西藏温泉两种中国新记录纤毛虫第一双小核草履虫和明布雷斯四膜虫的形态学和系统发育学研究
2020-03-04姜传奇谷思雨安瑞志
姜传奇 谷思雨 安瑞志 巴 桑* 缪 炜*
(1. 中国科学院水生生物研究所水生生物多样性与保护重点实验室, 武汉 430072; 2. 哈尔滨师范大学黑龙江省水生生物多样性研究重点实验室, 哈尔滨 150025; 3. 西藏大学理学院湿地与流域生态实验室, 拉萨 850000)
纤毛虫隶属原生生物界, 原生动物亚界, 是较复杂的单细胞真核生物。纤毛虫物种丰富, 已记载8000余种, 广泛分布于淡水、海水、土壤甚至一些极端环境中[1]。青藏高原是“世界屋脊”, 有着独特的自然环境和丰富的生物资源。其中, 西藏地区地热资源非常丰富, 各种地热显示遍布全区。1966年和1973—1976年, 王家楫等在第一次青藏高原综合科学考察中采集了大量的无脊椎动物标本, 随后完成的《西藏高原部分地区的原生动物》、《珠穆朗玛峰地区的原生动物》、《西藏水生无脊椎动物》共记述了西藏地区原生动物458种, 其中纤毛虫288种[2—4]。期间, 也对部分温泉的原生动物进行调查, 共鉴定到原生动物52种, 其中纤毛虫有13种[4]。近年来, 随着纤毛虫培养技术和研究手段的逐步发展完善, 可以对西藏温泉生境该类群的生物资源开展更深入和系统的研究。
本研究对位于西藏地区的日多温泉和羊八井温泉进行样品采集, 鉴定得到了两种寡膜纲纤毛虫中国新记录种: 咽膜类的第一双小核草履虫(Para-mecium primaurelia)和膜口类的明布雷斯四膜虫(Tetrahymena mimbres)。本研究详细描述了这两种纤毛虫的形态学特征, 测定了它们的SSU rDNA和COXⅠ基因序列并基于这两种分子标记进行了系统发育学分析。
1 材料与方法
1.1 样品采集与处理
两种纤毛虫第一双小核草履虫和明布雷斯四膜虫于2018年10月采自西藏高原, 其中P. primaurelia采自拉萨市墨竹工卡县的日多温泉(N 29°41′,E 92°14′; 海拔: 4365 m; 水温: 50—60℃),T. mimbres采自当雄县的羊八井温泉(N 29°58′, E 90°21′; 海拔:4350 m; 水温: 43℃)。使用采样瓶收集温泉原位水及悬浮物。样品采集后, 在实验室将不同采集地点的水样摇匀分别倒入培养皿中, 每个培养皿加入两粒已灭菌的麦粒, 将培养皿盖好盖子分别放入封闭样品盒内以避免样品交叉污染。培养4—5d后, 在解剖镜(ZEISS Stemi2000-C)下, 利用微吸管分离单克隆株系, 并使用过滤(滤膜规格: 0.22 μm)原位水加入灭菌麦粒于25℃扩大培养。
1.2 形态学研究
在解剖镜下利用微吸管挑取少量纤毛虫于载玻片上进行活体压片, 在显微镜下(100×—1000×)观察虫体的大小、颜色、形状、运动方式和伸缩泡等活体特征。采用蛋白银染色法[5]和氨银染色法[6]制片, 分别获得P. primaurelia和T. mimbres的纤毛图式和核器特征等。此外, 细胞核的形态还以2%多聚甲醛(PFA)固定细胞, 经0.5 μg/mL DAPI(4′, 6-二脒基-2-苯基吲哚)核酸染料染色,在荧光显微镜下观察。显微观察及拍照利用尼康显微镜(Nikon EC-LIPSE 80i)。系统分类和术语依据Lynn(2008)。
1.3 DNA提取和PCR扩增
第一双小核草履虫(P. primaurelia)和明布雷斯四膜虫(T. mimbres)的DNA提取利用REDExtract-NAmpTMPCR试剂盒, 利用微吸管分别挑取少量的T. mimbres和P. primaurelia细胞, 经蒸馏水清洗数次后, 放入PCR管中, 按照试剂盒说明书操作, 释放总DNA(www.sigmaaldrich.com)。两种纤毛虫SSU rDNA序列的扩增利用真核生物通用引物18S-F: 5′-AACCTGGTTGATCCTGCCAGT-3′和18S-R: 5′-TGATCCTTCTGCAGGTTCACCTAC-3′[7];P.primaurelia COXⅠ基因的扩增利用引物F199dT-B:5′-TGTAAAACGACGGCCAGTTCAGGWGCT GCMTTAGCHACYATG-3′和R1143dT: 5′-CAG GAAACAGCTATGACTARTATAGGATCMCCW CCATAAGC-3′;T. mimbresCOXⅠ基因的扩增利用引物F298dT: 5′-TGTAAAACGACGGCCAGT GCNCAYGGTYTAATNATGGT-3′和R1184dT: 5′-CAGGAAACAGCTATGACTADACYTCAGG GTGACCRAAAAATCA-3′[8]。PCR后, 扩增产物直接送测序公司测序。
1.4 系统发育学分析
除新测序列外, 其余序列均来自GenBank数据库, 包括109条SSU rDNA基因序列和112条COXⅠ基因序列(序列信息详见图版Ⅰ、图版Ⅱ)。选择七种盾纤类纤毛虫(Pseudocohnilembus persalinus、Pseudocohnilembus longisetus、Philasterides dicentrarchi、Miamiensis avidus、Cyclidium glaucoma、Entodiscus borealis、Plagiopyliella pacifica)作为外群构建系统发育树。序列比对利用MEGA 6.0中的Muscle程序[9], 比对完成后, 两端截平并手动删除无法明确比对的模糊区域。最大似然树(ML)的构建采用MEGA 6.0软件, 选择GTR+I+G最优模型, 节点置信值来自1000叠代重复抽样[9]。贝叶斯树(BI)利用MrBays 3.2.1软件构建, 选择GTR+I+G最优模型[10], MCMC模拟运行10000000代, 每100代取样1次, 前10000棵树舍弃, 剩余的树用来计算后验概率。使用MEGA 6.0软件调整树形[9]。
2 结果
2.1 第一双小核草履虫的形态学描述(图版 Ⅰ, 表1)
活体约(150—200) μm × (60—100) μm, 腹面观为长椭圆形或足形, 前端略尖, 具有明显的斜截区,后端口区以下逐渐变窄, 虫体不对称(图版 Ⅰ-A、E—I)。口区位于虫体腹面中部(图版 Ⅰ-A、F、G、N)。虫体浅灰色, 体内充满内含物, 包括食物颗粒、晶体和食物泡等(图版 Ⅰ-A、E—I)。大核一个, 椭圆形, 位于虫体中部, 约40 μm; 小核两枚,靠近大核, 约2 μm (图版 Ⅰ-A、R)。伸缩泡两个,分别于虫体前端和后端(图版 Ⅰ-A、H)。体纤毛整齐, 长约6 μm; 约十根尾纤毛聚集呈簇, 位于虫体尾部, 长约15 μm (图版 Ⅰ-I)。运动方式为沿身体长轴旋转前进, 不时在水中快速自由穿梭。
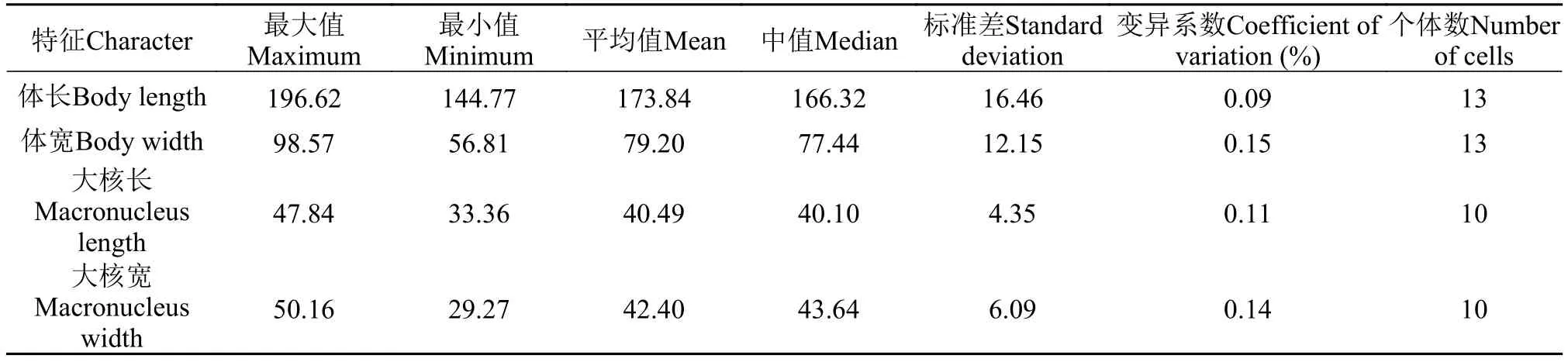
表1 第一双小核草履虫的形态学数据Tab. 1 Morphometric data of Paramecium primaurelia
体动基列60—80列(图版 Ⅰ-B, C, J, K), 口前和口后缝合线明显, 口前缝合线从虫体顶端延至体中部胞口处, 口后缝合线从口区底端延伸至尾部(图版 Ⅰ-B、K—M)。口器具有属典型特征: 由三片小膜(P1、P2、P3)和一片口侧膜(PM)组成。三片小膜均由四列毛基列组成, P1四列毛基粒分散, P1长于P2, P3最短(图版 Ⅰ-D、O—Q)。
2.2 明布雷斯四膜虫(Tetrahymena mimbres)的形态学描述(图版 Ⅱ, 表2)
活体约(35—50) μm × (15—25) μm, 呈卵圆形,前端稍尖, 后端钝圆(图版 Ⅱ-A、E—H), 长宽比约2﹕1。口区较小, 位于体前端1/4处, 约占体长的1/5—1/6。口纤毛长6—7 μm; 体纤毛密集排列, 长4—5 μm (图版 Ⅱ-A、F—H)。细胞质无色至灰白色, 含有数个随机分布的充满细菌的食物泡(直径约3 μm, 图版 Ⅱ-A、E—H)。大核一个, 球形或卵圆形, 直径约10 μm, 位于体中部, 无小核(图版 Ⅱ-I)。伸缩泡一个, 位于体后部, 直径约6 μm, 伸缩周期约1min (图版Ⅱ-A、H)。
体动基列16—18列, 纵向排列, 靠近虫体顶部毛基粒密集, 虫体后部较稀疏(图版 Ⅱ-B、C、J—N)。口区前有明显缝合线, 由口区前端延伸至虫体顶端无毛区(图版 Ⅱ-B、J、M)。口后动基列(PK)两列, 口后动基列1 (PK1)开始于口侧膜中间位置, 向后延伸至细胞末端; 口后动基列2(PK2)开始于口侧膜底部, 延伸至虫体末端(图版 Ⅱ-B、J)。口腔纵轴与体轴平行, 口区内为典型的“四膜”式构造, 包括三片口区小膜(M1、M2、M3)及一片口侧膜(PM)。M1略长于M2, M1与M2近乎平行, 均由三列平行的毛基粒构成; M3较短, 形状近似圆形。口侧膜起始于M2前端, 围绕三片小膜边缘至口区底部(图版 Ⅱ-D、O)。
2.3 系统发育学分析
本研究两种纤毛虫的SSU rDNA和COXⅠ基因序列已提交至GenBank数据库, 其中P. primaurelia的SSU rDNA序列(序列号: MK579298, 长度1649个碱基, GC含量43%)与GenBank中该种的序列(MG573196和MG573195)的相似度为100%、99%;其COXⅠ基因序列(序列号: MK715448, 长度859个碱基, GC含量41%)与GenBank中该种的序列(FN421324和FJ905141)相似度均为99%。T.mimbres的SSU rDNA序列(序列号: MK579316, 长度1655个碱基, GC含量43%)与GenBank中该种(EF070251)的相似度为99%; 其COXⅠ基因序列(序列号: MK715449, 长度876个碱基, GC含量27%)与GenBank该种的序列(EF070292)的相似度为99%。
本研究选择GenBank数据库中有SSU rDNA序列信息的38种四膜虫(46条序列)、20种草履虫(56条序列), 以寡膜纲七种盾纤类纤毛虫作为外类群构建了系统发育树(图1)。结果显示: (1)草履虫科单系。(2)新测序的西藏温泉种P. primaurelia与来自GenBank数据库中的四个P. primaurelia种群(MG573194、MG573195、MG573196和AF100 315)聚为一枝, 聚在Paramecium aurelia复合种(P.aureliacomplex)内。(3)四膜虫科单系, 包括两个主要类群:T. borealis类群(T. borealisgroup)和T. australis类群(T. australisgroup); (4)新测序的西藏温泉种T. mimbres与GenBank的T. mimbres(EF070251)聚为一枝, 与T. elliotti(EF070246)聚为姐妹枝, 聚在T. borealis类群中。
COXⅠ基因系统发育树利用GenBank数据库中四膜虫的57条和草履虫的48条序列, 以盾纤类纤毛虫为外类群构建(图2)。结果显示: (1)本研究新测序的P. primaurelia与GenBank中11种P. primaurelia种群均聚为一枝, 聚在P. aurelia复合种(P. aureliacomplex)中。(2)本次新测序的T. mimbres与Gen-Bank中T. mimbres(EF070292)聚为一枝, 与T.elliotti(EF079281)聚为姐妹枝, 共同聚在T. borealis类群中。
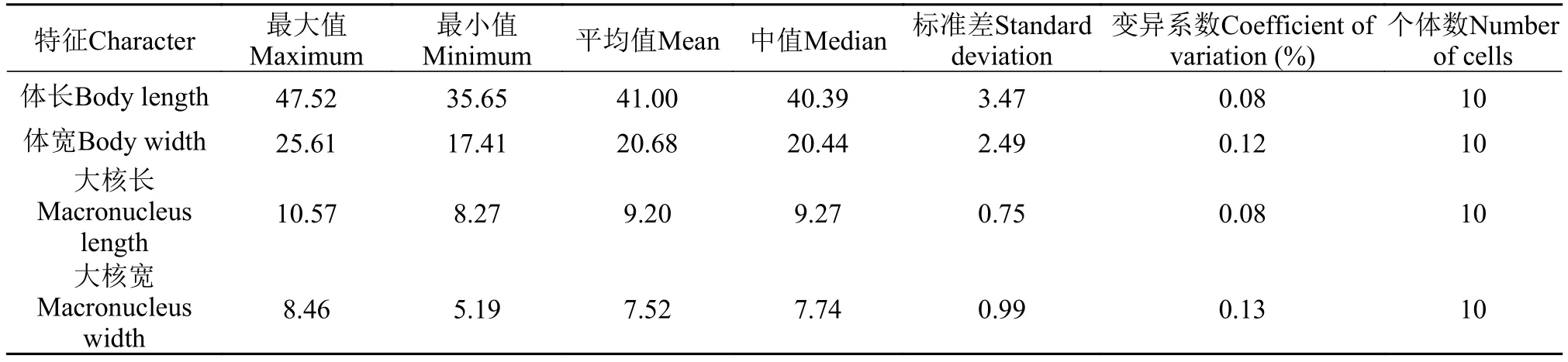
表2 明布雷斯四膜虫的形态学数据Tab. 2 Morphometric data of Tetrahymena mimbres
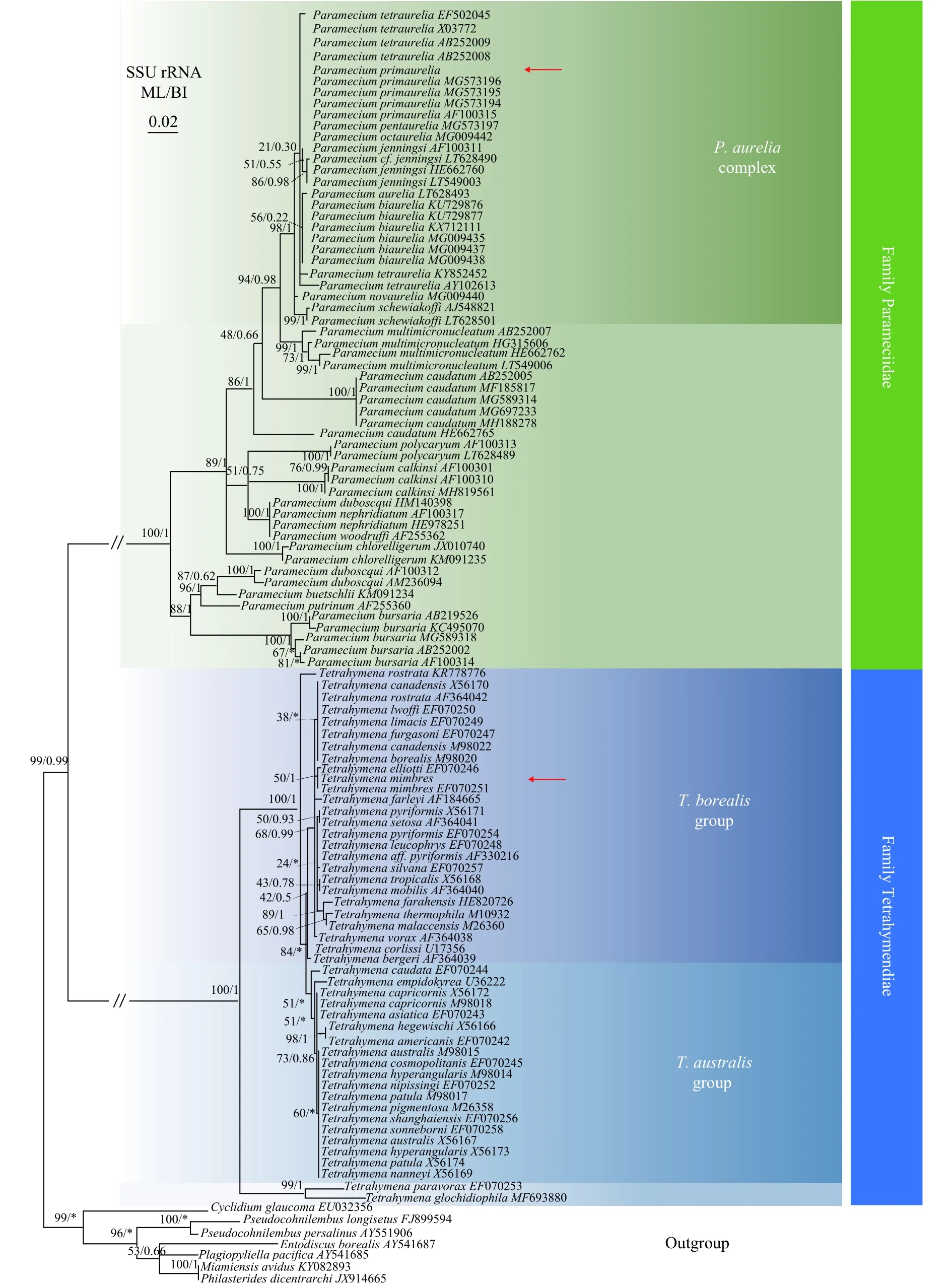
图1 基于核糖体小亚基基因序列构建的最大似然树Fig. 1 Maximum likelihood (ML) tree based on small subunit ribosomal DNA sequences

图2 基于线粒体细胞色素氧化酶Ⅰ基因序列构建的最大似然树Fig. 2 Maximum likelihood (ML) tree based on cytochrome oxidase subunit Ⅰgene sequences
3 讨论
第一双小核草履虫(Paramecium primaurelia)由Sonneborn于1975年命名, 属于P. aurelia复合种[11]。Gill和Hanson(1968)、Hufnagel(1969)和Sonneborn(1975)对P. aurelia复合种进行了形态学研究,P.primaurelia西藏种群符合P. aurelia复合种的形态学特征(虫体雪茄状, 体长80—200 μm, 是体宽的3—4倍; 伸缩泡两枚; 大核一枚、小核两枚; 体动基列60—80列、口区结构、前后缝合线等)[11—13]。本研究利用活体观察、蛋白银、DAPI染色法, 补充了该种活体特征、细胞核的形态和位置、纤毛图式和口区三片咽膜及口侧膜的排布模式, 并提供了该种西藏温泉种群的SSU rDNA基因和COXⅠ基因序列信息。这两种分子标记基因的系统发育学分析均支持P. primaurelia与GenBank中已发表的P.primaurelia种群聚为一枝, 聚在P. aurelia复合种中,与形态学分类结果一致。
明布雷斯四膜虫(Tetrahymena mimbres)最早将其认定为Tetrahymena pyriformis复合种中的NT-1株系[14], 于1987年由Meyer和Nanney将其更名为T.mimbres[15]。目前, 关于该种的研究较少, 无形态学方面的研究。本研究对该种活体形态、细胞核的形态和位置、体纤毛图式和三片口区小膜及口侧膜的排布模式进行了详细的描述, 并提供了该种西藏温泉种群的SSU rDNA和COXⅠ基因序列信息,系统发育学分析支持该种聚在T. borealis种群内部,与T. elliotti的亲缘关系较近。
利用采样瓶收集西藏温泉原位水及悬浮物, 置于实验室中封闭培养, 并在解剖镜下利用微吸管分离单克隆株系, 进一步培养获得了两种中国新记录纤毛虫。其中,P. primaurelia为广布种, 在美洲的南部、北部、中部, 欧洲的俄罗斯、马尔他、保加利亚, 非洲的纳塔尔, 亚洲的日本、缅甸等地均有报道[16—20], 显示出其较强的环境适应性; 首次在西藏日多温泉中的发现, 表明该种还具有较强的高原和高温适应性。T. mimbres的首次发现是在美国新墨西哥州的温泉(Mimbres hot springs)[21], 之后在美国德克萨斯州的温泉中也有报道[22], 表明该种为喜高温种类, 具有明显的高温适应性。T. mimbres首次在西藏羊八井温泉中的发现, 也体现出该种有较强的高原适应性。本研究借助纤毛虫培养技术和形态学、分子生物学研究手段, 鉴定出西藏温泉的两种中国新记录纤毛虫, 为西藏温泉原生动物生物资源的发掘提供新的方法和思路, 也为原生动物环境适应性研究提供基础性资料。
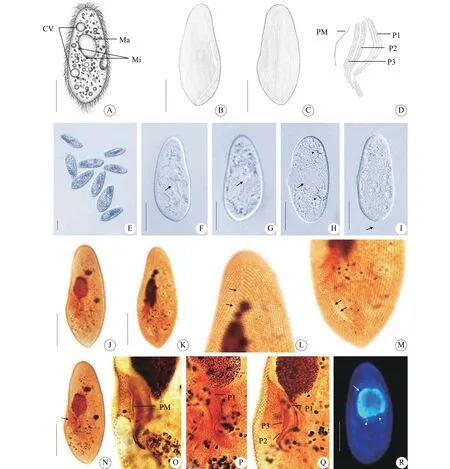
图版Ⅰ 第一双小核草履虫的形态学特征PlateⅠ Morphology of Paramecium primaurelia

图版Ⅱ 明布雷斯四膜虫的形态学特征PlateⅡ Morphology of Tetrahymena mimbres
