珠江浮游真核微型生物分子多样性及其与水环境的关系
2020-03-04朱昶宇陆凯晖伊珍珍
朱昶宇 陆凯晖 伊珍珍
(华南师范大学生命科学学院, 广州市亚热带生物多样性与环境生物监测重点实验室, 广州 510631)
珠江水资源总量丰富, 在全国七大江河中排名第二, 仅次于长江[1]。然而伴随着珠三角地区经济高速发展、城市规模不断扩大, 各种工业和生活污水、有机污染物不断迁移入珠江水域, 珠江下游水污染问题严重[2]。珠江主要支流西江和北江在流经广东省后污染状况同样不容乐观, 西江水质总体较差, 支流贺江水体整体处于中营养水平, 发生藻类水华的潜在风险较高[3]。珠江广州段和西江广东段承载着珠三角地区重要的城市供水任务, 与人民生活质量以及当地经济发展息息相关, 日趋严重的水污染所导致的水质性缺水, 已成为制约城市可持续发展的一个重要因素[4]。此外, 珠江流域处于热带-亚热带地区, 丰水期(4—9月)径流量大, 占全年径流总量的80%, 而枯水期(10月至翌年3月)的水量较少,仅占全年径流总量的24%[5], 这意味着对珠江水质状况的研究有必要考虑丰水期及枯水期之间的差异, 但目前相关研究较少[6,7]。
真核微型生物在水生生态系统中普遍存在, 主要包括原生动物、真核微藻、真菌及线虫等小型动物[8], 由于生命周期短、敏感性强等特点, 能迅速反映水环境变化[9,10]。例如, 浮游藻类群落分布与温度和营养盐变化显著相关[11], 某些原生动物类群丰度与氨氮浓度呈显著正相关[12]。徐润林等[6]对珠江广州市段的原生动物群落组成和群集过程的研究发现, 原生动物群落特征的变化与水质变化相吻合, 珠江在流经广州市区后水质发生了明显地恶化。20世纪90年代, 沈韫芬等[13]便建立了适用于我国淡水生态环境的原生生物(真核微型生物)群落监测方法, 并成为我国第一个淡水生物监测标准。近年来, 徐奎栋等[14]将其改良并应用到海水环境监测中, 取得了较好的效果。
由于个体微小(通常为几十至几百微米), 基于形态鉴定基础的真核微型生物群落研究, 高度依赖于研究人员的鉴定经验, 在一定程度上限制了原生动物多样性研究及其在水质监测中的广泛应用。这也是珠江水体真核微型生物群落结构与环境因子的关系研究较少的重要原因之一[3,6,15]。末端限制性片段长度多态性分析(Terminal restriction fragment length polymorphism analysis, T-RFLP)可以定性和定量的进行微生物群落结构分析及不同群落结构间的比较[16]。目前, T-RFLP被广泛应用于水体微生物多样性及其与水环境关系的研究中, 例如: 环境因素对微生物的群落结构和功能的影响[17],河流生态系统的健康评价[18], 指示生物的筛选[19]。
本文通过T-RFLP研究了珠三角地区珠江丰水期及枯水期浮游真核微型生物多样性, 对比分析了不同时期、不同区域真核微型生物的群落结构及其与环境因子的关系, 并筛选了潜在的敏感种和耐污种。以期较为全面地了解珠三角地区珠江水体中真核微型生物的动态变化, 为后续珠三角地区珠江水质监测提供可靠的数据参考。
1 材料与方法
1.1 水样采集
采样点分为珠江广州段(G)和西江广东段(X),分别在其丰水期(2016年6月—2016年9月)和枯水期(2016年10月—2017年1月)采集水样, 使用有机玻璃采水器采集表层水(30 cm—1 m)。其中, 丰水期设置了40个样点(西江广东段21个, 珠江广州段19个),枯水期设置了43个样点(西江广东段21个, 珠江广州段22个, 图1)。
1.2 水质分析
现场使用便携式多参数测量仪(ORION 520M-01A)测定采样位点的水温(Temperature, Temp)、pH、溶解氧(Dissolved oxygen, DO)和盐度(Salinity)。其余理化指标参照《Water Analysis Handbook》(美国哈希公司)于实验室测定, 总氮(Total nitrogen, TN)的测定采用过硫酸盐氧化法(0.5—25.0 mg/L); 氨氮(Ammonia nitrogen, AN)的测定采用USEPA纳氏试剂法(0.02—2.5 mg/L)或水杨酸法(0.4—50 mg/L); 总磷(Total phosphorus, TP)的测定采用USEPA PhosVer 3抗坏血酸法(0.06—3.5 mg/L);化学需氧量(Chemical oxygen demand, CODCr)的测定采用USEPA消解比色法(3—150 mg/L); 高锰酸盐指数(Permanganate index, CODMn)的测定采用高锰酸盐法(0.5—5.0 mg/L)。
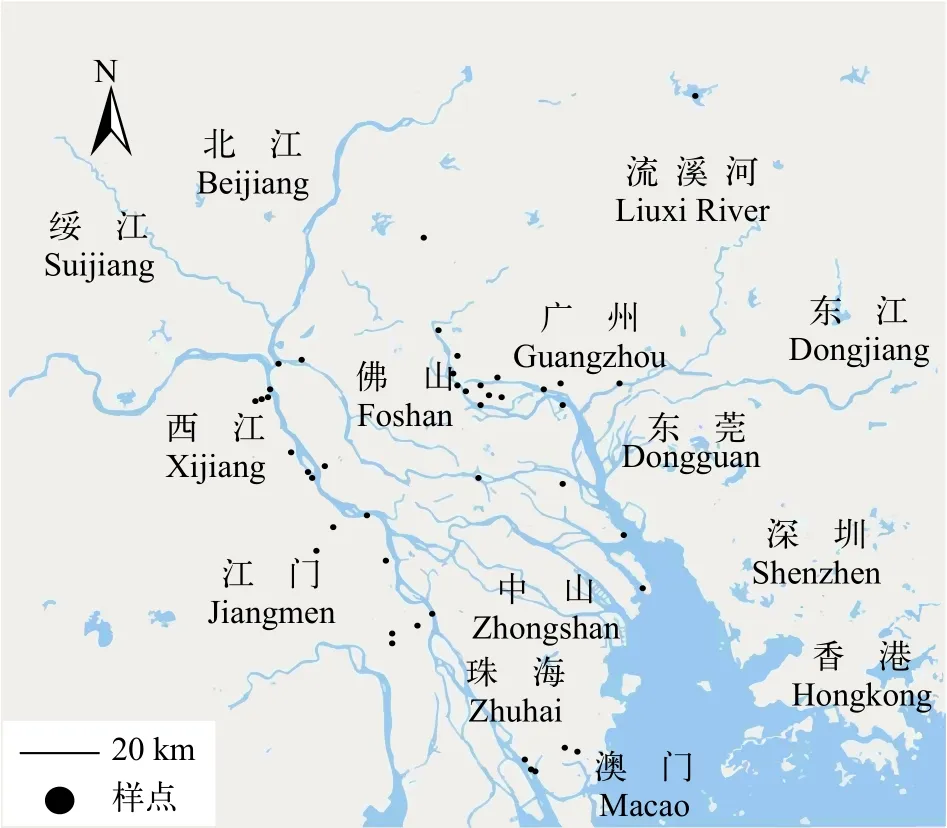
图1 采样点分布Fig. 1 Locations of sampling sites
1.3 生物富集与DNA提取
将200 mL水体抽滤到0.8 μm滤膜(美国颇尔公司)上, 用灭菌镊子将滤膜取下, 在超净工作台上用灭菌剪刀将滤膜剪碎, 使用PowerSoil DNA Isolation Kit (美国Mobio)进行环境总DNA提取。
1.4 末端限制性片段长度多态性分析(T-RFLP)
利用引物TAReuk454FWD1 (5′-CCAGCA(G/C)C(C/T)GCGGTAATTCC-3′), TAReukREV3(5′-ACTTTCGTTCTTGAT(C/T)(A/G)A-3′)[20]扩增真核微型生物18S rDNA的V4区。PCR体系为: 5 ×FastPfu Buffer, 2.5 mmol/L dNTPs, 1U FastPfu DNA聚合酶(北京全式金生物技术有限公司), 引物各5 μmol/L, 约10 ng基因组DNA, 反应体系总量为20 μL。PCR反应程序: 95℃预变性5min; 95℃变性30s, 55℃退火30s, 72℃延伸45s, 共27个循环; 最后72℃延伸10min, 终止于4℃。PCR产物用1%琼脂糖电泳检验扩增效果, 并采用超薄DNA产物纯化试剂盒DP203-02(北京天根生化科技有限公司)纯化。
预实验选用了ApaⅠ、BamH Ⅰ、EcoRⅠ、Hind Ⅲ、HinfⅠ、HaeⅢ等限制性核酸内切酶进行处理, 根据酶切效率、区分能力等因素, 最终选取HaeⅢ构建酶切体系: 10 μL回收产物, 2 μL QC缓冲液, 0.4 μLHaeⅢ酶(日本TaKaRa公司)和7.6 μL灭菌双蒸水。放置水浴锅中37℃温浴1h,1%琼脂糖电泳检验酶切效果。最后将剩余酶切产物进行毛细管电泳并检测荧光和荧光强度(上海生工生物工程有限公司)。
1.5 数据分析
数据取整, 保留50—500 bp内的末端限制性片段数据(T-RFs), 看作是物种种类数。计算同一样品的所有峰度面积之和, 以及各片段面积占所在样品总面积的比例, 看作是物种丰度[21]。
使用R语言(R-3.2.3)程序包Vegan[22]计算多样性指数, 基于Bray-Curtis距离[23]量化样本间群落结构差异, 使用Adonis进行组间差异显著性检验[24],并进行典范对应分析[25]、蒙特卡罗检验(Monte Carlo permutation test)[26]。多样性指数包括物种数(S), Shannon-Wiener指数(H′)[27]和Pielou指数(J′)[28]。典范对应分析反映物种或样点与环境因子间的关系。蒙特卡罗检验用于检验群落结构与理化因子相关性是否显著(重复数为999)。
参考地表水环境质量标准《GB 3838—2002》,综合丰水期及枯水期数据, 按水质类别将水质状况分为优(Ⅰ、Ⅱ类水质)、良(Ⅲ类水)、轻度污染(Ⅳ类水质)、中度污染(Ⅴ类水质)及重度污染(劣Ⅴ类水质)。使用R语言indicspecies程序包[29]进行组间指示种分析, 分别筛选了代表优(Ⅰ、Ⅱ类水)及重度污染(劣Ⅴ类水)潜在的敏感种和耐污种的末端限制性片段。并使用MICA工具进行末端限制性片段对应物种预测[30]。使用R语言vegan[22]程序包中的corr.test功能对物种(T-RFs)和理化因子进行Spearman秩相关分析, 并使用corr.test检验其相关性是否显著(重复数为999)。
2 结果
2.1 真核微型生物多样性
丰水期40个样本中共发现133个不同的末端限制性片段, 代表了至少133个不同的物种。样点末端限制性片段数目最少为2个(GG2_1), 最多为25个(XG5_1); 香农威尔指数最小也为0.6734(GG2_1),最大为3.0611 (XG5_1); 均匀度指数最小为0.5564(XS5_1), 最大为0.9715 (GG2_1, 表1)。由此可见,珠江广州段样本GG2_1物种组成单一, 且末端限制性片段分布不均匀; 而西江广东段样本XG5_1物种组成较为多样化, 且末端限制性片段分布较为均匀。
枯水期43个样本中共发现167个不同的末端限制性片段, 末端限制性片段数目最少为10个(GC1_2、GG3_2、GN2_2、XN2_2), 最多为29个(XG1_2和XS1_2)。珠江广州段样本GN2_2的均匀度指数最小(0.5375), 香农威尔指数也最小(1.2377),说明该样本物种组成单一, 且T-RFs分布不均匀。西江广东段样本XG1_2的均匀度指数最大(0.9493),香农威尔指数也最大(3.1966), 其末端限制性片段数目虽多(29), 但各个物种的数量相差不大, 物种组成较为多样化。
丰水期样本与枯水期样本的末端限制性片段数目与均匀度指数无显著差异, 但是香农威尔指数存在显著差异(P<0.05, 图2a), 枯水期样本多样性指数普遍高于丰水期样本(表1)。珠江广州段样本与西江广东段样本多样性指数均存在显著差异(P<0.05,图2b), 西江广东段样本多样性指数普遍高于珠江广州段样本(表1)。
真核微型生物非度量多维尺度分析(图3)显示,丰水期40个样本(WS)较枯水期43个样本(DS)的分布更为分散, 珠江广州段41个样本(GZ)较西江广东段42个样本(XJ)的分布更为分散。Adonis分析表明丰水期与枯水期样本之间、珠江广州段与西江广东段样本之间的真核微型生物群落结构均存在显著差异(P<0.05, 图3)。
2.2 真核微型生物群落结构及其与理化因子的关系
调查区域83个样本中, 水质类别为优、良、轻度污染、中度污染和重度污染的样本分别为3、7、9、7和57个。57个样本的总氮(TN)、氨氮(AN)和总磷(TP)指标均超过地表水劣Ⅴ类水标准,为珠江水体主要污染指标。枯水期样本温度、pH、盐度和总磷显著高于丰水期样本(P<0.05, 图4a),其余所测理化因子在枯水期和丰水期无显著差异。西江广东段样本化学需氧量显著高于珠江广州段样本(P<0.05, 图4b), 其余所测理化因子在西江广东段和珠江广州段均无显著差异。
丰水期样本典范对应分析(图5a)和相关性检验(表2)表明, 丰水期样本真核微型生物群落结构与溶氧量(DO)、化学需氧量(CODCr)、高锰酸盐指数(CODMn)、氨氮(AN)、总氮(TN)和总磷(TP)显著相关(P<0.05), 其中对群落结构影响最大的理化因子是总磷(TP), 相关系数(r2)为0.7885, 其次是氨氮(AN), 相关系数为0.6976。其余理化因子相关系数由大到小分别是高锰酸盐指数(CODMn, 0.6797)、化学需氧量(CODCr, 0.6170)、溶氧量(DO,0.5599)、总氮(TN, 0.5397)。枯水期样本典范对应分析(图5b)和相关性检验(表2)表明, 枯水期样本真核微型生物群落结构受温度(Temp)、化学需氧量(CODCr)、高锰酸盐指数(CODMn)、氨氮(AN)、总氮(TN)和总磷(TP)的显著影响, 其中受总磷(TP)和高锰酸盐指数(CODMn)影响最大, 相关系数分别是0.9289和0.7950, 其次是化学需氧量(CODCr,0.6815)、总氮(TN, 0.6638)、氨氮(AN, 0.6352)、溶氧量(DO, 0.4449)和温度(TP, 0.1695)。与丰水期相比, 枯水期样本真核微生物群落结构与造成污染的理化因子如总磷(TP)、总氮(TN)和化学需氧量(CODCr)等的相关系数要大于丰水期, 且温度(Temp)对枯水期样本真核微型生物群落结构有显著影响。
珠江广州段样本典范对应分析(图5c)和相关性检验(表2)表明, 珠江广州段样本真核微型生物群落结构与温度(Temp)、溶氧量(DO)、化学需氧量(CODCr)、高锰酸盐指数(CODMn)、氨氮(AN)、
总氮(TN)和总磷(TP)显著相关(P<0.05), 其中对群落结构影响最大的理化因子是高锰酸盐指数(CODMn), 相关系数(r2)为0.8313, 其次是化学需氧量(CODCr), 相关系数为0.7687。其余理化因子相关系数由大到小分别是氨氮(AN, 0.6086)、总氮(TN,0.4973)、总磷(TP, 0.1925)、温度(Temp, 0.1329)、溶氧量(DO, 0.1294)。西江广东段样本典范对应分析(图5d)和相关性检验(表2)表明, 西江广东段样本真核微型生物群落结构与溶氧量(DO)、化学需氧量(CODCr)、高锰酸盐指数(CODMn)、氨氮(AN)、总氮(TN)和总磷(TP)显著相关(P<0.05), 其中对群落结构影响最大的理化因子是总磷(TP), 相关系数(r2)为0.9285, 其次是高锰酸盐指数(CODMn),相关系数为0.8722。其余理化因子相关系数由大到小分别是氨氮(AN, 0.8681)、化学需氧量(CODCr,0.8200)、总氮(TN, 0.8047)、溶氧量(DO, 0.2305)。与珠江广州段样本相比, 西江广东段样本真核微型生物群落结构与理化因子相关系数普遍更大, 但是
温度对西江广东段样本真核微型生物群落结构无显著影响(P>0.05)。
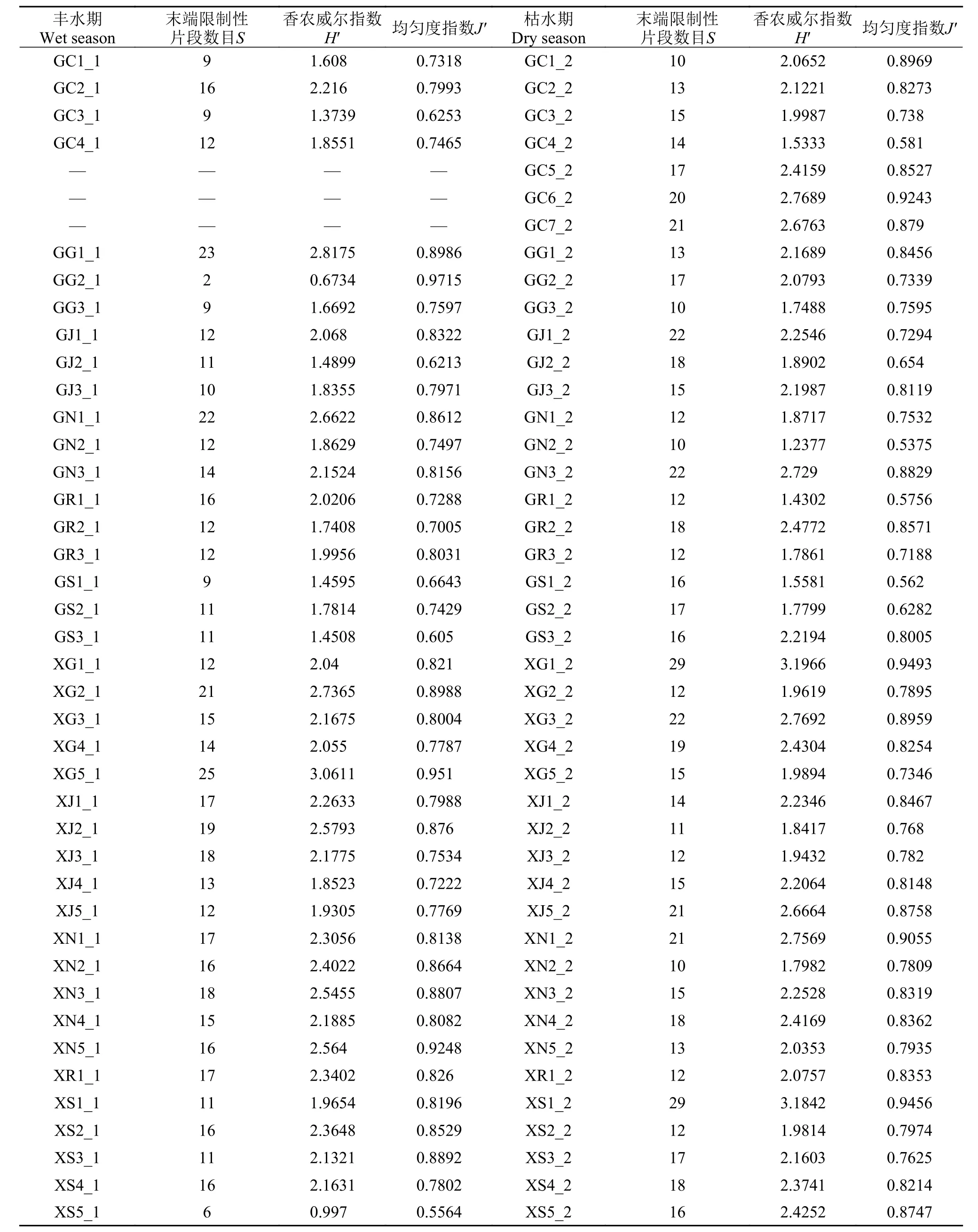
表1 真核微型生物多样性指数Tab. 1 Diversity index of microbial eukaryotes

图2 不同水期(a)和采样区域(b)多样性指数箱线图Fig. 2 The boxplots for diversity index of different water seasons or regions
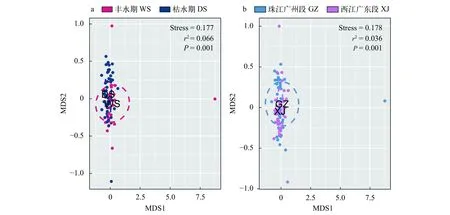
图3 不同水期(a)和不同区域(b)真核微型生物非度量多维尺度分析Fig. 3 The nMDS for microbial eukaryotes of different water seasons (a) and regions (b)
2.3 潜在指示物种筛选
为筛选潜在的对应水质指示物种, 参考地表水环境质量标准《GB 3838—2002》, 在水质类为优(I、II类水)、良(III类水)、轻度污染(IV类水质)、中度污染(V类水质)、重度污染(劣V类水)的样本组间进行组间指示末端限制性片段筛选(表3)。筛选出1个末端限制性片段, 片段长度为76 bp, 可能只存在于“优”级水质, 而在水质为其余类别的水体中基本不存在或数量非常少。筛选出了8个末端限制性片段, 只存在或大量存在于“重度污染”水体中,片段长度分别为83、117、192、235、376、377、391和406 bp。基于数据库比对, 潜在的敏感物种可能为后鞭类(Opisthokonta)的Cystobasidiumsp. 或变形类(Amoebozoa)的Protostelium nocturnum; 潜在的耐污物种可能为后鞭类(Opisthokonta)的Candida caryicola、梅奇酵母科(Metschnikowiaceae)或酿酒酵母目的未定科(Saccharomycetales incertae sedis)物种; 原始色素体生物(Archaeplastida)的Coccomyxa simplex、Botryosphaerella sudetica以及轮藻科(Characeae)、衣藻科(Chlamydomonadaceae)物种;变形类(Amoebozoa)的Acanthamoeba hatchetti; 囊泡类(Alveolata)的Sulfonecta uniserialis、Stenophora robusta、Theileriasp.、Babesia bicornis以及钟形虫科(Vorticellidae)和隐孢科(Cryptosporidiidae)物种; 隐藻类(Cryptophyceae)的Cryptomonas ovata; 不等鞭毛类(Stramenopiles)的Filos agilis和Blastocystissp.。
3 讨论
本研究T-RFLP数据表明, 丰水期与枯水期水体中的真核微型生物香农威尔指数和群落结构存在显著差异(图2a、图3a)。前人研究也表明季节是影响真核微型生物群落组成的重要因素, 例如:Countway等[31]报道同一样点不同季节的真核微型生物物种组成有显著差异; 北太平洋东部海域的真核微型生物群落存在显著的季节和年度差异[32]。与此同时, 本研究发现珠江广州段和西江广东段真核微型生物多样性指数、群落结构也存在显著差异(图2b、图3b), 表明地域甚至是小范围的地理位置差异对真核微型生物多样性有着显著的影响。同样, 先前文献报道, 青岛、厦门和湛江沿岸滩涂真核微型生物群落结构存在显著差异[33]; 美国黄石公园不同样点间真核微型生物群落相似性较低[34]。考虑到时间和地域的差异对物种多样性有显著的影响, 我们建议在利用真核微型生物监测水质时,必须建立在全面了解该地区不同时期、不同地点的真核微型生物多样性的基础上。
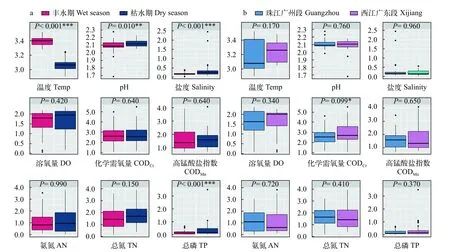
图4 不同水期(a)和不同区域(b)的理化因子箱线图。Fig. 4 The boxplots for physicochemical factors of different water seasons (a) and regions (b)
我们的研究数据表明, 珠江广州段和西江广东段水体受到氮、磷的严重污染, 普遍处于劣V类(57个样本, 占据了69%); 丰水期总磷浓度普遍低于枯水期, 西江广东段化学需氧量浓度普遍高于珠江广州段(图4)。排序分析和相关性检验(图5、表2)表明, 真核微型生物群落结构与化学需氧量、高锰酸盐指数、氨氮、总氮、总磷等水质监测相关理化因子显著相关, 能够较好地反映水质状况。化学需氧量和高锰酸盐指数代表着样点受有机物污染的程度[35], 其对真核微型生物群落结构有着显著的影响[36,37]。过量的氨氮会影响细胞膜稳定性及酶水解反应, 导致生物代谢缓慢, 严重时可导致生物死亡[38], 文献报道原生生物、金藻、硅藻与氨氮之间呈现显著正相关[12]。总氮和总磷通过影响浮游藻类的光合作用对其生长进行调控, 从而又影响了异养真核微型生物类群[39—43]。总氮和总磷是导致水体富营养化的主要理化因子[44], 此类营养盐主要来源于工业废水和生活污水, 而西江广东段与珠江广州段周边工业废水和生活污水排放量巨大[15]。此外, 本研究发现, 研究区域总氮和总磷的季节性变化不明显, 但枯水期高于丰水期, 这可能是由于丰水期水量大, 物理稀释作用导致总氮和总磷浓度降低, 也可能是因为丰水期水生生物活动频繁, 大量消耗了水体中的营养盐[35]。
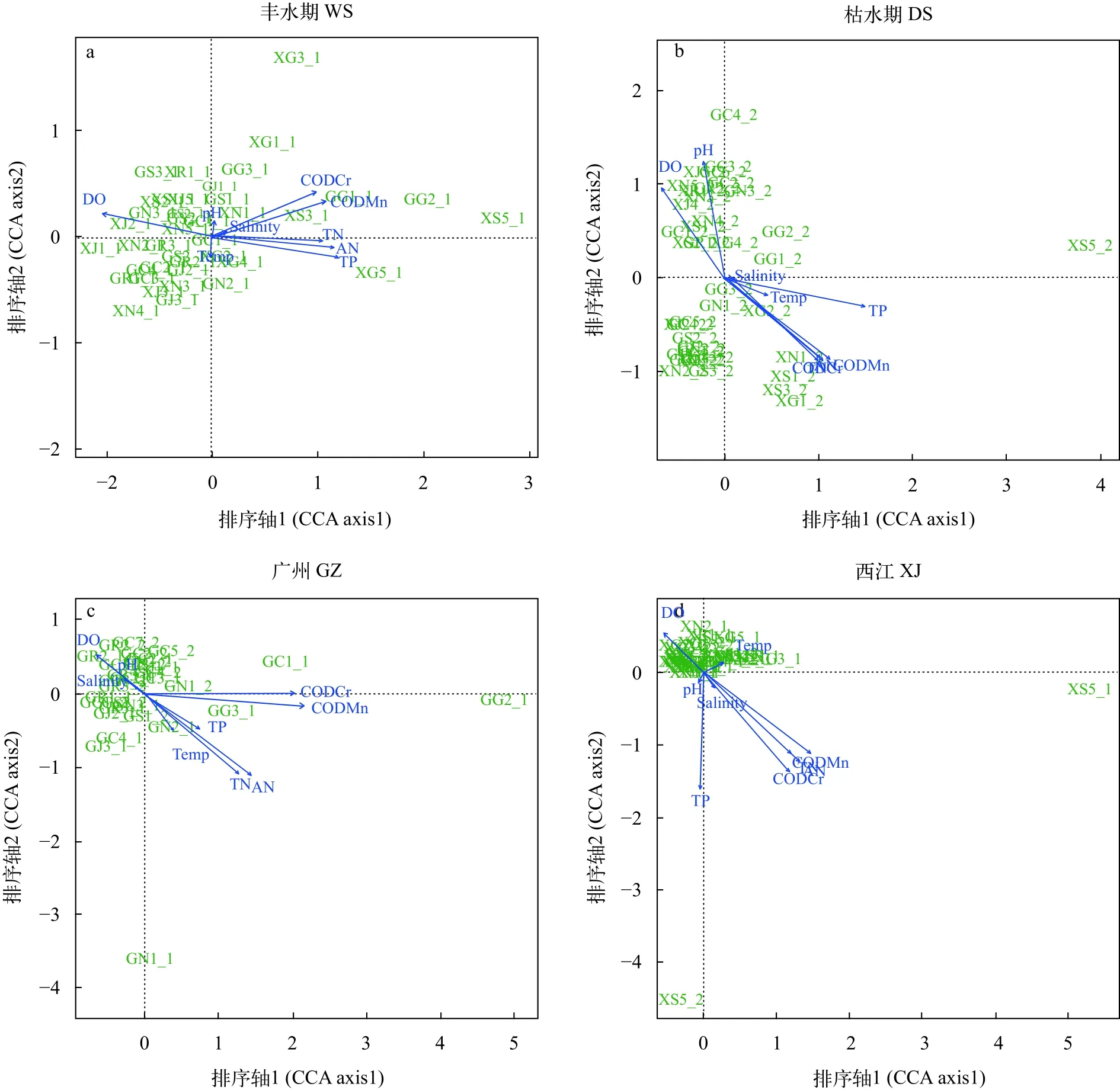
图5 丰水期样本(a)、枯水期样本(b)、珠江广州段(c)和西江广东段(d)样本真核微型群落结构与理化因子典范对应分析Fig. 5 Canonical correspondence analysis (CCA) between physicochemical factors and microbial eukaryotic community structures of wet season (a), dry season (b), Guangzhou Reach of the Pearl River (c) and Guangdong Reach of the Xijiang River (d)
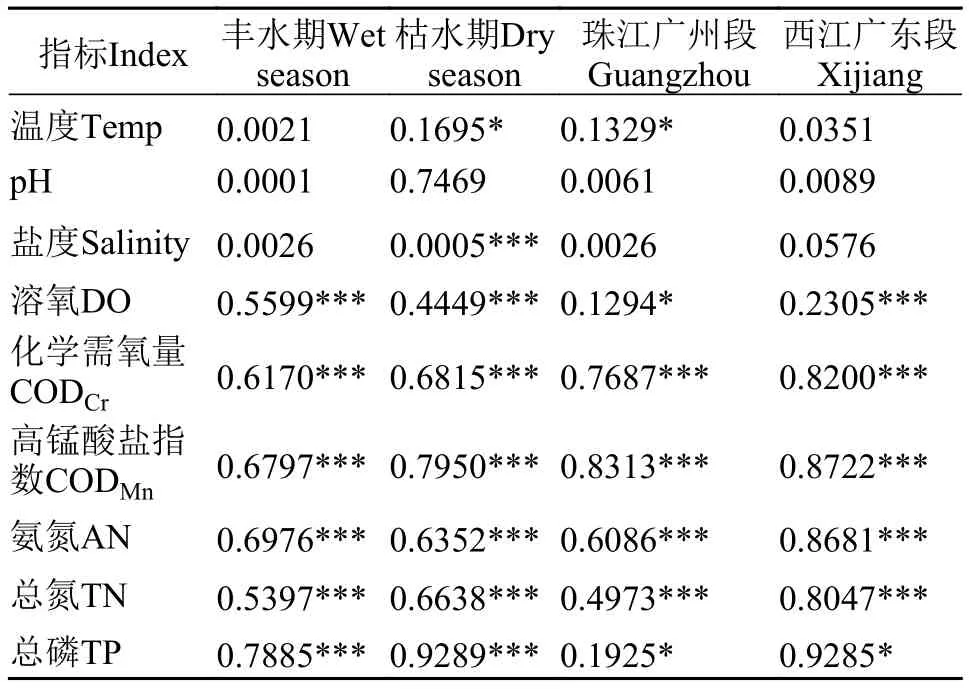
表2 真核微型生物群落结构与理化因子相关性检验Tab. 2 Correlation test between the microbial eukaryotic community structures of samples and physicochemical factors

表3 潜在的水质指示片段Tab. 3 Potential indicative T-RFs
某些真核微型生物, 会出现或大量存在于在不同污染程度的水域中, 从而可以作为水质评判的指示物种。许恒龙等[12]发现Peridinium(多甲藻虫属)的某些物种在较为清洁的水体中丰度较高, 可作为潜在的敏感类群。杨金鹏等[45]发现Uronemella(尾丝虫)在富营养水体中有较高的丰度, 可作为潜在的耐污类群。本研究分别筛选出1个、8个末端限制性片段作为候选的敏感和耐污片段, 并根据数据库给出了大致的物种/类群信息, 但由于缺乏具体的碱基信息, 这些片段具体代表哪些物种还有待进一步研究。
致谢:
感谢华南师范大学本科生张宝丹、陈静慧、陈俊宇在部分样品采集和实验操作中给予的帮助。
