肺形侧耳种质资源拮抗反应及RAPD分子标记遗传多样性分析
2020-03-24陈躬国陈剑吴汉琼刘新锐方洪枫骆志坚
陈躬国 陈剑 吴汉琼 刘新锐 方洪枫 骆志坚

摘要:【目的】分析肺形側耳种质资源拮抗反应和遗传多样性,为肺形侧耳种质资源鉴定、合理利用和优良菌株遗传选育等提供参考依据。【方法】分别通过拮抗反应和RAPD分子标记对41个肺形侧耳菌株进行鉴定及遗传多样性分析,并对这2种方法的聚类分析结果进行比对及相关性分析。【结果】拮抗反应试验结果显示,仅有少数菌株与其他菌株间出现明显拮抗反应,多数菌株间无拮抗反应或拮抗较弱,可能存在同种异名的现象。基于拮抗反应进行聚类分析,结果显示在遗传相似性系数0.65处,可将41个肺形侧耳菌株分为五大类群,第I和III类群分别包括1个菌株,第II类群包括31个菌株,第Ⅳ类群包括4个菌株,第Ⅴ类群包括4个菌株,拮抗反应明显的肺形侧耳菌株聚在不同的类群,无拮抗反应或拮抗反应弱的菌株则聚在同一类群里。RAPD分析结果显示,15条RAPD引物共扩增出117个位点,多态性位点有96个,多态性比例为82.1%;41个肺形侧耳菌株遗传相似系数为0.51~1.00,在遗传相似系数0.84处可将41个肺形侧耳菌株分为五大类群,其中,第Ⅰ类群包括3个菌株,第II类群包括4个菌株,第III类群包括6个菌株,第Ⅳ类群仅包括1个菌株,剩余的菌株属于第Ⅴ类群,结合拮抗反应试验结果发现有明显拮抗反应的菌株聚在不同类群。拮抗反应和RAPD分析的聚类结果具有显著相关性(r=0.674,t=5.95),表明拮抗反应和RAPD分子和标记在反映肺形侧耳种质资源遗传多样性上具有较强的一致性,均能很好体现肺形侧耳种质的遗传关系。【结论】肺形侧耳菌株存在同种异名和同名异种现象。在实际生产中可选择亲缘关系较远的材料作为亲本菌株,以增加后代的遗传变异,从而选育出综合性状优良的肺形侧耳菌株。
关键词: 肺形侧耳;拮抗反应;RAPD;遗传多样性;聚类分析
中图分类号: S646.14 文献标志码: A 文章编号:2095-1191(2020)12-2892-09
Abstract:【Objective】The object of this study was to analyze the antagonistic reaction and genetic diversity of Pleurotus pulmonarius germplasms, and provide reference for the identification,reasonable utilization of germplasms and genetically breeding of P. pulmonarius strains. 【Method】Forty-one strains of P. pulmonarius were analyzed by antagonistic reaction and RAPD molecular markers on their identification and genetic diversity. And the cluster analysis results of the two methods were compared and correlated. 【Result】The results of the antagonistic reaction test showed that there were obvious antagonistic reactions among some strains of P. pulmonarius, but there were no antagonistic reactions or weak antagonistic reactions among most strains. Also, phenomenons of synonym possibly existed in the P. pulmonarius germplasms. Cluster analysis based on antagonistic reaction showed that 41 strains of P. pulmonarius could be clustered into five groups at the genetic similarity coefficient of 0.65. Group I and III included 1 strain respectively, group II included 31 strains, group IV included 4 strains, and group V included 4 strains. The strains with obvious antagonistic reaction were clustered in different groups, the strains with no antagonistic reaction or weak response or antagonism were clustered in the same group. A total of 117 sites were amplified by 15 RAPD primers, 96 of which were polymorphic, with a polymorphism rate of 82.1%. The genetic similarity coefficient of the 41 strains of P. pulmonarius ranged from 0.51 to 1.00, and cluster analysis showed that the tested strains could be divided into five groups at the level of genetic similarity 0.84. Group I included 3 strains, group II included 4 strains, group III included 6 strains, group Ⅳ included only 1 strain, and the remaining strains belonged to group Ⅴ. Strains with antagonistic reaction clustered in different groups in RAPD cluster analysis. The clustering results of antagonistic response and RAPD analysis were significantly correlated (r=0.674, t=5.95), indicating that there was closely accordance between antagonistic reaction and RAPD molecular markers analysis, suggesting both analysis could well reflect the genetic relationship of P. pulmonarius germplasms. 【Conclusion】Phenomenon of different species with the same names and different names with the same species exist in P. pulmonarius. In actual production, germplasms with far genetic distance can be selected as parents to increase the genetic variation of the offspring, so as to breed the strains of P. pulmonarius with good comprehensive characters.
Key words: Pleurotus pulmonarius; antagonistic reaction; RAPD; genetic diversity; cluster analysis
Foundation item: National Edible Fungi Industry Technical System Funding Project(CARS-20); Guiding Project of Fujian Science and Technology Department(2018N0035); Regional Development Project of Fujian Science and Technology Department(2016N3018); Science and Technology Project of Fuzhou (2016-N-106)
0 引言
【研究意义】肺形侧耳[Pleurotus pulmonarius (Fr.) Quél.]商品名为秀珍菇,隶属于担子菌纲(Basidiomycetes)伞菌目(Agaricales)侧耳科(Pleurota-ceae)侧耳属(Pleurotus),富含多种蛋白质、必需氨基酸、多糖、重要矿物质及维生素,营养价值高,还具有抗氧化活性、降低血糖及辅助肿瘤化疗等药理活性(Wahab et al.,2014;Xu et al.,2014;Abidin et al.,2016)。目前肺形侧耳同种异名、同名异种现象较普遍(朱坚等,2007;忻雅等,2008),不仅影响实际生产中的菌种管理,还给种性鉴定、良种选育等科研及产业体系建立带来巨大阻碍。因此,对肺形侧耳种质资源进行鉴定和遗传多样性分析,有助于加强其菌种管理和新品种权保护,指导优良菌株选育中的亲本选配,对促进整个产业的持续健康发展具有重要意义。【前人研究进展】担子菌体细胞的非亲和性(Samatic incompatibility,SI)是担子菌在双核菌丝阶段发生的认识自我和排斥异己的过程,是保持群体遗传多样性的重要反映(Worrall,1997)。体细胞的非亲和性往往导致菌丝体间出现明显的分界线即拮抗反应,包括色素形成、致密的菌丝隆起、菌丝稀疏区域等,是体细胞非亲和性的具体体现。拮抗反应是确定体细胞不亲和性最简便的方法,广泛应用于食用菌菌株的初步鉴定和分类(贾定洪等,2013;王锦锋等,2017)。李黎(2011)通过木耳栽培种质资源的拮抗反应分析其遗传多样性,结果表明木耳菌株间的拮抗程度与亲緣关系呈正比,拮抗反应可有效地反映木耳种质的遗传多样性。谭伟等(2015)利用拮抗反应分析了通过种间杂交获得的黄白侧耳新菌株与亲本之间的遗传差异,证实了杂交菌株的真实性。RAPD分子标记已广泛应用于种内和种间的亲缘关系分析、个体鉴定、分类及差异表达基因筛选(Fonseca et al.,2008;温志强等,2011;杨军等,2016;张安世等,2018)。朱坚等(2007)通过RAPD分子标记对肺形侧耳菌株进行综合分析,结果发现菌株间遗传差异小,且存在同名异种或同种异名的现象。叶长文等(2013)利用RAPD分子标记分析了香菇品种135与自然变异抗性新菌株庆元9015的基因片段差异,对差异基因片段进行克隆和测序,结果发现其与水解酶或信息素受体的部分氨基酸序列有较高的同源性,可能与香菇对温度的敏感性有关。李红等(2018)应用RAPD分子标记对35个金针菇菌株进行遗传多样性分析,结果显示,该标记能将白色金针菇和黄色金针菇区分开来,与菌株农艺性状分类结果一致,说明RAPD分子标记用于菌类遗传多样性分析是可行的。【本研究切入点】虽然李维焕等(2010)、冯伟林等(2014)对肺形侧耳进行了遗传多样性分析,但供试材料较少,不够全面。目前,基于拮抗反应试验结果进行肺形侧耳聚类分析,以及利用RAPD分子标记进行其遗传多样性分析的研究鲜见报道。【拟解决的关键问题】分别通过拮抗反应和RAPD分子标记对41个肺形侧耳菌株进行鉴定及遗传多样性分析,并对这2种方法的聚类分析结果进行比对及相关性分析,为肺形侧耳种质资源鉴定、合理利用和优良菌株遗传育种等提供理论依据。
1 材料和方法
1. 1 试验材料
供试材料为41个肺形侧耳菌株,其来源如表1所示。PDA固体培养基、PD液体培养基、E.Z.N.A.? SP Plant DNA Kit购于Omega Bio-Tek公司。PCR随机引物委托上海生工生物工程有限公司合成。
1. 2 拮抗反应试验
将供试菌株活化后,在同一PDA固体培养基上按“品”字形接种的3个不同肺形侧耳菌株,置于25 ℃恒温箱中暗光培养10~13 d,以自身拮抗为对照,根据NY/T 1845─2010《食用菌菌种区别性鉴定(拮抗反应)》判定标准,观察不同菌株菌落间是否有菌丝隆起、有无色素沉淀、有无沟带等方面的特征及这些特征的强弱来判断肺形侧耳菌株间拮抗反应的有无和强弱。
1. 3 基因组DNA提取
将供试菌株进行活化后接种于PD液体培养基中,25 ℃下150 r/min恒温培养8 d。收集新鲜菌丝体经液氮研磨后,使用E.Z.N.A. ?Plant DNA Kit提取其基因组DNA,并用1%琼脂糖凝胶电泳进行检测其提取效果,紫外分光光度计测定纯度,-20 ℃保存备用。
1. 4 RAPD分析
试验所用的PCR随机引物为16个10碱基的寡核苷酸引物(表2)。反应体系25.0 ?L:10×PCR Buffer 2.5 ?L,25 mmol/L MgCl2 2.0 ?L,2.5 mmol/L dNTPs 1.4 ?L,5 U/?L Taq DNA聚合酶0.3 ?L,10 ?mol/L引物1.0 ?L,基因组DNA 模板0.4 ?L(约50 ng DNA),ddH2O补足至25.0 ?L。扩增程序为:94 ℃预变性5 min;94 ℃ l min,36 ℃ l min,72 ℃ 90 s,共进行35个循环,72 ℃延伸10 min。PCR产物用1.0%琼脂糖凝胶电泳检测,在GelDoc-It凝胶成像系统拍照观察。
1. 5 统计分析
观察电泳图谱并记录所有位点的二元数据,即在同一迁移位置上有条带赋值为“1”,无条带赋值为“0”,形成0/1矩阵。使用NTSYS-pc 2.10e计算肺形侧耳菌株间遗传相似系数,采用非加权组平均法(UPGMA)进行聚类分析并构建肺形侧耳菌株的系统发育进化树。最后,用Mantel检验进行协表征矩阵和遗传相似系数矩阵间的相关性分析,以检测聚类结果的可靠性。
2 结果与分析
2. 1 拮抗反应检测结果
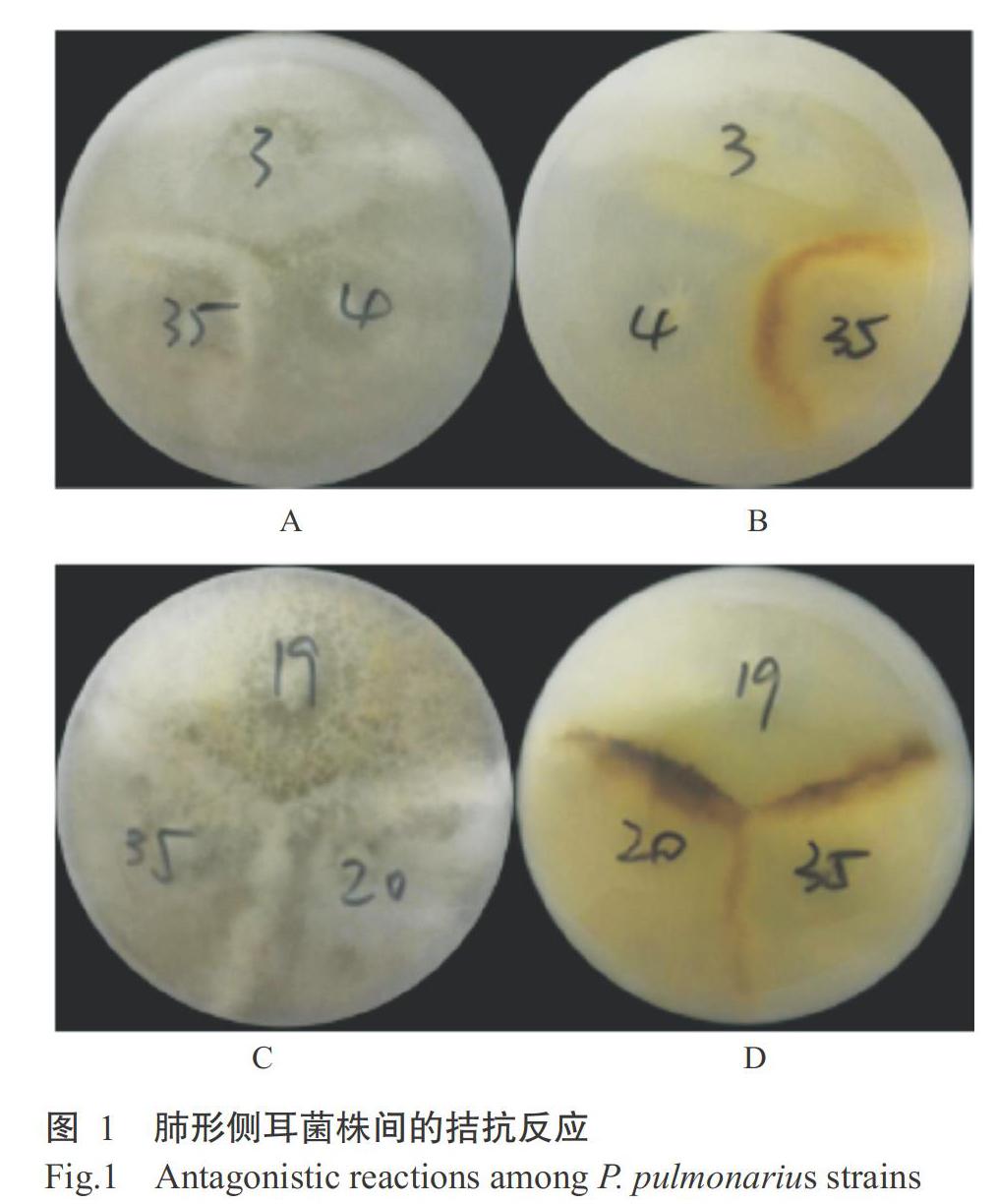
拮抗反应试验采用三点法对不同菌株两两组合进行对峙培养,来自不同遗传背景的肺形侧耳菌株菌丝的交汇区会形成分界线,除了个别菌株间的拮抗反应是隔离型外,大部分肺形侧耳菌株的拮抗现象几乎都是发生菌丝隆起的同时伴随着色素沉淀,从平板背面看色素颜色从淡黄到深褐(图1)。从菌丝隆起和色素沉淀的情况判断菌株间拮抗反应的有无与强弱,结果如表3所示。P1(秀罗源)与P2(秀珍菇)、P6(秀76)、P7(秀台1号)、P8(秀58)、P9(HC810)等17个菌株产生不明显的拮抗线,但与P22(凤尾菇)、P31(秀珍菇845)、P34(秀A)和P35(夏秀888)产生较明显的拮抗线;P12(中农秀珍菇)、P19(秀珍菇77)和P31(秀珍菇845) 3个菌株间相互无拮抗,但与其他菌株有拮抗反应;P10(夏丰一号)、P26(夏秀66)、P30(秀珍菇3号)和P35(夏秀888)4个菌株间相互无拮抗线,与其他菌株均产生较明显或不明显拮抗线;P22(凤尾菇)与其他菌株间均有不明显或较明显拮抗线,为独立菌株;其他大部分菌株间未产生拮抗线或拮抗线不明显。上述结果表明,仅有少数菌株与其他菌株间出现明显拮抗反应,多数菌株间无拮抗反应或拮抗较弱,可能存在同种异名的现象。
以肺形侧耳菌株间的拮抗反应强弱作为变量进行赋值。菌株间无拮抗反应(-),赋值为“0”,拮抗反应较弱不明显(+),赋值为“1”,拮抗反应较明显(++),赋值为“2”,拮抗反应明显(+++),赋值为“3”,建立数据矩阵,采用UPGMA进行聚类分析。将聚类结果转换为协表征矩阵,协表征矩阵和遗传相似系数矩阵的相关系数(r)为0.99,表明聚类结果能很好地体现肺形侧耳种质间的遗传关系。由图2可知,在遗传相似性系数0.65处,可将41个肺形侧耳菌株分为五大类群,除P1(秀罗源)和P40(台秀122)分别单独聚为第I类群和第III类群外,其余菌株聚为3个类群,其中,第II类群包括P8(秀58)、P17(秀63)、P24(凤尾5号)、P29(日本秀珍菇)和P33(秀12)等31个肺形侧耳菌株,菌株间的遗传相似系数均为0.68;第Ⅳ类群包括P10(夏丰一号)、P26(夏秀66)、P30(秀珍菇3号)和P35(夏秀888)等4个肺形侧耳菌株,菌株间的遗传相似系数均为0.74;第Ⅴ类群包括P12(中农秀珍菇)、P19(秀珍菇77)、P22(凤尾菇)和P31(秀珍菇845)等4个肺形侧耳菌株,菌株间的遗传相似系数为0.76。整体来看,拮抗反应明显的肺形侧耳菌株聚在不同的类群,无拮抗反应或拮抗反应弱的菌株则聚在同一类群里。
2. 2 肺形侧耳菌株RAPD分析结果
用16个寡核苷酸引物对41个供试肺形侧耳菌株的基因组DNA进行扩增,除引物S56扩增效果不理想外,其余15个引物扩增效果较好,共扩增出117个RAPD位点,各引物扩增的位点数为5~13个,平均每个引物扩增7.8个位点,其中多态性位点有96个,多态性比例为82.1%。引物S60和S85对肺形侧耳菌株的电泳结果如图3和图4所示。
综合这些引物扩增出的多态性位点,采用UPGMA对肺形侧耳菌株进行聚类分析。将聚类结果转换为协表征矩阵,协表征矩阵和遗传相似系数矩阵的相关系数(r)为0.96,表明RAPD聚类结果能很好地体现肺形侧耳菌株间的遗传关系。41个肺形侧耳菌株间遗传相似系数为0.51~1.00,其中菌株P12(中农秀珍菇)与P33(秀12)的遗传相似系数最小,为0.51,说明二者亲缘关系最远;菌株P4(秀迪1号)与P5(秀57)、P13(秀57-1)与P14(秀98)的遗传相似系数最大,均为1.00,表明菌株间親缘关系很近,极有可能是同种异名。在遗传相似系数0.60处,可将41个肺形侧耳菌株分为两大类群,菌株P12(中农秀珍菇)、P19(秀珍菇77)和P31(秀珍菇845)聚为一个类群,说明这3个菌株与其他菌株遗传差异明显,亲缘关系较远。在遗传相似系数0.84处,可将41个肺形侧耳菌株分为五大类群,即第Ⅰ类群包括P12(中农秀珍菇)、P19(秀珍菇77)和P31(秀珍菇845)等3个菌株;第II类群包括P10(夏丰一号)、P26(夏秀66)、P30(秀珍菇3号)和P35(夏秀888)等4个菌株;第III类群包括P33(秀12)、P34(秀A)、P36(秀珍菇163)、P39(秀珍16)、P40(台秀122)和P41(农秀1号)等6个菌株;第Ⅳ类群仅包括P3(秀917);剩余肺形侧耳菌株属于第Ⅴ类群(图5)。结合拮抗反应试验结果可知,有拮抗反应的菌株聚在不同类群。
2. 3 拮抗反应与RAPD分析的相关性分析结果
为了检验肺形侧耳种质资源的拮抗反应和RAPD分析结果的相关程度,利用Mantel检验对基于拮抗反应聚类分析的遗传相似系数矩阵和基于RAPD分析的遗传相似系数矩阵进行相关性分析,结果表明,拮抗反应试验和RAPD分析的结果具有显著相关性(r=0.674,t=5.95),说明拮抗反应和RAPD分子标记在反映肺形侧耳种质资源遗传多样性上有较强的一致性。
3 讨论
体细胞的非亲和性现象在子囊菌、担子菌和半知菌等真菌中均有报道(Williams and Wilson,1966;Worrall,1997;Cortesi and Milgroom,1998),而拮抗反应是担子菌的体细胞不亲和性的一种表现形式(黄亦存,1996)。拮抗反应试验是生理水平上鉴定不同菌株间遗传差异的传统方法,亲缘关系较近的菌株间无拮抗或拮抗不明显,若是亲缘关系较远,则菌株间有明显的拮抗反应。本研究拮抗反应试验发现,来自不同遗传背景的肺形侧耳菌株间出现隆起型拮抗线且在培养基平板背面有褐色色素沉淀,仅个别菌株间的拮抗反应是隔离型。试验中还发现部分菌株间菌落形态不一致,但并无隆起型或隔离型拮抗线,平板背面也无色素沉淀,推测其为体细胞亲和、亲缘关系相近的不同菌株。拮抗反应作为菌株鉴定的传统方法,虽然具有一定的局限性,但在识别同种异名、鉴定菌株之间的亲缘关系和遗传多样性等方面仍为一种简单有效的方法。通过拮抗反应结合分子标记的方法对菌株的亲缘关系和遗传多样性进行综合分析可得到更可靠的结果。本研究基于RAPD引物扩增出结果,采用UPGMA对肺形侧耳菌株进行聚类分析,结果表明41个肺形侧耳菌株间遗传相似系数为0.51~1.00,大部分菌株间遗传相似系数较高,说明菌株的亲缘关系较近,少数菌株间遗传相似系数为1.00,说明这些肺形侧耳菌株极有可能为同种异名。究其原因可能和肺形侧耳栽培品种主要引自我国台湾地区有关,造成基因资源较狭窄。间接说明肺形侧耳和其他食用菌一样存在引种频繁、穿插引种和部分菌株间存在同种异名等现象。
拮抗反应与分子标记分析结果相吻合在丛枝菌根(AM)真菌(Giovannetti et al.,2003)、木耳(李黎,2011)和灵芝等(潘丽晶等,2013)等中均有报道。但戚元成等(2010)将糙皮侧耳的19个菌株间的拮抗线反应类型作为变量进行聚类分析,再与基于RAPD分子标记分析的聚类结果进行比较,结果发现这2种方法得到的聚类结果存在明显差异,无相关性(r=0.115)。本研究利用Mantel检验对拮抗反应聚类分析的遗传相似系数矩阵和RAPD标记聚类分析的遗传相似系数矩阵进行相关性分析,结果发现,二者具有显著相关性(r=0.674,t=5.95);两种方法的聚类结果均显示,P12(中农秀珍菇)、P19(秀珍菇77)和P31(秀珍菇845)聚为一类,菌株P10(夏丰一号)、P26(夏秀66)、P30(秀珍菇3号)和P35(夏秀888)聚为一类,表明拮抗反应和RAPD标记在反映肺形侧耳种质资源遗传多样性上具有较强的一致性。
4 结论
肺形侧耳菌株存在同种异名和同名异种现象。在实际生产中可选择亲缘关系较远的材料作为亲本菌株,以增加后代的遗传变异,从而选育出综合性状优良的肺形侧耳菌株。
参考文献:
冯伟林,蔡为明,金群力,范丽军,沈颖越,宋婷婷. 2014. 秀珍菇菌株主要农艺性状比较及ISSR分子标记鉴定[J]. 食用菌学报,21(2):14-18. [Feng W L,Cai W M,Jin Q L,Fan L J,Shen Y Y,Song T T. 2014. Differentiation of fifteen Pleurotus geesterani strains using ISSR markers and comparison of selected agronomic characteristics[J]. Acta Edulis Fungi,21(2):14-18.]
黄亦存. 1996. 高等担子菌的性非亲和性系统和体细胞非亲和性系统在遗传多样性保存中的作用[J]. 生物多样性,4(1):41-44. [Huang Y C. 1996. Role of sexual incompatibility and somatic incompatibility of Basidiomycetes in the conservation of their genetic diversity[J]. Chinese Biodiversity,4(1):41-44.]
贾定洪,王波,郑林用,黄忠乾,谭伟,甘炳成,彭卫红. 2013. 应用拮抗及ISSR方法鉴定袋料香菇菌株[J]. 西南农业学报,26(2):832-834. [Jia D H,Wang B,Zheng L Y,Huang Z Q,Tan W,Gan B C,Peng W H. 2013. Identification of isolate from Lentinus edodes cultivation bags with antagonism and ISSR[J]. Southwest China Journal of Agricultural Sciences,26(2):832-834.]
李红,李超,张敏. 2018. 金针菇菌株遗传多样性的RAPD分析[J]. 江苏农业科学,46(1):19-22. [Li H,Li C,Zhang M. 2018. Analysis of genetic diversity in Flammulina velutipes using RAPD markers[J]. Jiangsu Agricultural Sciences,46(1):19-22.]
李黎. 2011. 中國木耳栽培种质资源的遗传多样性研究[D]. 武汉:华中农业大学. [Li L. 2011. Studies on genetic diversity of Auricularia auricula-judae cultivated germplasm resources in China[D]. Wuhan:Huazhong Agricultural University.]
李维焕,蔡德华,郑芳,马茜楠,杨树德,高兴喜. 2010. 秀珍菇菌株的亲缘关系分析[J]. 食品科学,31(17):267-271.[Li W H,Cai D H,Zheng F,Ma Q N,Yang S D,Gao X X. 2010. Analysis of the phylogenetic relationships of Pleurotus pulmonarius strains[J]. Food Science,31(17):267-271.]
潘丽晶,沈汉国,陈继敏,张妙彬,蔡晓平. 2013. 荧光AFLP和拮抗实验对灵芝菌遗传多样性研究[J]. 中国食用菌,32(6):41-45. [Pan L J,Shen H G,Chen J M,Zhang M B,Cai X P. 2013. Analysis of genetic diversity of Ganoderma lucidum by fluorescent-AFLP and antagonistic reaction[J]. Edible Fungi of China,32(6):41-45.]
戚元成,张小强,高玉千,申进文,邱立友. 2010. 中国栽培糙皮侧耳品种体细胞不亲和性实验与 RAPD 分析结果的比较[J]. 菌物学报,29(3):379-388. [Qi Y C,Zhang X Q,Gao Y Q,Shen J W,Qiu L Y. 2010. Comparison between somatic incompatibility and RAPD analysis of cultivated Pleurotus ostreatus in China[J]. Mycosystema,29(3):379-388.]
谭伟,周洁,曹雪莲,苗人云,贾定洪,黄忠乾,甘炳成,彭卫红,郑林用. 2015. 姬菇258和金凤2-1杂交构建姬菇新菌株[J]. 西南农业学报,28(6):2655-2660. [Tan W,Zhou J,Cao X L,Miao R Y,Jia D H,Huang Z Q,Gan B C,Peng W H,Zheng L Y. 2015. Hybrid strains breeding by Pleurotus cornucopiae No.258 and Pleurotus sp. Jin feng-2-1[J]. Southwest China Journal of Agricultural Sciences,28(6):2655-2660.]
王锦锋,张健,林占熺. 2017. 应用拮抗、ITS和RAPD技术进行香菇属菌株分类[J]. 亚热带农业研究,13(2):132-138. [Wang J F,Zhang J,Lin Z X. 2017. Classification of Lentinus edodes strains by antagonistic effect,ITS and RAPD techniques[J]. Subtropical Agriculture Research,13(2):132-138.]
溫志强,熊芳,陈吉娜,朱坚,谢宝贵. 2011. 分子标记鉴别灰树花种质资源的研究[J]. 热带作物学报,32(7):1330-1336. [Wen Z Q,Xiong F,Chen J N,Zhu J,Xie B G. 2011. Identification of Grifola frondosa germplasm resources using molecular markers[J]. Chinese Journal of Tropical Crops,32(7):1330-1336.]
忻雅,阮松林,王世恒,马华升,王伟科,王淑珍. 2008. 基于RAPD和EST-SSR标记的秀珍菇菌株聚类分析[J]. 食用菌学报,15(4):20-25. [Xin Y,Ruan S L,Wang S H,Ma H S,Wang W K,Wang S Z. 2008. Cluster analysis of ten Pleurotus geesteranus strains based on random amplification of polymorphic DNA(RAPD) and expressed sequence tag-single sequence repeats(EST-SSR) markers[J]. Acta Edulis Fungi,15(4):20-25.]
杨军,徐吉,宋一鸣,朱坚. 2016. 耐高温型灰树花菌株筛选及供试灰树花亲缘关系分析[J]. 基因组学与应用生物学,35(2):170-179. [Yang J,Xu J,Song Y M,Zhu J. 2016. Screen of Grifola frondosa thermotolerant strains and analysis of the genetic relationship of candidate strains[J]. Genomics and Applied Biology,35(2):170-179.]
叶长文,谭海芹,吴应淼,吴敏,朱旭芬. 2013. 香菇135和9015品种遗传多样性的RAPD分子标记研究[J]. 浙江大学学报(理学版),40(2):230-234. [Ye C W,Tan H Q,Wu Y M,Wu M,Zhu X F. 2013. Study on the genetic diversity of Lentinula edodes 135 and 9015 by random amplified polymorphic DNA(RAPD) markers[J]. Journal of Zhe-jiang University(Science Edition),40(2):230-234.]
张安世,司清亮,齐秀娟,张中海. 2018. 猕猴桃种质资源的SRAP遗传多样性分析及指纹图谱构建[J]. 江苏农业学报,34(1):138-144. [Zhang A S,Si Q L,Qi X J,Zhang Z H. 2018. Genetic diversity and fingerprints of Actini-dia germplasm resource based on SRAP markers[J]. Jiangsu Journal of Agricultural Sciences,34(1):138-144.]
朱坚,卢启泉,谢宝贵. 2007. 秀珍菇种质资源分子多态性研究[J]. 菌物学报,26(S):226-231. [Zhu J,Lu Q Q,Xie B G. 2007. Molecular polymorphism in germplasm resource of Pleurotus geesteranus[J]. Mycosystema,26(S):226-231.]
Abidin M H Z,Abdullah N,Abidin N Z. 2016. Protective effect of antioxidant extracts from grey oyster mushroom,Pleurotus pulmonarius(Agaricomycetes),against human low-density lipoprotein oxidation and aortic endothelial cell damage[J]. International Journal of Medicinal Mushrooms,18(2):109-121.
Cortesi P,Milgroom M G. 1998. Genetics of vegetative incompatibility in Cryphonectria parasitica[J]. Applied and Environmental Microbiology,64(8):2988-2994.
Fonseca G G,Gandra E A,Sclowitz L F,Correa A P A,Levy J A. 2008. Oyster mushrooms species differentiation through molecular markers RAPD[J]. International Journal of Plant Breeding and Genetics,2(1):13-18.
Giovannetti M,Sbrana C,Strani P,Agnolucci M,Rinaudo V,Avio L. 2003. Genetic diversity of isolates of Glomus mosseae from different geographic areas detected by ve-getative compatibility testing and biochemical and mole-cular analysis[J]. Applied & Environmental Microbio-logy,69(1):616-624.
Wahab N A A,Abdullah N,Aminudin N. 2014. Characterisation of potential antidiabetic-related proteins from Pleurotus pulmonarius(Fr.) Quél.(greyoyster mushroom) by MALDI-TOF/TOF mass spectrometry[J]. BioMed Research International. doi:10.1155/2014/131607.
Williams C A,Wilson J F. 1966. Cytoplasmic incompatibility reactions in Neurospora crassa[J]. Annals of the New York Academy of Sciences,129(1):853-863.
Worrall J J . 1997. Somatic incompatibility in basidiomycetes[J]. Mycologia,89(1):24-36.
Xu W W,Li B,Lai E T C,Chen L,Huang J J H,Cheung A L M,Cheung P C K. 2014. Water extract from Pleurotus pulmonarius with antioxidant activity exerts in vivo chemoprophylaxis and chemosensitization for liver cancer[J]. Nutrition and Cancer,66(6):989-998.
(責任编辑 陈 燕)
