草鱼肌间骨形态学观察与分析研究
2019-12-13段晓迪王蓉董传举
段晓迪, 王蓉, 董传举
(河南师范大学水产学院,河南新乡453007)
中国具有悠久的水产养殖历史,养殖总产量位居全球领先地位。近年来,全国水产品总产量逐年攀升,2017年淡水养殖产量已高达2905.29万吨,占水产品总量的45%以上[1]。在淡水养殖鱼类中,草鱼(Ctenopharyngodon idellus)的养殖规模最大且区域最广,其产量高达534.56万吨[1,2]。但肌间骨(Intermuscular bone,IB)的存在严重影响了草鱼的食用及深加工,使其不能达到国际优质鱼类的品质标准,进而极大的影响了草鱼的市场消费及经济价值[3]。
肌间骨(又称肌间刺或肌间小骨)是散布于鱼类肌间隔中的硬骨小刺,直接由间叶细胞骨化形成膜性硬骨[4,5],其发育过程无软骨阶段[6],通俗说法称之为鱼刺,解剖学上称之为种籽骨。呈针刺状,有分杈分枝现象,存在于轴上肌与尾部轴下肌之中,通过结缔组织系着穿过的肌节。依据肌间骨的着生部位,可将其分为三类:(1) 连接在髓弓上的髓弓小骨(epineural bone);(2)连接在椎体上的椎体小骨(epicentral bone);(3)连接在腹肋或脉弓上的脉弓小骨(epipleural bone)[4,7]。肌间骨仅存在于真骨鱼类中,低等真骨鱼类普遍存在肌间骨,而高等真骨鱼类几乎不存在肌间骨[8]。国内对于肌间骨的研究则主要集中在鲤科鱼类,如董在杰等对鲢(Hypophthalmichthys molitrix)、鳙(Aristichthys nobilis)、团头鲂(Megalobrama amblycephala) 和异育银鲫(Carassius auratus gibelio)肌间骨进行了初步分析[7];吕耀平等系统的对草、鲢、鳙、瓯江彩鲤(Cyprinus carpiovar. color)等鱼类肌间骨进行比较[9,10];陈琳等对黄河鲤(Cyprinus carpio haematopterus)、鲢、团头鲂、淇河鲫(Carassius auratusin Qihe river)的肌间骨形态发生、数目及分布等进行了详细报道[11-14]。除了形态学的研究,对于肌间骨的分子学研究也有了较大进展,例如:Wan等通过对团头鲂microRNA(miRNA)及转录组分析揭示了肌间骨发生的分子机制,并初步确定了与其发育相关的miRNA/基因相互作用通路[15,16];Zhang等通过对团头鲂Bmp基因家族的研究,发现Bmps可能是鱼类肌间骨形成过程中的启动子或抑制剂[17];Lv等研究表明MsxC可能在唇鳍骨(Hemibarbus labeo)的肌间骨发育中发挥作用[18]。尽管对于肌间骨的研究有了一定的进展,但肌间骨发生的分子调控机制比较复杂,相关机制的研究尚需深入开展。
草 鱼 属 鲤 形 目 (Cypriniformes), 鲤 科(Cyprinidae),雅罗鱼亚科(Leuciscinae),草鱼属(Ctenopharyngodon),分布于我国各大水系,通常喜居于中下层水域或水草茂盛区域,喜欢群体觅食,因其能迅速清除水域中各种水草,而有“开荒者”之称。目前对于草鱼肌间骨系统的形态学分析较少[10,19]。本研究以“四大家鱼”之一的草鱼为研究对象,对同一水域相同规格的草鱼肌间骨进行形态学观察及分析,为揭示草鱼肌间骨的演化途径、分子发生机制及高品质草鱼的培育提供初步的形态学基础。
1 实验材料和方法
1.1 实验材料
本实验所用草鱼均为随机采样野生二倍体,采样地点为:东经E113.79,北纬N27.57,采集后保存于-20℃冰箱。取3尾草鱼,分别命名为草鱼01,草鱼02及草鱼03(以下以样本号“01”,“02”,“03”表示)。
1.2 草鱼形态学测量
草鱼外部形态的测量采用传统方法。首先将-20℃冷冻保存的草鱼放置室温解冻,完全解冻后,用直尺及游标卡尺进行全长、体长、头长、体宽、体高等形态学数据的测量,并测量体重,记录并保存数据(表1)。

表1 草鱼形态学测量及肌间骨总数Tab.1 Morphological measurements and number of total intermuscular bones of grass carps
1.3 肌间骨观察方法
草鱼形态学指标测量完成后,去除鳞片和内脏,用纱布将鱼身包裹,沸水煮15 min后取出。去纱布,将鱼体放至解剖盘冷却至室温后进行解剖。将鱼体分为以脊柱为界的轴上部、轴下部及尾鳍基部三部分,用镊子从尾鳍基部向头部依次剥离肌肉,并按序取出肌间骨,按肌间骨左侧在上、右侧在下的顺序将其摆放至黑色背景卡片。对已排序的肌间骨形态进行分类,拍照记录并固定。
2 结果
2.1 草鱼的形态学测量及肌间骨总数
所采集的野生二倍体草鱼形态学测量数据见表1。全长在44.6-46.3 cm之间,平均值为45.7 cm;体长在37.4-39.2 cm之间,平均值为38.3 cm;头长在7.8-8.4 cm之间,平均值为8.1 cm;体宽在6.1-6.3 cm之间,平均值为6.2 cm;体高在8.6-9.6 cm之间,平均值为9.1 cm;体重在1158-1199 g,平均值为1182.3 g。三尾草鱼的肌间骨总数分别为131、129、134,平均值为131根(图1-3)。

图1 草鱼01形态图Fig. 1 Morphology of grass carp 01

图2 草鱼02形态图Fig. 2 Morphology of grass carp 02

图3 草鱼03形态图Fig. 3 Morphology of grass carp 03
2.2 草鱼肌间骨的形态
鱼类肌间骨有多种形态,根据其形态复杂程度可将其总结为7种类型:①“I”形(无分叉),②“卜”形(一端两分叉且不等长),③“Y”形(一端两分叉且等长),④一端多叉形(一端具3个及以上分叉),⑤两端两分叉形(两端均有两分叉),⑥两端多叉形(两端均有两个及以上分叉),⑦树枝形(两端均有4个以上的分叉)[10]。实验结果显示草鱼中不含树枝形肌间骨,但含有“L”形肌间骨。参考上述分类标准,我们将实验所用草鱼肌间骨分为“I”形、“L”形、“Y”形、“卜”形、“Y”形、一端多叉形、两端两分叉形及两端多叉形7种类型(图4)。

图4 7种肌间骨形态类型Fig.4 Seven shapes of IB morphology
实验所用的野生草鱼不同形态类型的肌间骨数目统计如表2所示。髓弓小骨上述7种形态均存在,其中“I”形平均37.3根,“L”形平均0.7根,“卜”形平均12.0根,“Y”形平均22.3根,一端多叉形平均2.0根,两端两分叉形平均10.3根,两端多叉形平均4.7根;脉弓小骨含“I”形、“L”形、“卜”形、“Y”形、一端多叉形5种肌间骨,其平均值依次为27.3根,3.0根,4.0根,6.7根,0.7根。髓弓小骨及脉弓小骨均以“I”形肌间骨数量居多,“Y”形数量次之,总体上髓弓小骨形态较为复杂。
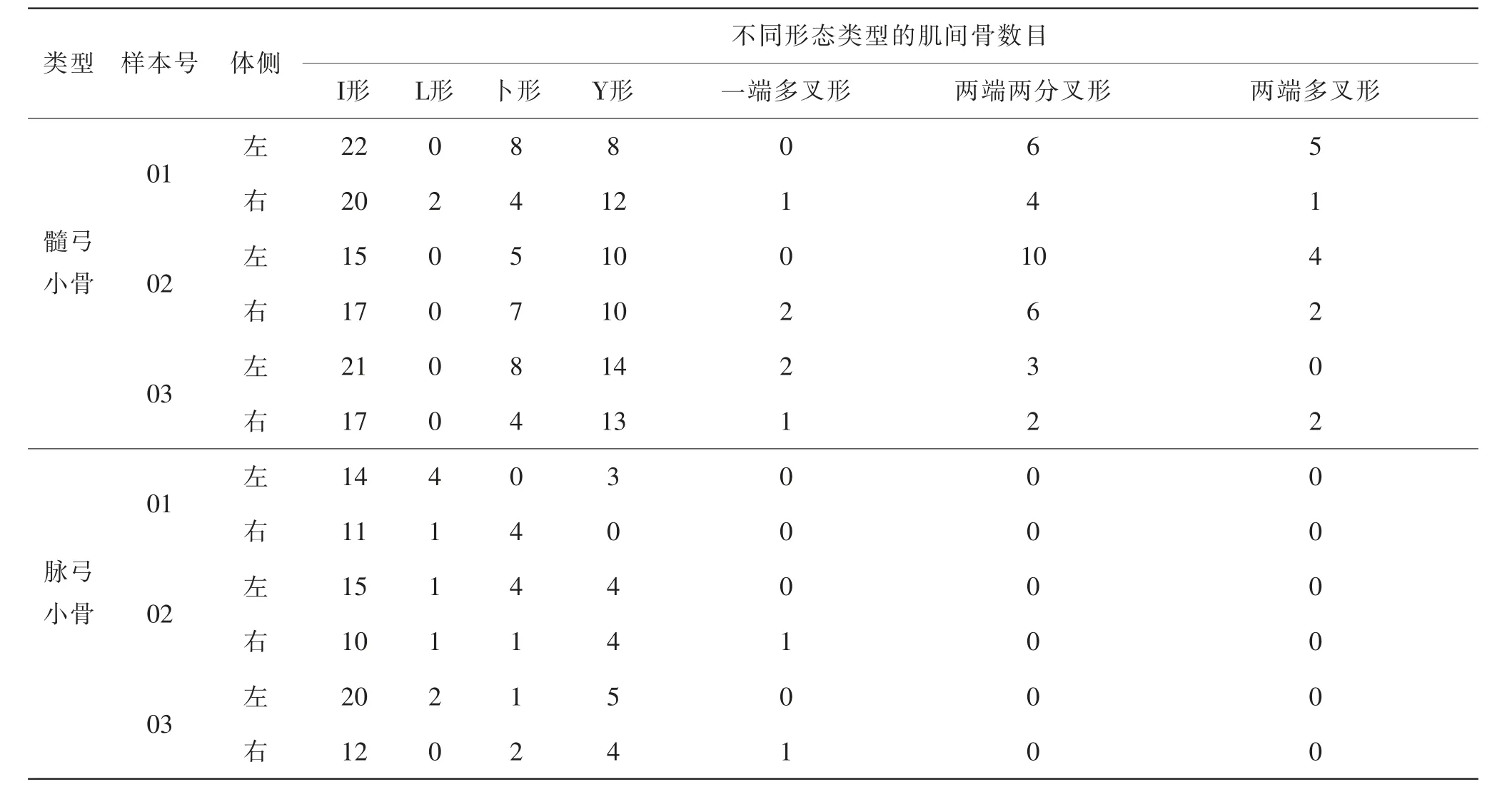
表2 草鱼不同形态类型的肌间骨数目统计Tab. 2 Statistics of the number of intermuscular bones in grass carp of different morphological types
2.3 草鱼肌间骨在不同部位的分布
草鱼成体两侧肌间骨的分布如图5-7所示,按照髓弓小骨及脉弓小骨在鱼体内的位置从头至尾按序摆放。按照鱼体左右两侧区分,01号草鱼左侧肌间骨为70根,右侧60根;02号草鱼左侧肌间骨为68根,右侧61根;03号草鱼左侧肌间骨为76根,右侧58根;三尾草鱼肌间骨左侧平均为71.3根,右侧为59.7根。以脊柱为界,01号草鱼轴上部肌间骨为85根,轴下部32根,尾鳍基部13根;02号草鱼轴上部肌间骨为77根,轴下部36根,尾鳍基部16根;03号草鱼轴上部肌间骨为79根,轴下部38根,尾鳍基部17根;三尾草鱼肌间骨轴上部平均为80.3根,轴下部35.3根,尾鳍基部15.3根。肌间骨不同形态类型的具体分布情况见表3。实验结果表明:轴上部肌间骨数量最多,且具两端两分叉形、两端多叉形等复杂肌间骨形态;轴下部腹腔前几乎无肌间骨,腹腔后含少量肌间骨;尾鳍基部肌间骨数量最少,左侧含量明显多于右侧,分布最为密集。轴下部和尾鳍基部肌间骨均以“I”形及“Y”形为主,形态单一。实验所用成体草鱼鱼体两侧的肌间骨数目及形态均不完全相同,呈不对称分布,由尾至头肌间骨的结构趋向复杂化。

表3 草鱼肌间骨的分布Tab. 3 Distribution of IB in grass carp

图5 草鱼01肌间骨分布Fig. 5 IB distribution of grass carp 01

图6 草鱼02肌间骨分布Fig. 6 IB distribution of grass carp 02

图7 草鱼03肌间骨分布Fig. 7 IB distribution of grass carp 03
3 讨论
3.1 草鱼肌间骨的数目
本实验采用传统形态学解剖的方法对野生二倍体草鱼肌间骨的数目进行研究分析。结果表明,体长38 cm左右的草鱼肌间骨数目约为131根,其中尾鳍基部约15.3根。这与吕耀平等[10]所报道的草鱼平均肌间骨数目为117根不相符,可能是由于尾鳍基部肌间骨与尾鳍形态相似,导致其被忽略而引起本实验结果与之有所差异。也有研究表明远缘杂交技术可减少鱼类肌间骨数量[3],据此推测实验对象也可能引起肌间骨数目的差异,其实验用鱼可能为杂交品种,肌间骨数量也少于野生品种。其次,同种鱼在不同的生态环境中也可能会导致肌间骨数目的差异。
尾鳍作为运动器官对鱼类的快速游动具有重要影响[20]。有学者认为肌间骨在鱼类大侧肌的运动中起辅助作用,并能在肌肉中传导机械力量[21,22]。本实验中尾鳍基部肌间骨最为密集且明显粗壮,因此推测尾鳍基部肌间骨能够协助鱼体的运动,其形态是由于长期辅助鱼体快速游动所致。
3.2 草鱼肌间骨的形态及分布
肌间骨细而长,多呈针状,影响人们对鱼类的食用。根据蒋燕[23]及黎玲[24]等的研究可知,鲤科鱼类仅有轴上肌肌节间的髓弓小骨和轴下肌肌节间的脉弓小骨,不包含隔膜中的椎体小骨。实验草鱼与之结果相符,仅含有髓弓小骨和脉弓小骨两种类型。
鱼类肌间骨形态多样[6],且与分布位置有一定关系。孟庆闻等通过对鲢肌间骨的研究发现,肌间骨在鱼体中呈两侧成对分布[25],但近年研究结果表明肌间骨并非两侧对称分布,其数目及形态均有一定的差异[26,27],本实验支持后者的研究结论,即肌间骨呈两侧不对称分布。本研究将鱼体划分为轴上部、轴下部以及尾鳍基部三个部分,研究表明草鱼的轴上部肌间骨数量较多且形态复杂多样;轴下部肌间骨数量较少,且形态较简单;尾鳍基部肌间骨形态最为简单粗壮,多为“I”形少量“Y”形。所有肌间骨形态较简单的一端向尾部倾斜,形态较复杂的一端则向头部倾斜,由头部向尾鳍有秩序地分布于各个肌节之间[28],该结果与董在杰[7]和聂春红[29]等对鲤科鱼类的研究表述一致。
3.3 草鱼肌间骨的演化
Patterson等认为最原始的肌间骨无分叉形态[8]。孟庆闻和苏锦祥等通过对鲢鱼肌间骨的研究,将其分为“I”、“卜”、“Y”及树枝形四种类型[25]。董在杰等根据几种常见鲤科鱼类肌间骨的形态将其分为两种基本类型:“I”形和“Y”形,并认为其余几种较复杂形态均是在此基础上演化而来[7]。真骨鱼类肌间骨的外形结构随鱼类由低等到高等而发生变化,且愈加复杂化[28]。通过对几种低等真骨鱼类进行解剖,有学者按照其肌间骨形态复杂性将其细分为更利于剖析各种形态肌间骨演变路径的7种类型(即“I”形,“卜”形,“Y”形,一端多叉形,两端两分叉形,两端多叉形,树枝形)[10]。本文参考上述方法将草鱼肌间骨分类,结果表明草鱼肌间骨分为“I”形、“L”形、“卜”形、“Y”形、一端多叉形、两端两分叉形及两端多叉形7种类型。有研究认为形态越复杂的肌间骨,其所需演化时间越长[3],本实验中两端两分叉、两端多分叉等较复杂肌间骨主要出现在头部,越接近尾部其形态越简单,且以“I”形为主,此结果支持上述观点。
如今,肌间骨的研究备受关注并取得一定进展,但其功能、演化路径暂无定论。对于草鱼肌间骨的演化路径也存在一定争议,有研究认为所有肌间骨均以“I”形为基础转化而来。草鱼肌间骨的演化路径可能为:①“I”形-“卜”形-“Y”形-一端多叉形或②“I”形-“卜”形-“Y”形-两端两分叉形-两端多叉形-树枝形。本实验研究发现在草鱼中同时出现一端多叉、两端两分叉及两端多叉形肌间骨。据此推测草鱼肌间骨的演化路径可能为:“I”形-“L”形-“卜”形-“Y”形-一端多叉形-两端两分叉形-两端多叉形,但关于草鱼肌间骨的具体演化路径还有待进一步的探索。
4 总结
本研究表明:草鱼无椎体小骨,其肌间骨的数量可能受水域环境的影响。进行肌间骨的研究分析时,尾鳍基部的肌间骨易被忽视,但在草鱼中,尾鳍基部肌间骨分布密集,形态最为粗壮,且可能与长期协助鱼体游动相关,有待进一步研究。本文在草鱼中发现“一端多叉形”为草鱼中最新发现的肌间骨类型。研究表明“L”形可能是“I”形演化至“卜”形的一种过渡形态,故推测草鱼的肌间骨演化路径可能为:“I”形-“L”形-“卜”形-“Y”形-一端多叉形-两端两分叉形-两端多叉形。
