基于线粒体COI基因的马鲅科鱼类DNA条形码研究
2020-03-26王业磷邓春兴裴丽梅黄志基
王业磷,章 群,邓春兴,徐 示,裴丽梅,黄志基,罗 纯
(暨南大学生态系,广州 510632)
马鲅科(Polynemidae)隶属于硬骨鱼纲(Osteichthyes)辐鳍亚纲 (Subclass Neopterygii)鲈形目(Perciformes),广泛分布于温带、亚热带和热带近岸海域以及淡水河流的入海口。马鲅科鱼类如六丝多指马鲅(Polydactylussexfilis),肉质鲜美、营养价值高;还有一些种类如长指马鲅(Polynemusparadiseus),形态优美、有较强的观赏性,是珍稀名贵的经济鱼类[1-2]。全世界已知马鲅科鱼类共有8属43种,其中中国有3属7种[3-5],分别为四指马鲅属(Eleutheronema)的四指马鲅(E.tetradactylus)和多鳞四指马鲅(E.rhadinum),多指马鲅属(Polydactylus)的五指多指马鲅(P.plebejus)、黑斑多指马鲅(P.sextarius)、六丝多指马鲅和小口多指马鲅(P.microstomus),以及丝指马鲅属(Filimanus)的西氏丝指马鲅(F.sealei)。在过度捕捞和海洋环境日益恶化的大背景下,马鲅科鱼类的渔业资源在不断减少,特别是市场需求大、价格昂贵的四指马鲅等具高经济价值的马鲅科鱼类,对于该科鱼类的多样性保护十分不利,亟需开展马鲅科鱼类种质资源保护和开发利用研究[2]。
准确的物种鉴定是种质资源保护和开发利用的前提。目前,MOTOMURA[1]对马鲅科鱼类基于形态学的再描述是该科鱼类分类的主要依据;叶振江等[6]对四指马鲅、五指多指马鲅和六丝多指马鲅以及常有民等[7]对多鳞四指马鲅的耳石形态特征的观察,也为该科鱼类的分类提供了部分依据。但由于马鲅科一些种类外部形态相似度高、分类特征不显著,极易混淆;另外颜色褪变或保存不完整以及不同发育阶段的形态变化,也给鉴定带来较大困难。四指马鲅属鱼类虽是名贵的海产鱼类,但早期的研究资料认为中国沿海分布只有四指马鲅一种,中国没有多鳞四指马鲅的记录[3-4]。在多鳞四指马鲅作为一个独立有效种确立后,有研究[8-9]认为原先在中国分布的四指马鲅全部为多鳞四指马鲅,而四指马鲅在中国并无分布。四指马鲅与多鳞四指马鲅形态区分主要是胸鳍颜色的不同,但由于颜色差异并不稳定以及存在着连续变化的可能,常常被误认为同一个种。DNA条形码技术[10-11]只需极少量的样品,通过一段标准的DNA序列的比较分析就可以对物种进行准确鉴定,从而避免了表型性状鉴定物种存在的不足。已有研究表明,线粒体COI基因5′端一段长度约652 bp的序列作为物种鉴定 的 DNA 条 形 码 在 鱼 类[12]、鸟 类[13]和 昆虫[14-15]等动物上都表现出了高可靠性和便利性,且在解决同种异名、异种同名和发现隐存种等方面有着突出优势,已成为了动物界首选DNA条形码标记[16]。
目前,中国马鲅科鱼类DNA条形码的研究仅见有严安琪等[17]对浙江海域四指马鲅和黑斑多指马鲅以及邓春兴[18]对中国近海四指马鲅属等少数物种局部地区的研究。本研究测定了在中国黄海、东海和南海7省10个地点采集的3属5种33条标本的线粒体COI序列,结合GenBank中下载的5属16种的COI序列,进行中国马鲅科鱼类DNA条形码研究,旨在明确该科鱼类的分类地位,为中国马鲅科鱼类种质资源的保护和开发利用提供科学依据。
1 材料与方法
1.1 实验材料与分子实验
依据 MOTOMURA[1]工作和《南海鱼类检索》[19]等相关资料对本实验室多年保存的马鲅科鱼类进行鉴定分类,提取保存于95%的乙醇中的中国马鲅科鱼类肌肉组织用于测定COI序列。本研究测序标本及GenBank下载序列来源详见表1。
1.2 DNA提取、PCR扩增与测序
采用酚-氯仿抽提法[20]提取总 DNA后,于-20℃中保存。使用通用引物[12]FishF1:5′TCAACCAACCACAAAGACATTGGCAC3′,FishR1:5′TAGACTTCTGGGTGGCCAAAGAATCA3′对线粒体COI基因进行PCR扩增。反应体系为30μL:15μL mix,1μL FishF1,1μL FishR1,12μL ddH2O,1μL DNA。反应条件为:95℃预变性5 min;94℃变性40 s,54℃退火40 s,72℃延伸50 s,共35个循环;最后再72℃延伸10 min。PCR产物经1%琼脂糖凝胶电泳检测合格后送华大基因公司进行测序。
1.3 数据处理
经BioEdit v 7.0.9对原始序列峰图人工校对,与从GenBank下载的COI序列联并后进行比对。用MEGA X计算碱基组成、变异位点、简约位点以及转换/颠换值;基于 K2P(Kimura2-parameter)模型,经1 000次重复抽样检测置信度构建邻接(neighbor-joining)树;分别计算种内种间和分支内分支间的遗传距离。在 https://bioinfo.mnhn.fr/abi/public/abgd/abgdweb.html网站上利用 ABGD(automatic barcode gap discovery)法进行假设种分析[21]。
2 结果与分析
2.1 马鲅科鱼类COI基因序列分析
在马鲅科鱼类6属20种74个体平均长度为648 bp(620 bp~652 bp)的COI基因序列中,没有碱基插入或缺失,保守位点394个,变异位点258个,简约信息位点251个。T、C、A、G平均含量分别为:28.9%、29.3%、22.3%、19.5%,A+T含量(51.2%)高于G+C含量(48.8%),有一定的偏倚性,与其他硬骨鱼类COI序列碱基特征基本一致[12]。转换与颠换比3.37,说明序列突变没有达到饱和,适用于系统发育分析[22]。
2.2 种内与种间遗传距离
基于K2P模型计算出马鲅科20种鱼类的种内遗传距离为0~11.9%,平均遗传距离为0.9%;种间遗传距离为0.1%~27.4%,平均遗传距离为19.6%;种间平均遗传距离是种内平均遗传距离的22倍,符合HEBERT[10-11]提出的物种鉴别“十倍法则”,即种间遗传距离远大于种内遗传距离的10倍以上。在DNA条形码研究中,除腔肠动物门(Cnidaria)外,2%是绝大部分物种划分的参考值[10-11],即种内遗传距离一般小于2%,种间一般大于2%。六丝多指马鲅种内遗传距离为11.9%,远大于2%;多鳞四指马鲅种内遗传距离为2.3%;其余种类均远小于2%。黑斑多指马鲅与马达加斯加多指马鲅(Polydactylusmalagasyensis)的种间遗传距离为1.0%,马伦氏多指马鲅(Polydactylusmullani)与七丝指马鲅(Filimanusheptadactylus)种间距离仅为0.1%,都小于2%;其他各种类间的种间遗传距离均远大于2%。
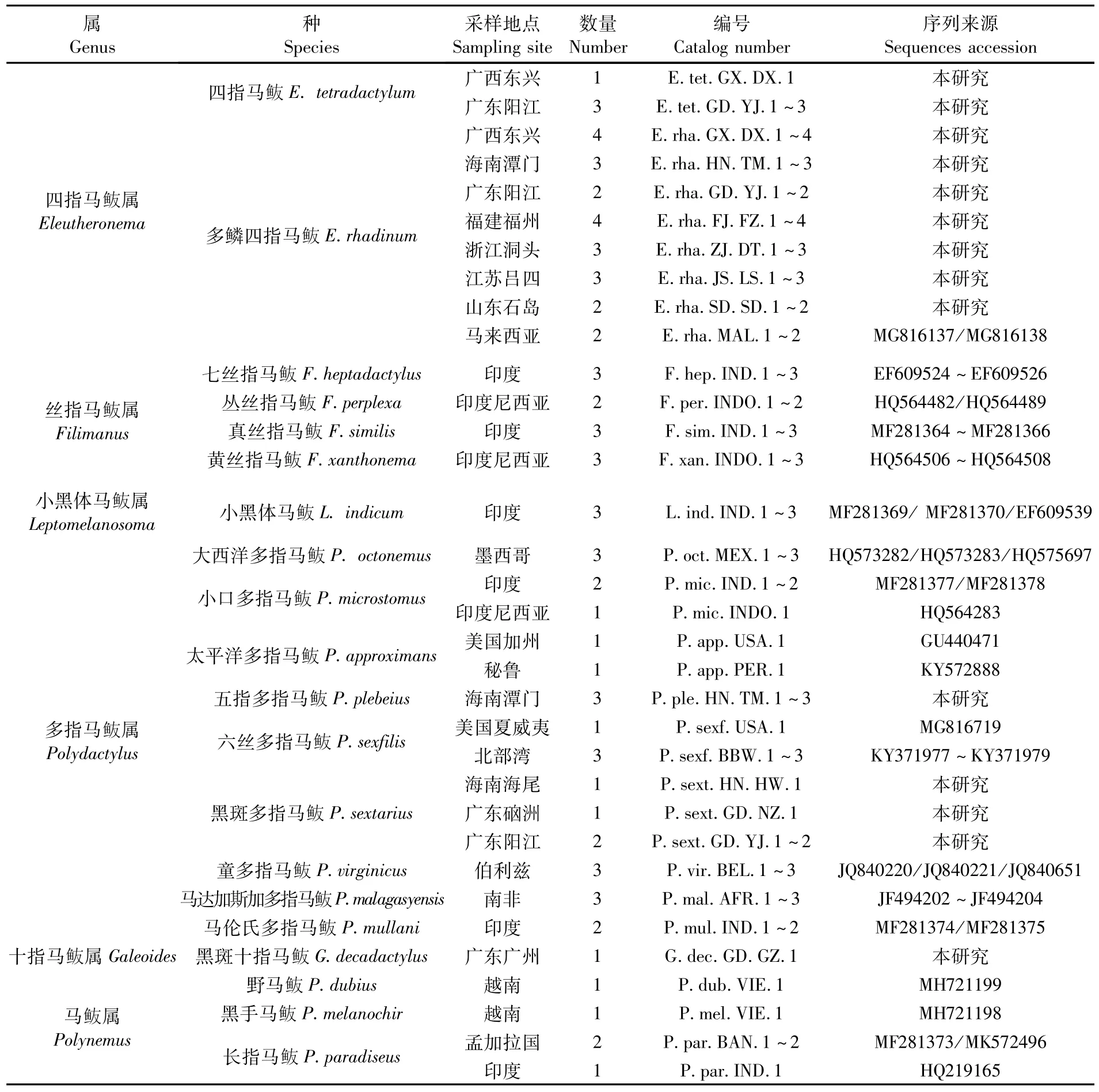
表1 本研究中用于COI分析的马鲅科鱼类样本的信息Tab.1 Specimen information of Polynemidae in the present study
2.3 马鲅科鱼类分子系统发育树
在邻接树中(图1),74条序列形成了19个分支;分支间的平均遗传距离为20.4%(2.8%~27.6%)是分支内平均遗传距离0.1%(0.0%~0.2%)的204倍。20种鱼类有14种形成了单系分支,支持其物种有效。多鳞四指马鲅形成了2个自展数据支持率为99%的分支,部分福州个体、全部东兴个体以及马来西亚个体形成1支(Clade2);广泛分布于中国沿海的其余个体形成1支(Clade1);分支间遗传距离为4.8%。黑斑多指马鲅、北部湾的六丝多指马鲅和马达加斯加多指马鲅混杂于Clade16中,自展数据支持率为99%,分支内遗传距离为0.7%。马伦氏多指马鲅与七丝指马鲅混杂于Clade14中,自展数据支持率为99%,分支内遗传距离仅为0.1%。四指马鲅属、马鲅属分别形成单系群。在ABGD分析中(图3),当种内先验遗传距离P为0.46%~2.15%时,马鲅科被分为16个假设种。夏威夷的六丝多指马鲅(Clade4)与五指多指马鲅(Clade5)被归为一个假设种;马鲅属的长指马鲅(Polynemusparadiseus)、黑手马鲅(P.melanochir)和野马鲅(P.dubius)(Clade6、Clade7、Clade8)也被归为一个假设种,这与遗传距离和邻接树的结果不一致;其他种的划分与邻接树形成的分支相一致。
3 讨论
马鲅科20种鱼类的种间平均遗传距离(19.6%)是种内平均遗传距离(0.9%)的22倍;在邻接树上形成了14个单系分支,分支间平均遗传距离(20.4%)是分支内平均遗传距离(0.1%)的204倍,表明存在明显的DNA条形码间隙,符合鉴别物种的“十倍法则”,说明COI基因适用于作为马鲅科鱼类鉴定的DNA条形码。
多鳞四指马鲅最早由JORDAN等[23]根据中国台湾样品提出,但在早期,多鳞四指马鲅被认为是四指马鲅的年幼个体[24]。MOTOMURA等[25]于2002年根据侧线鳞数(82~95vs71~80,前者为多鳞四指马鲅,下同)和胸鳍颜色(黑色vs黄色)等形态特征将其与四指马鲅分开,并确定为一个独立的有效种被采用至今。在本研究中,多鳞四指马鲅与四指马鲅种间遗传距离为16.0%,远大于2%种内遗传阈值,且是种内平均遗传距离1.2%的13倍,从分子水平上支持了该分类的有效性。但多鳞四指马鲅种内遗传距离(2.3%)略大于2%的物种遗传阈值,且在NJ树上形成了2个明显的分支,分支间遗传距离(4.8%)是分支内平均遗传距离(0.1%)的48倍,符合“十倍法则”。杨喜书[26]发现南海印度鲬(Platycephalusindicus)种内遗传距离为4.8%,形成了2个支持率为99%的分支,且分支间的遗传距离(10.4%)是分支内平均遗传距离(0.4%)的26倍,认为印度鲬可能存在隐存种。陈璐等[27]发现三线鯻(Teraponputa)种内遗传距离(2.7%)大于2%,形成了2个分支,分支间遗传距离为5.0%,但与分支内的平均遗传距离(0.85%)不满足“十倍法则”,认为其可能为两个亚种或不同种。参照上述研究,推测中国沿海分布的多鳞四指马鲅可能已分化成2个亚种乃至2个物种,其中东兴个体和部分福州个体为一种或亚种,且在马来西亚见有分布;另一种或亚种在中国黄海、东海以及南海都有分布。多鳞四指马鲅是海水鱼中的名贵品种,同时作为潜在的养殖新品种,具有广阔的开发前景[28];在今后种质资源开发、利用和保护中,应考虑将其归为2个管理保护单位。
六丝多指马鲅形成了2个距离较远的分支:1)夏威夷个体独立成支;2)北部湾个体与黑斑多指马鲅和马达加斯加多指马鲅混为一支。夏威夷的六丝多指马鲅个体在BOLD网站上有照片记录,经仔细比对后确定鉴定无误。黑斑多指马鲅在中国沿海较常见[29-30],六丝多指马鲅较稀少,现有报道分布于中国南海南部及中国台湾[5,19],在北部湾尚未发现有其记录;二者都具有6条游离胸丝,但前者侧线起点附近具有一黑色大斑,后者无黑斑[1]。北部湾个体可能是因游离胸丝数相同而被误鉴定成六丝多指马鲅,实际应为黑斑多指马鲅。游离的胸丝数是马鲅科鱼类分类的重要特征之一,但由于有些马鲅科鱼类具有相同的胸丝数,且有时胸丝数还可能发生变异,如本实验室还获得具5条胸丝的多鳞四指马鲅;因此,不能以此作为其鉴定的唯一特征,需要结合多个不同特征来确定。

图1 基于线粒体COI基因序列的20种马鲅科鱼类邻接树Fig.1 Neighbor-joining tree based on mtDNA COI sequences of 20 Polynemid species

图2 19个分支的分支内(a)与分支间(b)的遗传距离Fig.2 Histograms of genentic distances within(a)and among 19 clades(b)

图3 马鲅科鱼类的ABGD分析图Fig.3 Automatic barcode gap discovery analysis of Polynemidae
黑斑多指马鲅与马达加斯加多指马鲅的种间遗传距离为1.0%,在邻接树上混为一支。黑斑多指马鲅分布广泛,从印度西南部沿岸到巴布亚新几内亚再到日本宫崎;马达加斯加多指马鲅目前只见于马达加斯加、莫桑比克以及南非沿岸[1];两者外部形态极为相似,都有6条游离胸丝和侧线起点附近有一黑色大斑块,曾被认为同一个种[31],但前者的鳃耙数(25~30)少于后者(29~35),鳔退化,且第二背鳍基部长相对明显短于后者,被确定为新种[32]。许多研究表明,某些种类在形态上虽有一定的差异,但在分子上可以表现出相近的关系,如唐楚林等[33]发现红鳍笛鲷(Lutjanuserythopterus)与马拉巴笛鲷(L.malabaricus)的种间遗传距离仅为0.3%,但可以根据侧线下鳞的排列方式进行区分;宫亚运等[34]发现中颌棱鳀(Thryssamystax)与黄吻棱鳀(T.vitrirostris)的种间遗传距离只有0.6%,但可以通过背鳍数和鳃耙数进行区分。黑斑多指马鲅与马达加斯加多指马鲅的情况与之类似,可能为同一种的2个亚种;同时由于标准COI条形码序列长度较短且保守,对近期分化形成的物种往往难以区分[35],也有可能为近期进化形成的2个不同物种。
马伦氏多指马鲅与七丝指马鲅也混为一支,种间遗传距离为0.1%;它们都有7条游离胸丝,也曾被混淆[24],但前者侧线起点附近有一黑色大斑块,后者没有,可能仍属于同一物种或为近期分化形成的不同种。马伦氏多指马鲅、七丝指马鲅和马达加斯加多指马鲅的序列源自于GenBank,笔者无法对对应的鱼类进行形态鉴定确认;由于形态过于相似,也不排除形态鉴定出错的可能。
黑斑十指马鲅(Galeoidesdecadactylus)主要分布于非洲西海岸[1],该鱼在中国尚未有记录,本实验室样品购自广州市场,但未知其明确产地。五指多指马鲅较少见,本实验室也只获得几条,同时在中国大陆有记录的六丝多指马鲅以及在中国台湾有报道的小口多指马鲅和西氏丝指马鲅都未能采集到,特别是多鳞四指马鲅的分类还存在问题等,在今后的研究中,还需进一步扩大采样的范围,补充样品的种类和数量,在详细的形态测量基础上结合多种分子标记展开研究,以期进一步明确中国马鲅科鱼类的分类地位。
