含量往往高于硅藻,会增加贝类对甲藻的摄食选择[40],同时夏季硅藻易形成细胞长链难以被贝类摄食,而甲藻是单个细胞,更易被贝类摄取[39],因此甲藻在贝类增殖放流后丰度明显减少。
为了进一步探明引起调查海域浮游植物群落变化的主要原因是贝类放流还是季节变化,对环境因子类似的非贝类养殖区的浮游植物群落结构季节变化与本研究进行了对比(表3)。5月,套尔河贝类增养殖区布氏双尾藻、威利圆筛藻为优势种,这与孙慧慧[16]在黄河口区域及张晓举和赵开[14]在莱州湾西部春季的调查结果类似;8月,增养殖区优势种中肋骨条藻、旋链角毛藻、伏氏海线藻、柔弱角毛藻和尖刺伪菱形藻均是夏季常见的浮游植物种群,与非养殖区浮游植物群落结构类似,没有明显差异[40]。浮游植物群落结构变化是多种环境因子综合作用的结果,不同年份间也存在群落结构差别,因此难判别贝类放流对群落结构的具体影响程度[14,16,35,41-42]。同时贝类对浮游植物的影响是双重的,一方面,贝类偏好摄食含有特种形式脂肪酸的浮游植物(如圆筛藻Coscinodiscus、具槽帕拉藻Paraliasulcata、菱形 藻Nitzschia、舟 形 藻Navicula、斜 纹 藻Pleurosigma等[2]),会导致这类浮游植物生物量和丰度减少;另一方面,排泄作用产生大量富含有多种氨基酸的营养物质,促进了这类浮游植物生长。因此,套尔河贝类增养殖区增殖放流活动对浮游植物群落结构的耦合关系具体机制仍有待研究。

表3 本研究与环境类似的非贝类养殖区春夏季浮游植物的优势种对比Tab.3 Comparation of dominant species of phytoplankton in spring and summer between non-shellfish breeding areas with similar environment and this study
4 小结
通过对套尔河邻近海域贝类增养殖区环境因子与浮游植物群落结构的调查发现:
1)调查海域环境因子季节性变化明显,套尔河河流输入对河口邻近海域盐度、营养盐浓度及结构影响明显,丰水期盐度整体呈现由河口向离岸逐渐递增的分布特点,调查区域存在DIP绝对限制和相对限制,且8月DIP限制比5月更明显。
2)调查海域浮游植物群落结构与丰度存在明显的时空差异,河口区域浮游植物丰度明显高于离岸海域,浮游植物群落结构季节性演替明显。RDA分析显示环境因子对浮游植物群落结构及丰度影响明显,5月影响浮游植物群落结构和丰度的主要环境因子是氮营养盐浓度和盐度;影响8月浮游植物群落结构和丰度的主要环境因子是盐度、温度、DIP和NH4-N浓度;不同优势种与环境因子间呈现不同的耦合关系。
3)贝类增养殖有一定的移除氮磷、降低营养盐浓度、改善水体富营养化的作用,但其对环境因子和浮游植物群落结构耦合关系的作用还需要进一步研究。
