好氧迁移外源有机物转变成内源聚合物驱动内源反硝化
2020-03-23崔有为金常林
崔有为, 金常林
(北京工业大学环境与能源工程学院, 北京 100124)
生活污水脱氮能够有效解决水体富营养化,但是生活污水普遍存在碳氮比(ρ(C)/ρ(N))低、碳源不足导致的反硝化不充分的问题. 污水处理厂普遍通过采用投加外源碳源提高反硝化脱氮能力[1]. 现有的脱氮处理工艺使污水原水中的碳源在好氧条件下被氧化浪费,产生CO2温室气体,同时在反硝化阶段投加碳源增加了污水厂处理成本. 如何高效利用污水中的有机物用于脱氮是污水处理技术革新的目标. 如果能够避免原水中的有机物在好氧环境下被氧化消耗,而是迁移有机物到缺氧段作为反硝化碳源,将是高效利用原污水碳源、节约处理费用的有效途径[2]. 作为能源和碳源被微生物内源储存的物质有聚羟基脂肪酸酯(polyhydroxyalkanoates,PHA)和糖原(Glycogen,Gly). PHA积累菌是在非稳态生长条件下储存碳源的一类微生物. 驯化微生物内源储存采用盛宴饥饿机制(feast/famine,F/F)运行模式[3-4],即在好氧条件下充足碳源(盛宴)和缺乏碳源(饥饿)阶段交替运行,而且给予长期的饥饿期(即较低F/F). 这样,活性污泥在碳源非稳态供应下适应了内源优先储存的生长模式,激发了内源储存能力. F/F非稳态运行模式是一种高效的积累PHA的驯化方式,目前发现的具有PHA积累能力的微生物达300多种[5]. 除了PHA积累菌外,聚磷菌(phosphate accumulating organisms,PAOs)也可以积累PHA. PAOs在厌氧条件下将细胞中的聚磷水解为正磷酸盐释放胞外并获取能量,利用聚磷水解释放的能量吸收污水中挥发性脂肪酸合成贮能物质PHA[6-8]. 在好氧条件下,PAOs以氧为电子受体氧化胞内贮存的PHA,利用该反应产生的能量过量地从污水中摄取磷酸盐,将聚磷作为贮存物贮于胞内. 强化生物除磷(enhanced biological phosphorus removal,EBPR)工艺中还存在一类与PAOs竞争的聚糖菌(glycogen accumulating organisms,GAOs). GAOs在厌氧条件下降解积累的Gly,吸收外碳源合成PHA;在好氧条件下消耗PHA用于合成Gly. 因此,在好氧条件下能够内源储存碳源的主要微生物是PHA积累菌和GAOs.
关于PHA积累菌在全程好氧条件下合成和利用PHA的研究被广泛地报道. 最近的研究发现PHA积累菌除了利用分子态氧氧化储存的PHA产生能量外,还可以在缺氧条件下利用化合态氧氧化PHA产生能量. 这为实现好氧内源储存碳源,缺氧反硝化硝酸盐提供了理论基础[9-11]. 因此,本研究采用好氧/缺氧(aerobic/anoxic,O/A)工艺模式运行,探索水中碳源的好氧迁移用于缺氧反硝化的可行性. 研究重点考察内源物质储存和脱氮效率、内源反硝化计量,以及菌群结构的动态变化等关键问题. 虽然PHA积累菌和GAOs的生物学机制得到广泛研究,但是好氧条件下利用这些能够储存碳源的微生物迁移外源有机物在缺氧条件下内源反硝的研究还未见报道,这种碳源迁移的模式可以降低污泥和CO2的产量,同时实现污水中有机物的原位利用[12].
1 实验材料和方法
1.1 污水和接种污泥
实验原水采用北京工业大学家属楼化粪池处理出水. 生活污水的水质指标如表1所示. 活性污泥取自北京某污水处理厂. 该活性污泥具有较好的有机物去除、硝化、脱氮性能. 实验反应器接种初始污泥质量浓度在3 000 mg/L左右.

表1 生活污水水质条件
1.2 反应器的运行
试验采用序批式反应器(sequencing batch reactor,SBR)(见图1),有效体积为8 L,底面内直径为18.8 cm,高为40 cm. 生活污水化学需氧量(chemical oxygen demand,COD)质量浓度较低且波动较大,为了更好地实现与观察内源积累和反硝化过程,采用投加葡萄糖提高进水COD. 运行第1—40天,投加葡萄糖使进水COD质量浓度在300 mg/L左右,曝气量60 L/h. 为更明显观察迁移外碳源实现内源积累和内源反硝化脱氮效果,第41—180天增加葡萄糖投加量使进水COD质量浓度在500 mg/L左右,同时提高曝气量至120 L/h. SBR的启动采用O/A模式运行(见图1(b)). 每个周期按照进水、曝气、搅拌、沉淀、排水和闲置顺序进行. 沉淀结束排水1/2. O/A时长比为0.11,污泥龄(sludge retention time,SRT)为20 d,温度为(28±2) ℃. 第1—65天,系统存在部分硝化,好氧曝气1 h后进行缺氧搅拌;第66—180天,硝化消失,以溶解氧(dissolved oxygen,DO)跃升作为好氧盛宴结束的信号,此时停止曝气,内源积累达到最大值. 好氧盛宴曝气结束后投加0.293 g/L KNO3进行缺氧搅拌,好氧盛宴积累的内源聚合物有效用于反硝化脱氮过程,此时认为内源积累和内源反硝化脱氮启动成功.
1.3 内源反硝化计量实验
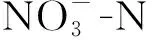
1.4 分析方法
1.5 微生物群体分析
为了测量微生物种群多样性和菌群结构的改变,采集接种原泥和驯化120 d的驯化污泥进行分子生物学分析,污泥样品在-20 ℃保存. 分析方法依据文献[14,16],使用E.Z.N.A.®固体DNA试剂盒(Omega Bi-TEK,Norcross,GA,美国)从样品中提取微生物DNA. 随后采用扩增细菌16S核糖体RNA基因的V4~V5区且带条形码的引物经PCR扩增. 每个样品设3个平行样,将对应的3个PCR扩增产物混匀为1个样本;胶回收阳性克隆条带用Tris 盐酸进行洗脱,根据2.0%琼脂糖电泳对各阳性胶回收产物的初步定量结果,采用蓝色荧光定量系统(QuantiFluorTM-ST, Promega, 美国)定量检测待测样PCR产物后,将各待测样的PCR产物根据测序量要求进行混合. 根据标准方案,在Illumina MiSeq平台上将纯化扩增子整合测序. 序列被保存到NCBI数据库,存档序列为SRP2905300.
1.6 计算
污泥中的PHA碳浓度计算公式为
c(PHA)=ρ(PHA)×0.046 5
(1)
污泥中的Gly碳浓度计算公式为
c(Gly)=ρ(Gly)×0.033 3
(2)
周期内好氧盛宴期内源聚合物(PHA+Gly)总产率计算公式为
(3)
式中:Δ(PHA+Gly)为周期内好氧盛宴期积累的内源聚合物浓度;ΔS为周期内好氧盛宴期降解的外源有机物浓度.
比反硝化速率(specific denitrification rate,SDNR)计算公式为
(4)
微生物细胞聚合物与反硝化脱氮间的ρ(C)/ρ(N)计算公式为
(5)
好氧盛宴期胞内聚合物的比生成速率(qP)、缺氧反硝化期间的胞内聚合物的比降解速率(rP)和有机物的比摄取速率(qS)计算公式分别为
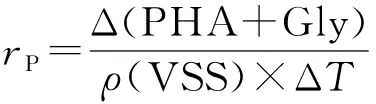
(6)
(7)
VSS、PHA以及Gly的分子式分别是CH1.8O0.5N0.2[3]、CH1.5O0.5[17]和CH2O(Gly检测实际以葡萄糖为标准曲线).VSS、PHA和Gly的摩尔质量分别是24.6、21.5、30.0 mg/mmol.
2 实验结果与讨论
2.1 系统性能
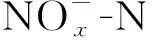
表2对比前期污泥和驯化稳定后的特性. 接种污泥前期PHA和Gly碳浓度很低,分别为0.17 mmol/L和0.85 mmol/L,内源产率仅为0.13±0.04. 经过长期驯化,污泥内PHA和Gly浓度得到显著增加,积累量分别平均为2.38 mmol/L和5.63 mmol/L,内源产率YP/S提高为0.56±0.09. 值得注意的是,内源积累物中Gly浓度得到了明显增加,最大Gly浓度由初期污泥的(8.53±0.87) mmol/L增加至(24.86±1.21) mmol/L. Gly积累量占总内源积累量的70.28%,成为主要的内源积累物质. 有研究发现在好氧F/F条件以葡萄糖为碳源下会导致Gly的积累[18],这可能也是本研究高Gly积累的主要原因. 对比驯化前后期,污泥合成的PHA中PHB和PHV质量分数没有发生显著变化,PHB质量占PHA总质量的80%左右.
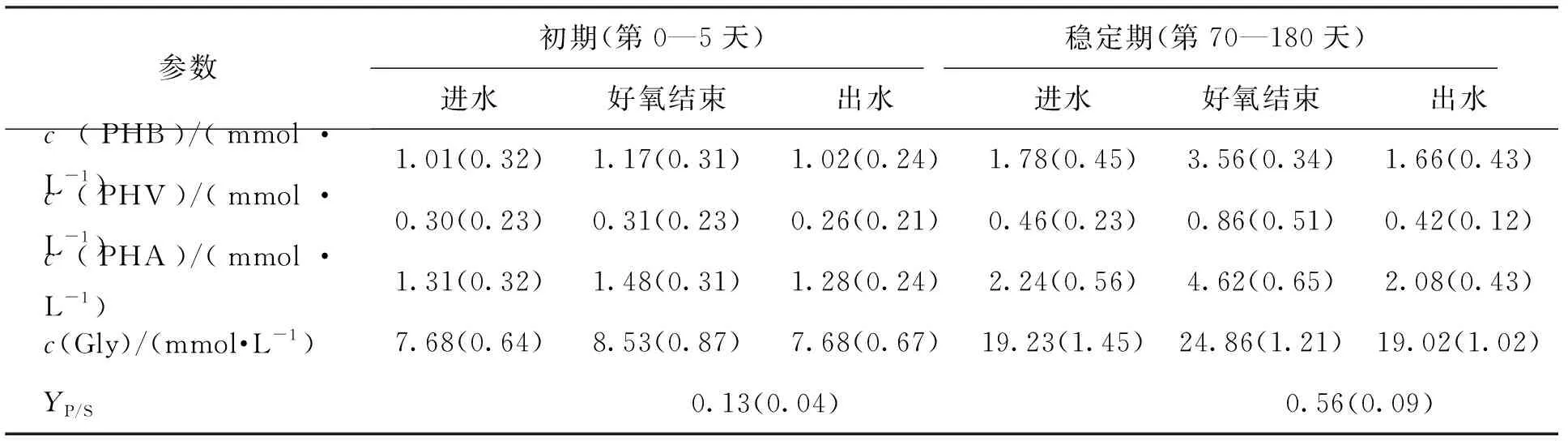
表2 PHA和Gly在系统运行初期和稳定期的浓度和内源聚合物平均总产率YP/S
注:进水、好氧结束、出水分别代表每周期此时的系统污泥中聚合物浓度,平均总产率为好氧盛宴结束后内源聚合物PHA和Gly的产率之和,括号内为标准偏差.
2.2 周期分析
2.3 内源反硝化计量
2.4 微生物种群变化
对接种原泥和驯化污泥进行16S rRNA MiSeq 焦磷酸测序分析,定性种群结构的变化(见图6). 门水平分析微生物菌群丰度(>1%)的变化(见图6(a)). 菌群结构发生了显著差异性变化,原泥优势菌门Proteobacteria由原来的45.45%降低为次级菌门,丰度为28.99%. TM7成为富集菌群的优势菌群,丰度为36.78%. Bacteroidetes丰度由原来的28.26%降低为16.40%. 此外,Chloroflexi、Chlorobi和Acidobacteria菌群丰度均呈现不同程度的降低,相反,Actinobacteria、OD1、GN02菌群丰度略微增加. Others由原来的6.88%降低为3.80%. 从纲水平看(见图6(b)),经过驯化Betaproteobacteria由系统最优势纲变为次纲,比例由31.22%降为18.05%. TM7- 3成为优势纲,由0升高到36.87%. Lgnavibacteria (1.93%~1.29%)、Deltaprotebacteria(5.48%~2.23%)、Gammaproteobacteria(4.94%~3.20%)、Anaerolineae(8.04%~2.83%)、Bacteroidia(8.15%~5.93%)、Sapeospirae (12.07%~6.13%)、Acidmicrobiia(3.54%~<1.00%)、Flavobacteriia(3.94%~<1.00%)、Sphingobacteriia(3.83%~<1.00%)丰度降低;Actinobacteria(<1.00% ~5.02%)、unclassified Bacteroidetes(<1.00%~3.75%)和Alphaproteobacteria(3.77%~4.67%)等菌纲丰度增加, Others由11.33%降低为7.18%.
在F/F非稳态条件、O/A工艺模式下,系统活性污泥微生物经过长期驯化菌群结构和多样性发生了显著变化. 驯化条件促进了TM7菌门微生物的生长,但是TM7菌群未曾报道具有内源积累和反硝化功能[24]. TM7菌门的增加相对降低了原泥中的优势菌门Proteobacteria和Bacteroidetes的丰度. Proteobacteria是污水处理厂常见的一种优势菌门微生物[25]. Proteobacteria和Bacteroidetes菌门部分微生物曾被报道具有内源积累和内源反硝化的功能[26-27]. Chloroflexi具有吸收外碳源的功能[28-29]. Bacteroidetes菌门下的unclassified Bacteroidetes和Proteobacteria菌门下的Alphaproteobacteria被富集并可能负责PHA的积累[26,30]. 此外,Proteobacteria菌门下的Dechloromonas曾同时被报道具有除磷功能[31-32].
3 结论
1) 在F/F为0.11的非稳态关键条件下,O/A工艺模式可以实现混合菌群高效率积累内源聚合物PHA和Gly驱动内源反硝化脱氮.
2) Gly是F/F非稳态条件、O/A工艺模式下微生物积累的主要内碳源,并主导了内源反硝化脱氮效果.
3) 在F/F非稳态条件下,O/A工艺模式改变了污泥的种群结构和丰度,TM7为系统优势菌门,unclassified Bacteroidetes和Alphaproteobacteria可能贡献了PHA的积累.
