刀鲚肌间骨新类型的发现
2020-03-04常永杰周佳佳张丽红孟佑廉高泽霞
常永杰 周佳佳 张丽红 孟佑廉 高泽霞 ,
(1. 华中农业大学水产学院, 农业部淡水生物繁育重点实验室/农业动物遗传育种与繁殖教育部重点实验室, 武汉 430070;2. 淡水水产健康养殖湖北省协同创新中心, 武汉 430070; 3. 池塘健康养殖湖北省工程实验室, 武汉 430070)
鱼类的肌间骨(Intermuscular bone, IB)又名肌间刺或肌间小骨, 是位于脊椎骨两侧肌隔中的硬骨小刺, 由肌隔结缔组织骨化而成的硬骨, 且通过结缔组织连在其他骨骼上[1—4]。目前被认知的鱼类肌间骨类型包括三类, 髓弓小骨、椎体小骨和脉弓小骨[5]。我国大宗淡水鱼类普遍存在肌间骨, 对人类的食用、出口和深加工造成很大困扰, 严重影响了其经济价值[6]。
近年来有关鱼类肌间骨的研究大多集中在肌间骨的数目及形态, 如国外Patterson和Johnson[5]对125属真骨鱼代表种的肌间骨进行了详细的观察和分析。国内对肌间骨形态学的研究集中于鲤科鱼类, 董在杰等[7]对鲢(Hypophthalmichthys molitrix)、鳙(Aristichthys nobilis)、团头鲂(Megalobrama amblycephala)和异育银鲫(Carassius auratus gibelio)的肌间骨进行了初步的探究; 柯中和等[8]、万世明等[3]、陈琳等[4]分别对鲢、团头鲂和黄河鲤(Cyprinus carpio haematopterus)肌间骨发育的形态学进行了观察。吕耀平等[9]对骨舌鱼总目、海鲢总目、鲱形总目和骨鳔总目等共计14种低等真骨鱼类的肌间骨形态学进行了系统研究, 认为肌间骨的形态是由简单向复杂演化, 然后逐渐退化。在肌间骨的发现和分类上面仅有秉志[1]提及, 现今延用的是Patterson和Johnson[5]的分类标准。
刀鲚(Coilianasus), 隶属鲱形目(Clupeiformes),鳀科(Engraulidae), 鲚属(Coilia), 是经济价值较高的鱼类[10,11], 其上颌骨较长, 向后延伸达到胸鳍基部,故又名长颌鲚。短颌鲚(C.brachygnathus)和刀鲚隶属于同一属, 其上颌骨较刀鲚短, 向后延伸不超过鳃盖骨。鲚虽然骨刺较多, 但肉嫩味美, 深受人们喜爱。目前对鲚属鱼类, 尤其是刀鲚种群的生物学参数、繁殖特征等方面有着大量的研究[12—14], 但针对鲚属鱼类肌间骨的研究却很少提及。本文主要对刀鲚和短颌鲚肌间骨的分布进行观察, 在人们普遍认知的鱼类三种类型肌间骨基础上, 发现了另外一种类型肌间骨的存在。此外本文对刀鲚肌间骨数目和长度进行研究分析, 作为对鱼类肌间骨形态学研究的补充。
1 材料与方法
1.1 实验材料
用于骨骼染色的刀鲚体重128 g, 体长34 cm; 短颌鲚体重45.8 g, 体长23 cm。形态解剖刀鲚共10尾,平均体重(49.58±6.16) g, 平均体长(24.39±4.39) cm。以上用于染色的刀鲚和短颌鲚于2017年8月份从鄱阳湖捕捞获得, 解剖的刀鲚个体于2018年3月份购于江苏省启东市连兴港码头。刀鲚胸鳍上部游离丝状鳍条5—6枚, 脊椎骨77—82根, 臀鳍甚长, 与尾鳍基相连; 上颌骨游离, 超过鳃盖后缘, 延伸至胸鳍基部。短颌鲚基本特征与刀鲚相似, 但其上颌骨较刀鲚短, 向后延伸不超过鳃盖骨。因刀鲚和短颌鲚活鱼运输较困难, 所购买的实验鱼均为冰鲜鱼。
1.2 刀鲚物种鉴定
提取刀鲚鳍条组织DNA, 根据NCBI数据库中已有的刀鲚COⅠ基因序列(GBGCA10305-15; Gen-Bank: KJ710626), 利用Primer Premier 5.0设计PCR引物(COI-F: 5′-TATTTAGTATTCGGT GCCTG-3′; COI-R: 5′-TGCTACTTCTCGTTTGGC-3′), 送至武汉擎科生物技术公司合成; 以实验鱼DNA为模板, PCR扩增目的基因(COⅠ), 并送至武汉昆泰锐生物公司测序。PCR反应产物测序后获得实验鱼的COⅠ基因序列1304 bp, 与NCBI数据库中已有的刀鲚COⅠ基因序列匹配度高达99%。结合捕获地点和形态学特征可确定实验鱼为刀鲚。
1.3 整体骨骼染色方法
骨骼染色方法在白凤熙[15]染色方法的基础上略有改动, 具体方法如下: (1)无水乙醇脱水固定1周;(2)将标本转移至2% KOH和3% H2O2的混合溶液中漂白3h; (3)将漂白的标本转移至2% KOH溶液中透明; (4)3h后去除鳞片和内脏, 继续透明(透明时间以鱼体隐约半透明为佳, 约6h); (5)经透明的标本转移至新的茜素红染色液(1 g茜素红溶于100 mL 1%KOH溶液中配制成染色液), 染色时间约6h; (6)染好色的标本依次在20%、50%和100%甘油进行梯度透明。最后对染色结果用数码相机(佳能IXUS170)进行拍照观察。
1.4 肌间骨数目、形态的观察和测定
实验鱼经纱布擦干鱼体后进行体重与体长的测量。然后用纱布将整条鱼包裹好, 置于沸水中煮10—15min, 取出, 待其自然冷却后去除纱布, 用镊子小心剥去皮肤和肌肉, 根据刀鲚染色结果来挑取肌间刺, 并依次从前往后排列, 然后用数码相机(佳能IXUS170)拍照, 并用游标卡尺(广陆0—200, 精度0.01 mm)测量肌间骨长度。
1.5 肌间骨组织学观察
取刀鲚髓弓小骨、脉弓小骨、背部上方和腹部下方的肌间骨样品(包含骨组织周围的结缔组织)于4%多聚甲醛固定, 并进行后续的组织学分析。固定后的样品于双蒸水中漂洗后转于EDTA脱钙液(谷歌生物公司)中进行脱钙处理, 在室温下肌间骨脱钙12h; 脱钙完成后将样品进行一系列脱水处理后用石蜡进行组织包埋, 并进行连续切片(5 μm);将切片于二甲苯中脱蜡透明、梯度酒精复水后将其分成两份, 分别进行茜素红染色和阿利新蓝染色。其中茜素红染色过程为: 将切片放于茜素红染液中浸染5min, 自来水洗去切片浮色后转置于60℃烘箱烘片30min, 烘片结束后于二甲苯透明, 最后中性树脂封片; 阿利新蓝染色过程为: 阿利新蓝染液中浸染15min, 置于水中洗去浮色后转置于核固红复染5min, 依次对切片进行水洗、二甲苯透明处理, 最后用中性树脂封片。
2 结果
2.1 肌间骨形态及分布
对刀鲚和短颌鲚整体骨骼染色结果观察发现,刀鲚和短颌鲚不仅有髓弓小骨、脉弓小骨和椎体小骨, 在其背部上方和腹部下方也存在肌间骨, 分别存在于背部上方和腹部下方肌膈的皱褶部(图1)。解剖后观察显示刀鲚肌间骨的形态有“1”形、“卜”形、“y”形、一端多叉形和“(”形5种类型。髓弓小骨和脉弓小骨的形态较复杂, 主要有“1”形、“y”形、“卜”形, 少量“(”形和“一端多叉”形(图2); 椎体小骨的形态仅有“1”形; 背部上方肌间骨形态简单,多数为“(”形, 分布于背鳍下方至尾鳍处; 腹部下方肌间骨多为“1”形, 少数是“(”形, 分布于肛门至尾鳍处; 背部上方和腹部下方肌间骨较其他部位的肌间骨短而细(图2)。
2.2 肌间骨的数目和长度
刀鲚的肌间骨的数目见表1。个体肌间骨总数范围为492—543, 平均为521; 左侧肌间骨范围为233—274, 平均为256; 右侧肌间骨范围为239—301, 平均为265; 左右两侧肌间骨数目差异不显著(P>0.05)。背部上方肌间骨、椎体小骨、腹部下方肌间骨与髓弓小骨和脉弓小骨在数目上存在显著差异(P<0.05), 背部上方肌间骨、椎体小骨、腹部下方肌间骨三者之间数目差异显著(P<0.05), 髓弓小骨与脉弓小骨数目差异不显著(P>0.05)。
刀鲚各种类型肌间骨的长度如表2所示。背部上方肌间骨的长度为(1.44±0.16) cm; 髓弓小骨的长度为(2.09±0.24) cm; 椎体小骨的长度为(1.02±0.17) cm;脉弓小骨的长度为(2.13±0.19) cm; 腹部下方肌间骨长度为(1.08±0.12) cm。髓弓小骨和脉弓小骨的长度显著长于背部上方肌间骨(P<0.05)和腹部下方肌间骨(P<0.01), 背部上方肌间骨的长度显著长于椎体小骨和腹部下方肌间骨(P<0.05)。不同部位肌间骨长度相关性分析结果显示背部上方肌间骨的长度与腹部下方肌间骨(r=0.744)和髓弓小骨(r=0.762)呈显著正相关(P<0.05), 髓弓小骨的长度与腹部下方肌间骨长度的相关性极显著(P<0.01), 其他部位的肌间骨之间没有显著的相关性(表3)。此外, 不同部位的肌间骨与个体体长和体重也存在一定的相关性分析(表4), 其中体长与脉弓小骨(r=0.967)长度存在极显著相关性(P<0.01), 体重与脉弓小骨(r=0.742)的相关性显著(P<0.05), 与其他部位肌间骨长度没有显著的相关性。
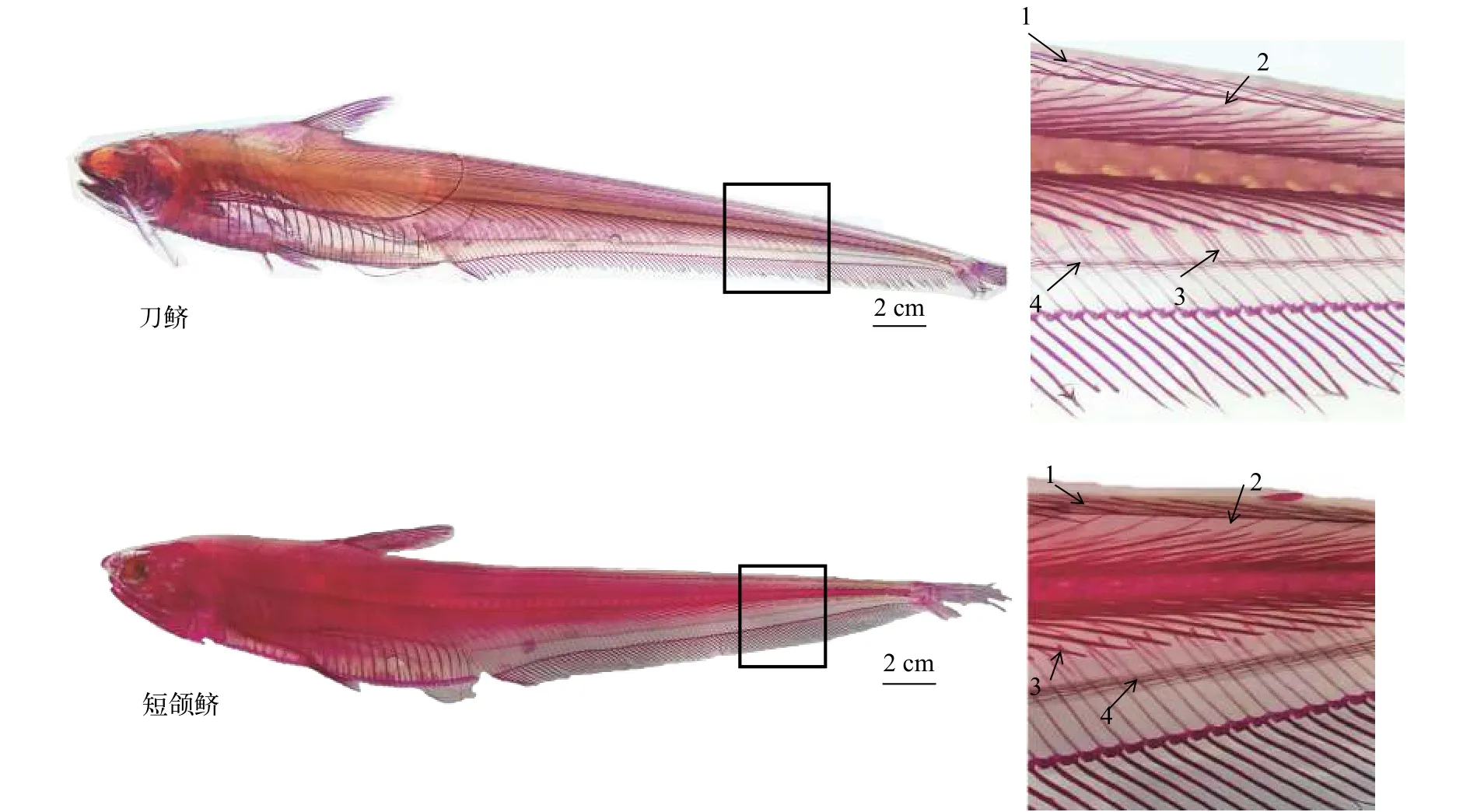
图1 刀鲚和短颌鲚整体骨骼染色肌间骨结果图Fig. 1 Whole-mount staining of skeleton for intermuscular bones in C. nasus and C. brachygnathus
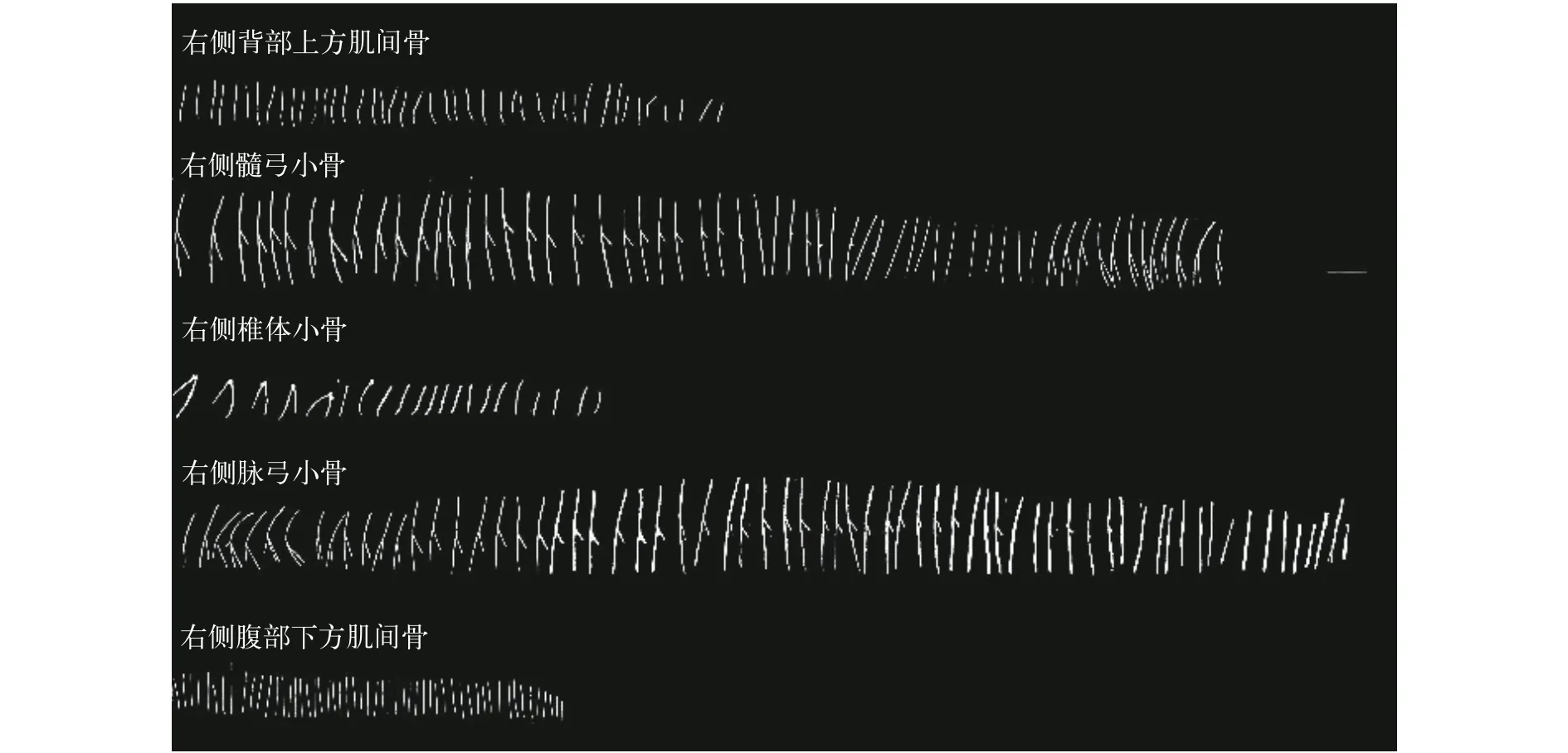
图2 刀鲚右侧肌间骨形态图Fig. 2 The morphology of IBs in C. nasus

表1 刀鲚肌间骨的数目Tab. 1 The number of IBs in C. nasus
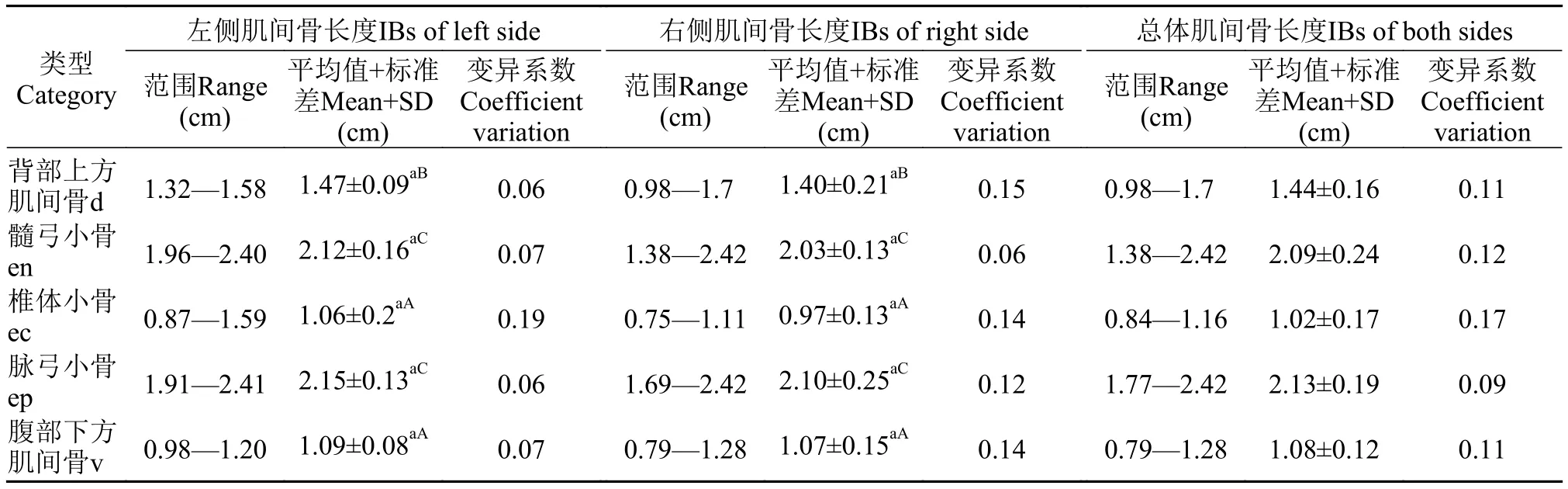
表2 刀鲚肌间骨的长度Tab. 2 The length of IBs in C. nasus
2.3 组织学结构
为了解刀鲚不同部位肌间骨(椎体小骨除外)的组织学结构及其差异, 本研究采用茜素红与阿利新蓝两种染色方法分别对其切片组织进行染色分析,茜素红可使钙化结节或硬骨组织染成红色, 阿利新蓝可使软骨组织、结缔组织染成蓝色。染色结果显示刀鲚的背部上方肌间骨(图版Ⅰ-1—4)、髓弓小骨(图版Ⅰ-5—8)、脉弓小骨(图版Ⅰ-9—12)和腹部下方肌间骨(图版Ⅰ-13—16)均被茜素红染成红色, 而阿利新蓝未使肌间骨组织着色, 但其四周结缔组织被染色为蓝色, 表明各种类型的肌间骨均为钙化的硬骨组织。肌间骨外周结缔组织染色结果表明在髓弓小骨和脉弓小骨中, 每根肌间骨并不是独立存在的, 而是由肌膈等结缔组织连接在一起,形成一个网状的结构(图版Ⅰ-7、11); 在背部上方肌间骨(图版Ⅰ-3)和腹部下方肌间骨(图版Ⅰ-15)中,各肌间骨之间未发现明显互相连接的结缔组织。
3 讨论
Patterson和Johnson[5]根据鱼类肌间骨附着的部位把其分为三类, 包括髓弓小骨、椎体小骨和脉弓小骨。本研究以刀鲚为主要的研究对象, 鱼体解剖和整体骨骼染色结果都显示, 刀鲚除了包含Patterson和Johnson[5]描述的三种肌间骨以外, 还有另外两种在国内研究中未被提起的背部上方和腹部下方肌间骨的存在。Chapman[16]在鲱科鱼类中也发现有两类额外的独立肌间骨, 背部分布于除前5—6个肌节之外的肌节中, 腹部分布于从肛门至尾鳍基部的肌节中, 这两类肌间骨都存在于肌膈的皱褶部, 将其统一命名为肌骨杆(Myorhabdoi)。本研究在刀鲚和短颌鲚中发现的背部上方肌间骨和腹部下方肌间骨与Chapman[16]描述的肌骨杆位置相似, 因此将本研究中发现的背部上方肌间骨和腹部下方肌间骨定义为背上肌骨杆和腹下肌骨杆。Chapman[16]认为这种类型的肌间骨是鲱科鱼类的典型特征, 鳀科鱼类没有, 这与本研究发现的鲱形目鳀科的刀鲚相悖。吕耀平等[9]报道的刀鲚肌间骨只有三种类型, 未提及背上和腹下肌骨杆的存在,因此其报道的肌间骨数目仅有221根, 较本研究的521根少很多。本研究发现的刀鲚椎体小骨的平均数目为42根, 这与蒋燕等[17]的44根接近。刀鲚背上和腹下肌间骨类型(统称肌骨杆)的发现丰富了国内鱼类肌间骨类型的基础资料。

表3 刀鲚肌间骨数目和长度的相关性分析Tab. 3 The correlation between IB’s number and IB’s length in C. nasus from different parts
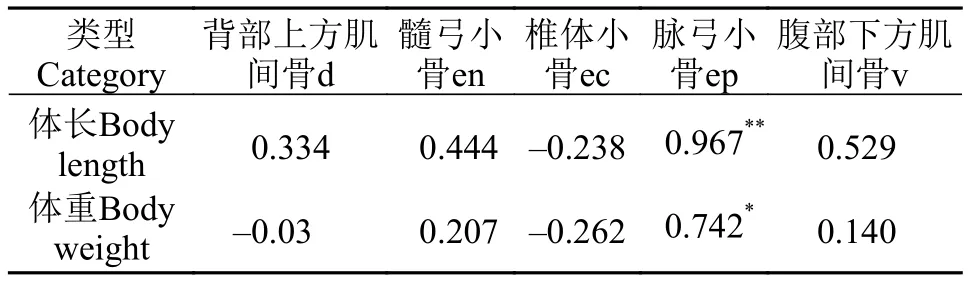
表4 刀鲚体长体重与肌间骨长度的相关性分析Tab. 4 The correlation of IBs’ length with body length and body weight in C. nasus from different parts
关于鱼类肌间骨的形态类型, 苏锦祥等[18]曾把鲢的肌间骨分为4种类型: “1”形、“卜”形、“y”形和树枝形。根据低等真骨鱼类肌间骨形态的复杂程度, 吕耀平等[9]为便于分析各种形态肌间骨演化规律, 将其归纳为7种类型(“ I”形、“卜”形、“Y”形、一端多叉形、两端两分叉形、两端多叉形、树枝形)。万世明等[3]将团头鲂肌间骨形态分为6种形态,分别是“1”形、“卜”形、“y”形、一端多叉形、两端多叉形、“(”形。董在杰等[7]通过对几种常见的鲤科鱼类肌间骨的研究把其主要归于两类: “1”形和“y”形, 其他形态均是在其基础上分化的。本文采用万世明等[3]的分类方法, 把刀鲚肌间骨分为从最简单的“1”形到最复杂的一端多叉形在内的5种形态。刀鲚肌间骨形态的分类研究已有文献报道, 但可能由于采取的分类标准不同, 本文对刀鲚肌间骨的形态比吕耀平等[9]的分类多出一种“(”形。刀鲚的椎体小骨形态相较于海鳗(Muraenesox cinereus)和鲥(Tenualosa reevesii)简单, 仅有“1”形, 与蒋燕等[17]的研究一致。刀鲚肌间骨解剖结果可以看出靠近尾部的肌间骨形态结构较为简单, 中间部位的形态较为复杂且较两端长, 这可能与鱼体不同部位的形态结构及机体自身的运动相关。
肌间骨是由肌隔结缔组织骨化而成的硬骨。本研究各种类型的肌间骨均被茜素红染成红色, 而阿利新蓝未使肌间骨组织着色, 但其四周结缔组织被染色为蓝色, 组织学结果证实了刀鲚各种类型的肌间骨均为钙化的硬骨组织。哺乳动物肌膈分化的相关研究表明, 肌膈中的间充质干细胞可以分化为肌腱源性干细胞, 而肌腱源性干细胞可以分化为成骨细胞、软骨细胞、脂肪细胞、成纤维细胞和肌腱细胞[19]。因此, 肌间骨可能是由间充质干细胞首先分化为肌腱源性干细胞, 之后在肌腱源性干细胞分化相关基因的作用下分化为成骨细胞, 进而形成骨细胞, 最终形成肌间骨, 但其具体的形成机制还有待进一步研究。


图版Ⅰ 刀鲚背部上方肌间骨(1—4)、髓弓小骨(5—8)、脉弓小骨(9—12)、腹部下方肌间骨(13—16)组织的茜素红(1, 2, 5, 6, 9, 10,13, 14)和阿利新蓝(3, 4, 7, 8, 11, 12, 15, 16)染色结果PlateⅠ Histological analysis of different IBs in C. nasus, transverse sections (1-16). transverse sections, stained with Alizarin red (1, 2, 5,6, 9, 10, 13, 14) and Alcian blue (3, 4, 7, 8, 11, 12, 15, 16), showing the dorsal IBs (1—4), epineurals (5—8), epipleurals (9—12), ventral IBs (13—16) of C. nasus黑色圆圈为各类肌间骨所在位置, 偶数字母(2, 4, 6, 8, 10, 12, 14 and 16)图片为对应奇数字母(1, 3, 5, 7, 9, 11, 13 and 15)图片中黑色实心圆圈部位的放大图The IBs indicated by black circle in odd number (1, 3, 5, 7, 9, 11, 13 and 15) were magnified in even number (2, 4, 6, 8, 10, 12, 14 and 16)
