奶牛子宫内膜组织α7-nAChR介导的抗炎效应
2020-03-13阿如娜曹金山贺鹏飞内蒙古农业大学兽医学院内蒙古呼和浩特010018农业部动物疾病临床诊疗技术重点实验室内蒙古呼和浩特010018
阿如娜,刘 科,陈 灰,曹金山,贺鹏飞 (1.内蒙古农业大学 兽医学院,内蒙古 呼和浩特010018;2.农业部动物疾病临床诊疗技术重点实验室,内蒙古 呼和浩特010018)
高产奶牛泌乳早期由于能量负平衡导致机体免疫力下降,奶牛分娩后生殖道细菌易于上行感染导致子宫内膜炎。每年我国分娩母牛子宫内膜炎发病率高达30%~40%,患牛人工授精次数增加,配种和受孕间隔延长,妊娠率、受孕率降低致使治疗费用及奶牛淘汰率增加,给我国奶牛业造成了巨大的经济损失[1]。当前对于子宫内膜炎治疗主要使用抗生素及前列腺素类药物,但由于抗生素药物残留,前列腺素对无活跃黄体奶牛疗效尚无定论且前列腺素频繁使用扰乱奶牛发情周期,因此开发新型抗菌药及抗炎药成为当前研究热点。α7-n ACh R 广泛的存在于神经细胞及非神经细胞,如星形胶质细胞、小胶质细胞、少突胶质前细胞及其他内皮细胞上[2]。最新的研究进展表明,激活免疫细胞和内皮细胞上的α7-n AChR 可以调节炎症和免疫反应,并且可能成为治疗一系列慢性难治性炎症疾病及炎性细胞因子相关疾病的新靶点。α7-n ACh R 由同源的α7 亚基组成的五聚体,其对Ca2+具有高度通透性,而作为第二信使的Ca2+在α7-n ACh R 发挥生物学效应中具有重要作用[3]。激活α7-n ACh R 可以通过胞内Ca2+信号诱导PKA 依赖的胞外信号调节激酶-1(ERK1)和ERK2的活化[4-5]、抑制IκB的磷酸化来抑制NFκB的活化,下调核转录因子NF-κB、Toll样受体4(TLR4)的表达及抑制促炎因子释放发挥抗炎作用[6-7];同时,激活α7-n ACh R 可以募集jak2蛋白形成异源二聚体,并触发信号转导子和转录激活子3(stat3)介导的信号转导,负性调节NF-κB 与DNA的结合,增强细胞因子信号传导抑制因子3(SOCS3)的活性参与抗炎效应[8]。在胶质细胞中,α7-n ACh R 可以上调环氧合酶2(cox-2)的活性促进前列腺素E2(PGE2)合成发挥抗炎作用[9]。
目前尚无子宫内膜α7-n ACh R 参与炎症调控的研究报道。课题组前期通过RT-qPCR 检测到奶牛子宫内膜组织α7-n AChR 表达,但其是否参与炎症调控尚不清楚。因此,本研究拟通过体外培养奶牛子宫内膜组织并建立LPS 炎症模型,应用α7-n AChR 特异性激动剂及抑制剂,使用RT-qPCR 检测炎症细胞因子il-1β、il-6、il-8、tnf-α、cox-2及mpges-1 m RNA 表达水平变化,初步阐明奶牛子宫内膜组织α7-n ACh R 的抗炎效应,为奶牛子宫内膜炎的治疗及新药开发提供新的思路。
1 材料与方法
1.1 试验药品胎牛血清(Inc,中国),DMEM/F-12(Gibco,美国),青霉素(Gibco,美国),链霉素(Gibco,美国),两性霉素B(GENERAY,中国),PNU-282987(MCE 生物公司,美国),甲基牛扁碱(MLA,MCE 生物公司,美国),脂多糖(LPS,Sigma,美国),AxyPrep Multisource Total RNA Miniprep Kit(Axygen Scientific,美国),SYBR Premix ExTaqⅡ(Ta KaRa,日本),Primer Script RT Master Mix(Ta KaRa,日本),引物均由Invitrogen公司合成。
1.2 试验仪器高速低温离心机,德国Beckman公司;高压自动蒸汽灭菌锅,日本Hirayama公司;低速台式离心机,中国上海安亭公司;超纯水仪,中国Biotech公司;-80℃低温冰箱,德国Thermo公司;-20℃低温冰箱,中国美菱公司;酶标仪,美国Bio-TEK 公司;制冰机,日本Panasonic公司;ABIVii ATM7型实时荧光定量PCR 仪,德国Life公司。
1.3 试验组织健康奶牛双侧子宫角采自内蒙古呼和浩特市某屠宰场,样品保存在无菌PBS缓冲液中,1 h内送回试验室,通过肉眼观察卵泡大小及黄体形态,选取发情前期的双侧子宫角部位组织进行组织培养。
1.4 组织培养将新鲜组织用PBS(含100 IU/m L双抗,2.5 mg/L两性霉素B)冲洗3遍,4℃保存1 h;无菌条件下,纵向剖开子宫角用弯剪刀和眼科镊子取下将含有子宫内膜上皮细胞的子宫内膜组织并剪成直径大小约为2 mm 的碎块,放入含有5 m L 培养液(DEME/F-12,20%胎牛血清,100 IU/m L 双抗,2.5 mg/L两性霉素B)的六孔板中,放入细胞培养箱中(5%CO2、37℃)培养,每天更换1次培养基,连续培养5 d直至组织生长稳定之后,将培养基换成无血清的培养基培养10~12 h,再换成含血清无双抗的培养基,进行药物处理8和12 h收集组织。
1.5 LPS作用时间筛选终质量浓度1 mg/L LPS刺激离体奶牛子宫内膜组织,分别培养2,4,8,12,24 h,应用RT-qPCR 技术评估组织中的il-6 m RNA的相对表达量。
1.6 药物处理把培养5 d的子宫内膜组织的培养基换成无双抗的的DMEM/F-12培养基,试验分为16组,分别为空白对照组、LPS组、LPS与α7n AChR激动剂组(PUN-282987 5,10,15 mmol/L),激动剂试验组分别设立5 mmol/L MLA 抑制剂组,每组6个重复,分别培养8和12 h,收集样品,进行炎性细胞因子检测。
1.7 总RNA提取及cDNA合成将各试验组组织称取40 mg,应用组织匀浆机进行研磨,按照说明书提供的方法使用总RNA 提取试剂盒提取组织总RNA,通过酶标仪检测总 RNA 的吸光度,D260 nm/D280 nm 值为1.9~2.0,质量浓度在500~1 000 mg/L可进行下一步试验;将总RNA 质量浓度调整到500 mg/L,然后按照说明书提供的方法使用反转录试剂盒mRNA 反转录合成cDNA。
1.8 RT-qPCR 检测炎性细胞因子RT-qPCR 反应体系为:SYBR Premix ExTaqⅡ10.0μL,上游引物0.4μL,下游引物0.4μL,Rox 0.4μL,cDNA 2.0μL,水6.8μL,总计20.0μL;反应条件为:95℃预变性30 s;95℃变性15 s,58℃退火34 s;72℃延伸30 s,共40 个循环。RT-qPCR 反应中所用的引物序列见表1。
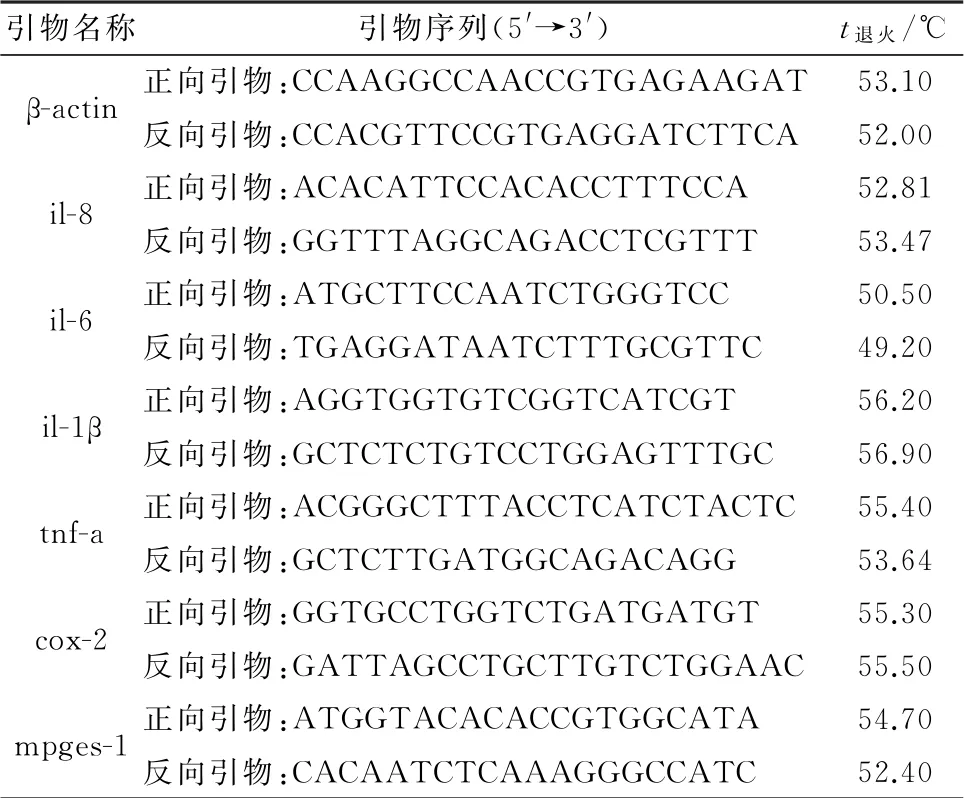
表1 基因序列
1.9 数据处理采用2-ΔΔC t法计算目的基因的相对表达量;使用SPSS15.0软件经ANOVA 统计学分析试验数据,结果表用±s x表示并用Graphpad Prism5.0软件进行绘图。
2 结果
2.1 奶牛子宫内膜组织LPS炎症模型时间优化由图1可知,LPS处理奶牛子宫内膜组织之后il-6 m RNA 相对表达水平在8和12 h最高,并在12 h达峰值,因此本研究选择8和12 h作为组织LPS刺激炎症模型作用时间。
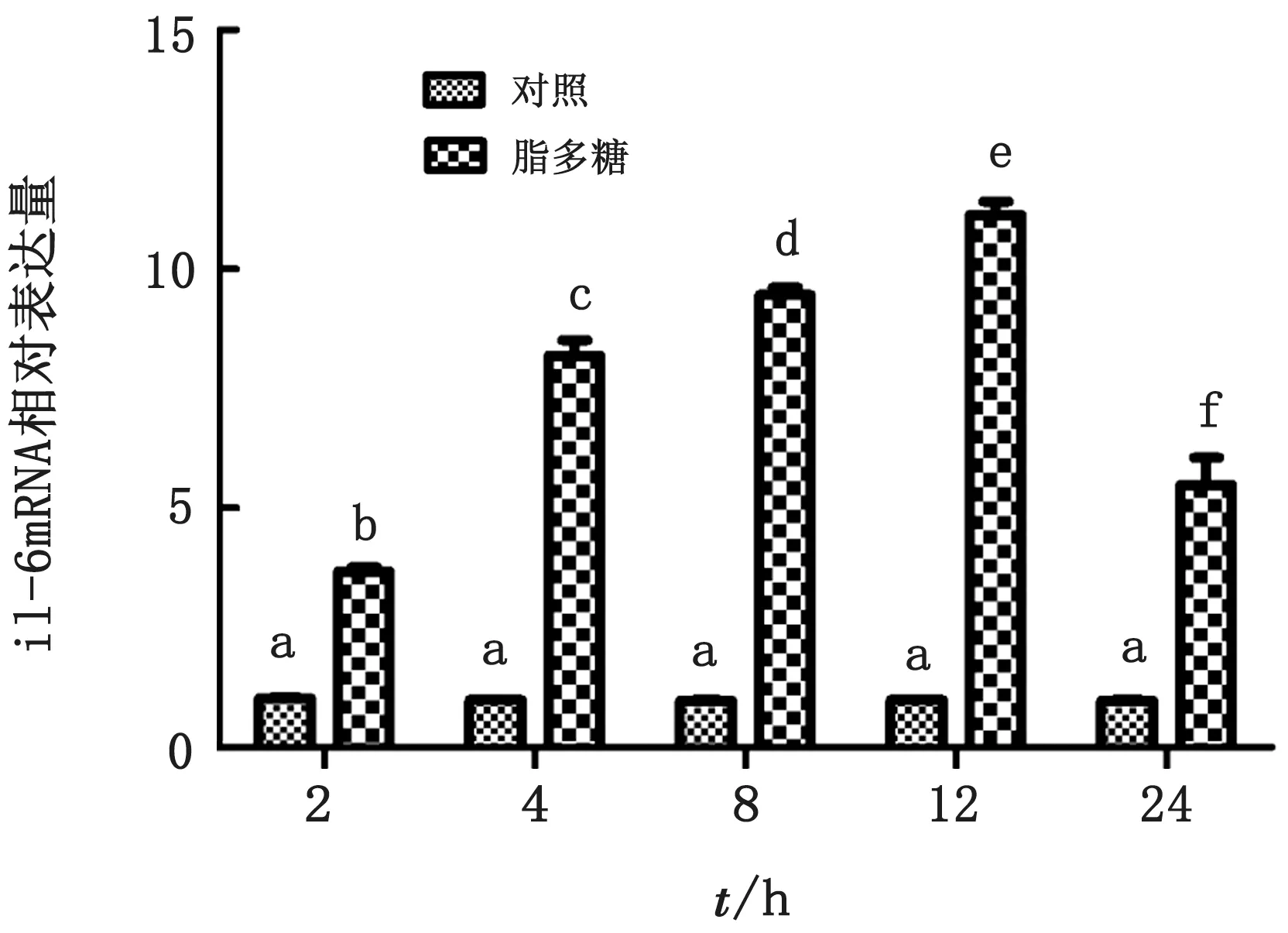
图1 LPS不同作用时间il-6 mRNA 相对表达水平 字母相异表示差异显著(P<0.05)。下同
2.2 奶牛子宫内膜组织α7-n AChR 介导的LPS炎症模型炎性细胞因子表达由图2可知,LPS刺激奶牛子宫内膜组织8 h能够显著上调促炎细胞因子il-1β、il-6和tnf-α m RNA 的 表 达,α7-n ACh R 特 异性激动剂PNU-282987在5,10,15 mmol/L均能显著抑制细胞因子的表达,α7-n ACh R 特异性拮抗剂MLA 能够明显抑制PNU-282987的作用。

图2 LPS作用奶牛子宫内膜组织8 h及不同浓度药物处理il-1β、il-6和tnf-αmRNA 相对表达水平 A.il-1βmRNA 相对表达水平;B.il-6 m RNA 相对表达水平;C.tnf-αm RNA 相对表达水平;1.CON;2.LPS;3.LPS+5 mmol/L PNU-282987;4.LPS+5 mmol/L PNU-282987+5 mmol/L MLA;5.LPS+10 mmol/L PNU-282987;6.LPS+10 mmol/L PNU-282987+5 mmol/L MLA;7.LPS+15 mmol/L PNU-282987;8.LPS+15 mmol/L PNU-282987+5 mmol/L MLA。下同
由图3A 可知,LPS刺激奶牛子宫内膜组织8 h能够明显上调炎症趋化细胞因子il-8 m RNA 的表达,α7-n AChR 特异性激动剂PNU-282987在5,10,15 mmol/L 均能显著抑制il-8 m RNA 的表达,α7-n AChR 特异性拮抗剂MLA 能够明显抑制PNU-282987 的 作 用;cox-2 和mpges-1 在PNU-282987在5和10 mmol/L浓度下与il-8具有相同趋势,但在15 mmol/L 浓度下的作用虽然明显低于LPS组,但MLA 不能明显逆转该效应(图3B,C)。
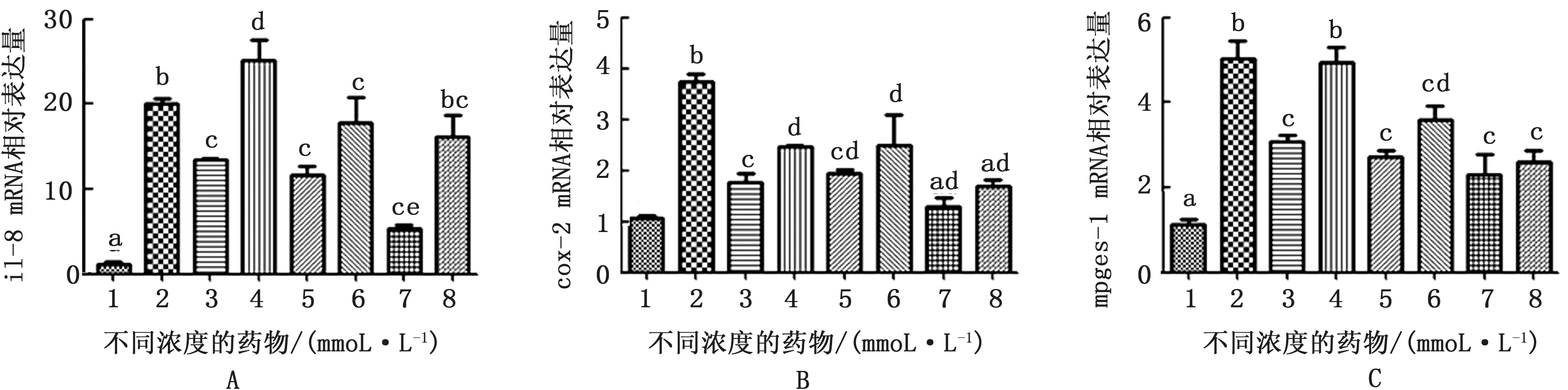
图3 LPS作用奶牛子宫内膜组织8 h及不同浓度药物处理il-8、cox-2和mpges-1 m RNA 相对表达水平 A.il-8 m RNA 相对表达水平;B.cox-2 m RNA 相对表达水平;C.mpges-1 mRNA 相对表达水平
由图4可知,LPS刺激奶牛子宫内膜组织12 h能够明显上调促炎细胞因子il-1β、il-6和tnf-αm RNA 的 表 达,α7-n ACh R 特 异 性 激 动 剂PNU-282987在5,10,15 mmol/L 均能显著抑制细胞因子的表达,PNU-282987在15 mmol/L时il-6和tnf-α的表达量未被相应的拮抗剂显著抑制,其他剂量下的PNU-282987均被α7-n ACh R 拮抗剂MLA 明显抑制,其他组α7-n ACh R 特异性拮抗剂MLA 均能明显抑制PNU-282987的作用。
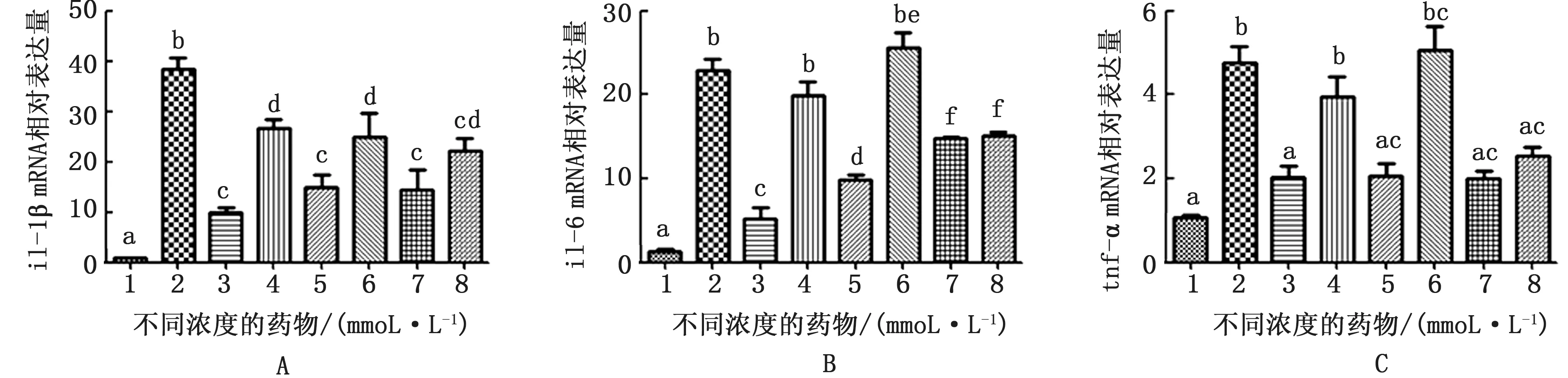
图4 LPS作用奶牛子宫内膜组织12 h及不同浓度药物处理il-1β、il-6和tnf-αm RNA 相对表达水平 A.il-1βm RNA 相对表达水平;B.il-6 m RNA 相对表达水平;C.tnf-αmRNA 相对表达水平
由图5可知,LPS刺激奶牛子宫内膜组织12 h能够促进趋化性细胞因子il-8的表达量,α7-n ACh R特异性激动剂PNU-282987在5,10,15 mmol/L均能显著抑制该因子的表达,除15 mmol/L 组外,α7-n ACh R 特异性拮抗剂MLA 能够明显抑制PNU-282987的作用;LPS可明显上调cox-2 和mpges-1 m RNA 表达,激动剂在5,10,15 mmol/L 均能显著抑制cox-2和mpges-1的表达,MLA 能够明显抑制激动剂上调cox-2和5 mmol/L mpges-1效应。
3 讨论
炎症对机体防御、组织再生及代谢调节具有重要作用,但过度的炎症反应则引起组织损伤。奶牛子宫内膜炎是产后奶牛高发疾病,革兰阴性菌感染是产后奶牛子宫内膜炎的主要病因,革兰阴性菌脂多糖(LPS)可被子宫上皮细胞模式识别受体(pattern recognition receptors,PRRs)识别,从而激活胞内信号通路诱导细胞因子、趋化因子的分泌促进炎症发生[10]。当前对奶牛子宫内膜炎防治措施主要是应用抗菌药及激素。由于细菌耐药性及药物残留,前列腺素类药物抗炎治疗在无黄体期效果存在争议,因此探寻新的抗炎靶位及开发新型药物是当前防治奶牛子宫内膜炎的研究热点。根据目前研究结果,无论天然药物还是微生物制剂都尚不能达到满意的临床效果,因此开发新型抗菌、抗炎药成为奶牛子宫内膜炎防治的研究重点[11]。“烟碱抗炎通路”在免疫反应及炎症级联中发挥重要作用,α7-n ACh R 在胆碱能抗炎通路中起到核心作用,激动α7-n AChR 可以显著抑制炎性细胞因子tnf-α、il-1β、il-6和il-18 的产生,是炎症疾病治疗的潜在靶点[12-13]。
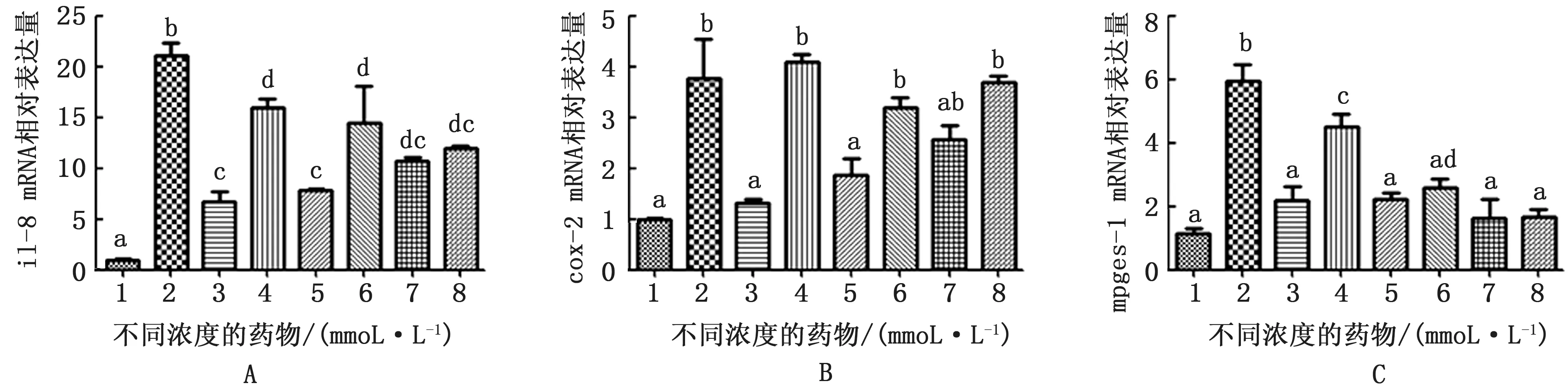
图5 LPS作用奶牛子宫内膜组织12 h及不同浓度药物处理il-8、cox-2和mpges-1 m RNA 相对表达水平 A.il-8 m RNA 相对表达水平;B.cox-2 mRNA 相对表达水平;C.mpges-1 m RNA 相对表达水平
Il-6是评价炎症模型常用的细胞因子[14],由图1可知,LPS处理奶牛子宫内膜组织8和12 h il-6 m RNA 的相对表达量最高,且12 h达峰值,因此本研究选择8和12 h作为LPS处理组织时间点。
在机体受到感染和损伤时,先天性免疫系统被激活并释放促炎性细胞因子如tnf-α、il-1β、il-6,它们引发的炎症反应在控制感染和促进组织修复中起重要的作用[15]。本研究结果表明,PNU-282987作为α7-n ACh R 特异性激动剂能够抑制由LPS致炎的奶牛子宫内膜组织分泌的炎性细胞因子tnf-α、il-1β、il-6的表达,而MLA 能抑制PNU-282987的作用从而上调促炎性细胞因子的表达。Il-8属于CXC 趋化因子亚家族,是激活和趋化中性粒细胞的主要趋化因子,主要趋化中性粒细胞迁移至血管内皮,促进其在炎症部位的聚集,促进内皮炎症的发展,加重内皮损伤[15-20]。LPS能诱导单核细胞、巨噬细胞、内皮细胞等合成分泌il-8[21-24]。在本试验中,PNU-282987能下调奶牛子宫内膜组织分泌的趋化性细胞因子的表达,而MLA 可逆转该效应。上述结果表明,奶牛子宫内膜α7-n ACh R 参与炎症反应,激动α7-n ACh R 具有抗炎效应。
Cox-2为诱导型酶,在炎症反应细胞如成纤维细胞、巨噬细胞和滑膜细胞中大量表达,促进PGE2的生成而参与炎症反应[25]。Mpges-1 与炎症关系非常密切,其在体外可被致炎因子如LPS、il-1β诱导而大量表达[26]。PGE2是一类在体内广泛存在的脂类调节因子,也是炎症反应中疼痛和发热的介质,参与炎症的病理、生理过程[27]。Mpges-1 是PGE2合成的终末限速酶,与cox-2 一起参与PGE2的合成[28],mpges-1水平的升高与cox-2表达和PGE2生成量的增加呈正相关。在本研究中,单独给予LPS诱导炎症后能够促进cox-2 和mpges-1 的表达,PNU-282987同时给药时能够明显抑制该效应,但在15 mmol/L试验组中,MLA 并不能完全逆转激动剂效应,或许与拮抗剂剂量不足有关,仍需要后续试验进一步验证。上述结果表明,奶牛子宫内膜组织α7-n ACh R 可能通过调控PGE2的合成参与炎症反应。
