洱海漂浮草垫的物种组成及分布
2020-03-04赵雅璇齐亮宇候泽英吴爱平
赵雅璇 齐亮宇 候泽英 钟 文 刘 力 吴爱平
(1. 湖南农业大学生物科学技术学院, 长沙 410128; 2. 中国环境科学研究院湖泊创新基地, 北京 100012)
水位是决定植物生存的最重要的生态因子之一, 水位决定了湿地植物群落的组成以及分布, 因为不同的植物物种对水位有不同的反应和适应能力[1]。湖泊水位上升后, 会影响湿地植物的地上形态, 如植物的株高、茎节数、分枝数、叶面积、叶长和生长速率等[2], 使水生植物的生存环境发生变化, 从而使其个体数量、种群数量乃至整个生态系统的平衡状况随着环境的改变而改变。水位上升后, 水生植物光合作用有所抑制, 且高水位导致沉水植物大量消亡, 进而使得水质变混浊, 水体的pH降低, 溶解氧降低, 营养盐的浓度降低, 浮游植物的数量减少[3]。因而湖泊水位的变化对湖泊生态系统的健康和稳定有非常大的影响作用[4,5]。一般说来, 水生植物对水位变化主要有形态和生理的适应方式[6], 如果水位继续上涨, 超出了水生植物在形态和生理的适应限制, 大部分水生植物生长和繁殖将受到极大的限制, 甚至死亡。而很多挺水植物却能够形成漂浮草垫, 避免了水位上涨对其生长和繁殖的影响。先前研究表明在许多主要以挺水植物为优势物种的浅水沼泽和湿地中一些挺水植物如芦苇(Phragmites australis)、香蒲(Typha orientalis)等容易形成漂浮草垫[7], 但是在以沉水植物为优势物种的深水湖泊中, 哪些挺水植物更容易形成漂浮草垫却没见报道, 这也是本研究重点关注的问题。
洱海作为云南第二大高原湖泊, 其水生生态系统已从健康型向亚健康型退化[8], 20世纪80年代, 西洱河发电站的建立, 洱海长期在高水位运行, 大量水生植物因过高的水深而淹没致死, 整个水生生态系统受到严重影响。尤其是2003年水华发生后, 为缓解洱海水华的发生, 洱海管理局把洱海运行水位由1971 m调整到1974 m, 水位突然而急剧的抬升,对水生植物的生长和繁殖造成极大的影响[3,9]。在植物生存的适宜水深范围内, 植物生物量会随着水位的升高而增加, 但当水位过高时, 植物的生物量会显著减少[10]。水位变化过大或风浪过大容易造成水生植物连根拔起, 而且其幼苗也会受到损伤,不利于水生植物的生长[11]。一些挺水植物在高水位和高风浪作用下能形成漂浮草垫从而避免深水对其生长和繁殖的影响而存活下来, 因为形成漂浮草垫的挺水植物比其他有根植物对风浪等干扰更敏感, 且对水位变化有很好的适应能力[7]。湿地漂浮草垫的演替不仅导致水生植物物种组成上的改变, 还会引起生态群落随时间变化的更替[12]。随湖泊营养程度改变, 这种改变和更替更加明显, 并最终导致沉水植物被挺水植物和浮叶植物所替代[13]。由于漂浮草垫生长的野外环境复杂多样, 水位变化是影响漂浮草垫生长分布的一个重要因素, 国内外研究主要关注沼泽和浅水湿地的漂浮草垫, 通过对漂浮草垫群落的物种组成及分布模式调查, 其结果表明芦苇属(Phragmites)、香蒲属(Typha)、藨草属(Scirpus)、薹草属(Carex)、莎草属(Cyperus)等挺水植物更容易形成漂浮草垫[14]。这些研究都主要关注沼泽和浅水湿地这些水深相对较浅的湿地漂浮草垫群落的物种组成及分布, 而对深水湖泊如洱海最大水深超过20 m的湖泊还没有研究过。因此,有必要对水深相对较深的湿地也进行相似的研究,以摸清不同类型湿地漂浮草垫群落的物种组成及分布模式。先前的研究表明, 洱海的挺水植物为多优的群落, 而近些年来, 洱海挺水植物以菰(Zizania latifolia)为单优的群落[15]。另外, 在洱海野外调查过程中我们发现组成洱海漂浮草垫的挺水植物主要是菰(图1)。为此, 本研究就洱海水位上升的过程中, 通过分析洱海漂浮草垫群落的物种组成及分布模式, 探究挺水植物菰是否比其他挺水植物更能形成漂浮草垫而存活下来成为挺水植物群落中的单优种。

图1 洱海漂浮草垫野外调查情况Fig. 1 Observed floating mats in Lake Erhai
1 材料与方法
1.1 研究区概况
洱海四季如春, 属亚热带高原气候, 年平均气温为16.2℃, 全年日照时间2250—2480h[16]。洱海因其独特的生态地理特征和适宜的气候条件, 水生植被的物种丰富(38科, 76属, 100种)[17]。菰是洱海西岸分布面积最大, 生物量最多的挺水植物,它对洱海湖泊生态系统结构的稳定和生态功能的健康发挥起着重要的作用[8]。
1.2 试验方法
于2017年8月21日至2017年12月15日, 在云南洱海分夏季和冬季沿调查洱海分湖漂浮草垫的分布情况及植物组成类型(图2)。根据每个漂浮草垫面积的大小, 从草垫边缘向其中心分别取3—6个面积为1 m×1 m或者0.5 m×0.5 m的小区(面积小于1 m2的取整个草垫)。在采样处用测绳测量离岸距离,现场记录各漂浮草垫物种数、各物种的生物量、植物的最长根、面积、离岸边最近边缘的离岸距离和离岸边最近边缘的分布水深, 用便携式多参数水质分析仪(YSI-85)现场测量草垫水面下及草垫边缘20 m外开阔水体的温度(T)、pH和溶解氧(DO)等理化指标, 并采集草垫下及草垫边缘20 m外开阔水域的水样, 带回实验室测定水样的形态氮、形态磷等指标含量。样方内植物样品80℃烘干至恒重, 称取植物干重, 以干重代表生物量, 相对生物量=该种生物量/所有种生物量之和, 频度=物种出现的样方数/整个样方数×100, 结合不同漂浮草垫的分布面积, 估算全湖漂浮草垫的总生物量, 根据所测物种干重平均值和各物种盖度, 计算平均生物量。
水样中的营养盐指标分析按《水和废水监测分析方法》进行, 总氮(TN)采用碱性过硫酸钾消解紫外分光光度法; 硝态氮(-N)采用氨基磺酸分光光度法; 氨氮(-N)采用纳氏试剂比色法; 总磷(TP)、溶解性总磷(DTP)采用碱性过硫酸钾消解钼锑抗分光光度法; 亚硝态氮(-N)采用N-(1-萘基)-乙二胺分光光度法, 化学需氧量(COD)采用快速消解分光光度法。其中溶解性总磷、氨态氮、硝态氮、亚硝态氮需要在原水样经过0.45 μm孔径醋酸纤维素滤膜过滤后按上述方法测定。
1.3 数据分析
使用软件包SPSS 17.0进行统计数据分析。方差齐性用Levene检验, 均值之间的差异用 Duncan检验确定(P<0.05)。采用单因素方差分析方法, 分析了漂浮草垫物种数、生物量、平均生物量、最长根与面积的关系。采用单因素方差分析方法, 分析了夏季漂浮草垫和冬季漂浮草垫在总氮(TN)、总磷(TP)、溶解性总磷(DTP)、氨氮(-N)、硝氮(-N)、亚硝氮(-N)。

图2 洱海漂浮草垫位点分布Fig. 2 The observed sites with floating mats in Lake Erhai
2 结果与讨论
2.1 漂浮草垫分布规律
漂浮草垫面积与离岸距离的分布情况如图3所示, 夏季与冬季漂浮草垫的面积大小主要集中在200 m2以下, 呈碎片状, 形成的漂浮草垫在风浪作用下被分开并漂走, 最终漂流到风浪相对较小的湖边或者湖湾。漂浮草垫的面积相对较小, 这说明洱海漂浮草垫还处在形成的初期阶段, 或者形成的漂浮草垫很容易受到水位变化和风浪的影响, 被冲散而不再连接在一起, 因此很难形成大面积的漂浮草垫。大于1000 m2的漂浮草垫主要是由于参杂着大量的水葫芦(Eichhornia crassipes)和菱(Trapa)等漂浮植物。夏季漂浮草垫集中在离岸0—50 m的范围内,冬季漂浮草垫集中在离岸0—60 m范围, 漂浮草垫的数量随离岸距离增加而整体呈下降趋势, 夏季漂浮草垫主要集中在离岸距离10 m以内, 冬季漂浮草垫主要集中在离岸距离21—30 m。这主要是由于冬季水位上升后, 水面面积扩大, 导致冬季漂浮草垫的离岸距离比夏季的离岸距离远。
漂浮草垫水深的分布情况如图3所示, 夏季漂浮草垫的分布水深集中在0—250 cm, 冬季分布在51—200 cm, 夏季和冬季都主要集中在51—100 cm,夏季和冬季漂浮草垫的数量随水深的增加整体呈下降趋势。这主要是由于深水的开阔水域风浪相对较大, 不利于漂浮草垫稳定的定殖, 而水太浅的区域则容易受到回浪的影响, 同样也不利于漂浮草垫稳定的定殖[7]。因此, 漂浮草垫主要分布水深在50—200 cm的区域。
漂浮草垫物种丰富度的分布情况如图3所示夏季漂浮草垫的物种数在1—10种, 冬季漂浮草垫物种数在1—8种, 而夏季冬季漂浮草垫物种数都主要集中在3—4种, 这同样也说明洱海的漂浮草垫还处于形成的初期阶段, 而且漂浮草垫容易受风浪影响,导致很难形成大面积的漂浮草垫, 进而难以有足够的时间以保持有更多物种。
漂浮草垫植物最长根的分布情况本研究结果显示(图3), 夏季漂浮草垫植物最长根分布在0—160 cm,冬季漂浮草垫植物根长分布在0—120 cm。夏季和冬季的漂浮草垫中植物的最长根长都主要集中在40—120 cm, 最长根的长短可能主要与形成漂浮草垫的植物有关。而由于形成的漂浮草垫相对年轻或者由于风浪干扰经常被拆散, 因此最长根也集中于相对较短的区间。
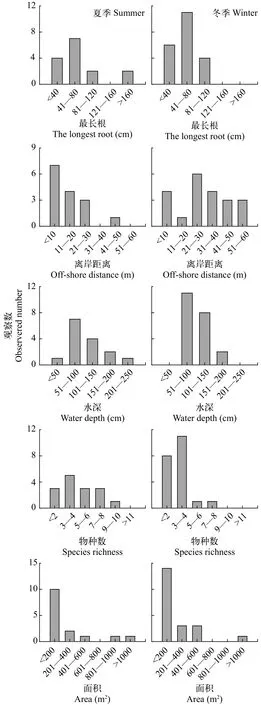
图3 漂浮草垫夏季冬季水深、最长根、离岸距离、物种丰富度、面积的分布观测数Fig. 3 Parameters of water depth, the longest root, off-shore distance, species richness and area of floating mat in summer and winter
2.2 草垫物种数、生物量、平均生物量、最长根与面积的关系
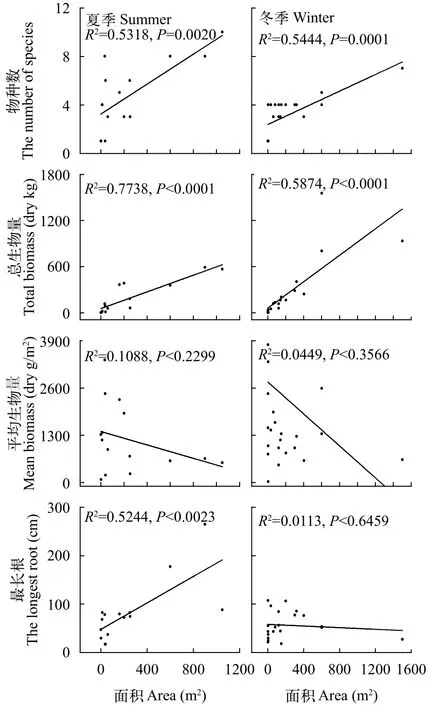
图4 物种数、生物量、平均生物量、最长根与面积的关系Fig. 4 The relationship between the number of species, total biomass, mean biomass, the longest root and area of floating mats
在本研究中, 夏季与冬季漂浮草垫上的物种数和生物量都与漂浮草垫的面积呈显著正相关(图4,P<0.05), 即随着漂浮草垫面积的增大, 漂浮草垫上物种数增多, 漂浮草垫总生物量也增大, 这与Ma-carthur等[18]的岛屿理论相符。而漂浮草垫的单位平均生物量与漂浮草垫的面积相关性不显著(图4,P>0.05)。因此尽管随着漂浮草垫面积的增加, 漂浮草垫的单位平均生物量有减小的趋势, 但漂浮草垫的总生物量还是随漂浮草垫面积的增加而增加。夏季漂浮草垫植物的最长根与漂浮草垫面积大小呈显著正相关(R2=0.5244,P<0.05), 而冬季漂浮草垫植物的最长根则跟漂浮草垫面积大小相关性不显著(R2=0.0113,P>0.05)。这可能是由于夏季漂浮草垫中面积越大的漂浮草垫里含有更多的菱,而菱的不定根很长, 水深的增加促使菱根系长度减小, 根系细长, 侧根增多, 促使夏季最长根和面积之间的正相关关系。但是, 冬季由于菱死亡以后, 所有的漂浮草垫中都不含有菱, 因此冬季漂浮草垫植物最长根与漂浮草垫面积大小相关性不显著。
2.3 草垫内外水质理化指标
漂浮草垫下物理化学指标的变化说明不同的植物采取不同的策略来适应草垫下环境条件的变化[19]。漂浮草垫下水体的总氮、总磷和溶解性总磷显著高于开阔水域, 而漂浮草垫下溶氧显著低于开阔水域(图5), 这与先前的研究结果一致[20]。漂浮草垫下水体N和P含量偏高主要是由于组成漂浮草垫的水生植物形成的大量凋落物以及产生的次生代谢产物被微生物分解释放到水体中所致[21], 而漂浮草垫下溶氧偏低则是由于微生物的活动导致水体中的溶氧被微生物所消耗[22]。洱海水体中营养盐浓度变化与运行水位息息相关, 湖泊运行水位高时, 湖泊各营养盐浓度低, 反之各营养盐浓度高[23]。水生植物不仅可以从底泥中直接获取营养盐也可以从水体中获取[24], 从水体中获取各营养盐导致各营养盐间接从底泥中释放, 进而增加了漂浮草垫下营养盐的浓度, 这可能也是造成漂浮草垫下水体N和P浓度高于开阔水域N和P浓度的原因之一。先前研究表明开阔水域氧气更容易溶解到水里, 浮游植物光合作用更强, 浮游植物光合作用有促于水体中的溶氧的增高, 该结果也解释了草垫下溶氧的浓度要低于周围开阔水域的溶氧浓度[25]。

图5 漂浮草垫夏季冬季水质理化指标Fig. 5 Physical and chemical indexes of water quality of floating mats in summer and winter
2.4 漂浮草垫的物种组成及优势种
漂浮草垫的物种组成本次调查夏季洱海漂浮草垫共18个, 冬季共22个, 共采集到植物15科, 19属,26个物种, 其中入侵物种4种, 本地种22种, 夏季物种24种, 冬季物种22种, 均为洱海的常见种(表1)。组成洱海漂浮草垫的浮叶植物有2种, 湿草植物6种,沉水植物8种, 漂浮植物7种, 挺水植物3种。生活型以沉水植物为主, 包括马来眼子菜(Potamogeton malaianus)、扭叶眼子菜(Potamogeton intortifolius)、篦齿眼子菜(Potamogeton pectinatus)、金鱼藻(Ceratophyllum demersum)、黑藻(Hydrilla verticillata)、穗花狐尾藻(Myriophyllum spicatum)、苦草(Vallisneria natans)、黄丝草(Potamogeton maackianus)。组成洱海漂浮草垫的挺水植物有芦苇、菰、香蒲。结果表明组成洱海漂浮草垫的植物种类主要以沉水植物为主, 而先前的研究都表明组成沼泽和湿地的漂浮草垫的植物种类主要以挺水植物为主,这可能主要由于洱海水深相对较深, 而且洱海具有相对较多的沉水植物种类, 而先前研究沼泽和湿地的沉水植物种类相对较少且处于非优势地位[26]。在洱海西洱河发电站建立以后, 一直保持高水位运行, 先前以菰、香蒲和芦苇为多优挺水植物群落现在也已经演变成菰为单优挺水植物群落。
漂浮草垫出现植物的频度及相对生物量调查发现洱海夏季漂浮草垫, 菰出现的频度最高(73.33%),其次是凤眼莲和水花生各占60.00%; 从相对生物量上来看菰也是最高的(41.91%), 其次是黄丝草和苦草各占21.97%和17.24%。冬季漂浮草垫频度菰最高(66.67%), 其次是凤眼莲和金鱼藻各占47.62%;从相对生物量来看菰也是最高的(43.38%), 其次是凤眼莲和绿狐尾藻各占31.16%和11.91% (表2)。该结果表明菰无论在夏季还是冬季, 它的频度和相对生物量都是最高的, 是形成洱海漂浮草垫挺水植物的单一优势物种, 说明菰在洱海水位变化(水位上升)过程中较其他挺水植物更易形成漂浮草垫。结果表明水位升高后, 菰发展出随水位升高形成漂浮草垫的生存策略, 该生存策略使得菰避免了深水对它的胁迫, 使其在水位变化和风浪的影响下依然能够存活下来, 从而相比其他挺水植物具有一定的竞争优势, 这也可能是菰成为洱海挺水植物群落单优物种的原因之一, 至于在水位上升过程中菰为什么比其他挺水植物更容易形成漂浮草垫的原理和机制则需要更进一步的研究。

表1 夏季和冬季漂浮草垫上植物名录Tab. 1 List of plants on floating mats in summer and winter
3 结论
(1)通过对洱海漂浮草垫物种组成的调查发现漂浮草垫上的植物都是洱海常见种, 且分布不均匀。夏季共调查到漂浮草垫18个, 植物分属15科,18属, 24种, 冬季共调查到漂浮草垫22个, 植物分属13科, 17属, 22种, 形成洱海漂浮草垫的植物总共15科, 19属, 26个物种。其中菰在夏季和冬季出现的频度和相对生物量都是最高的, 夏季菰出现的频度是73.33%, 相对生物量是41.91%, 冬季菰出现的频度是66.67%, 相对生物量是43.38%, 使菰在漂浮草垫挺水植物中成为了单一优势种。
(2)调查结果表明, 漂浮草垫上物种数、生物量都与漂浮草垫面积呈正相关, 漂浮草垫单位平均生物量与漂浮草垫的面积负相关性不显著, 夏季漂浮草垫植物的最长根与漂浮草垫面积大小呈显著正相关, 而冬季漂浮草垫植物的最长根则跟漂浮草垫面积大小相关性不显著。
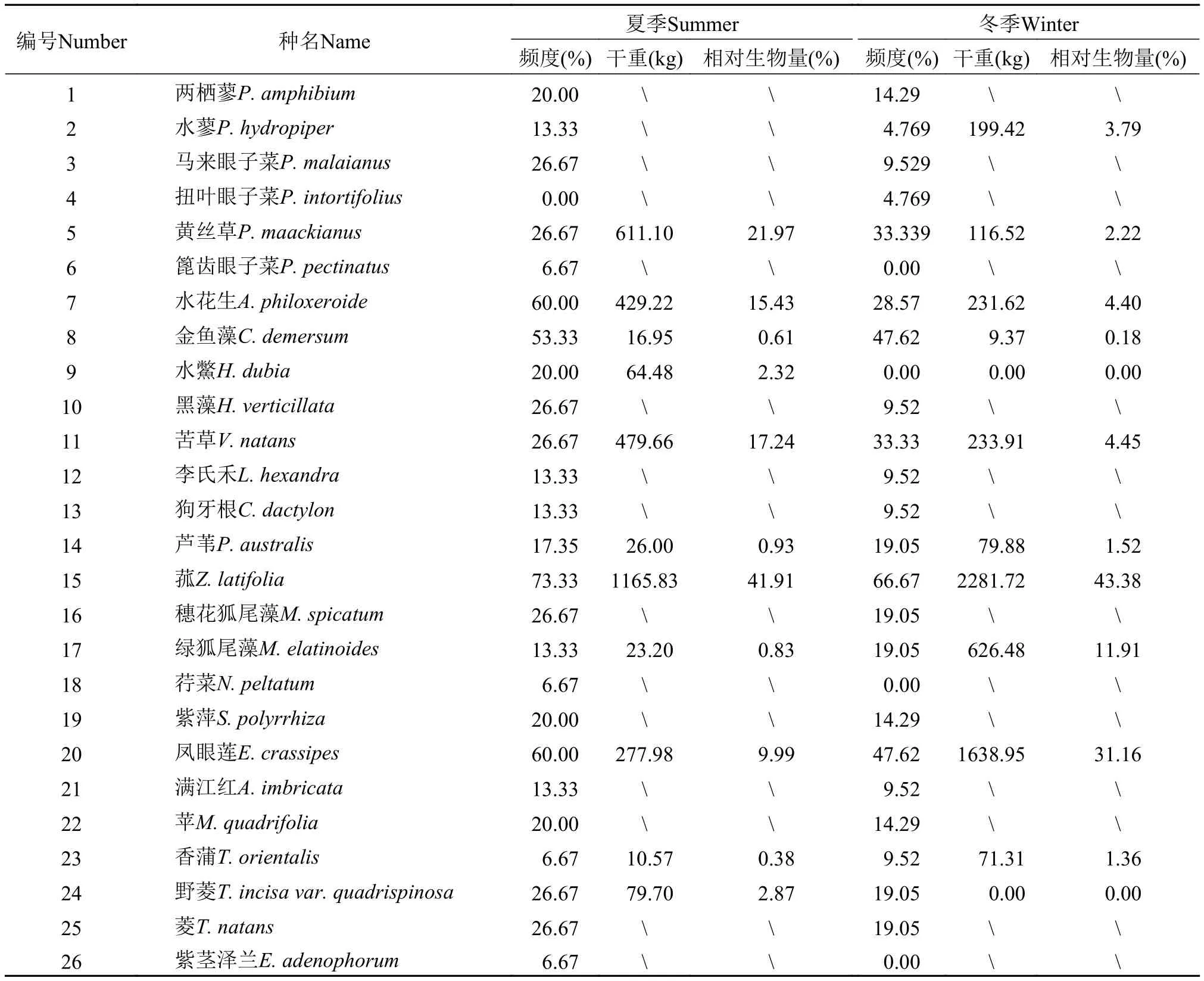
表2 夏季和冬季漂浮草垫植物出现频度及相对生物量Tab. 2 Frequency and relative biomass of floating mats in summer and winter
(3)调查结果显示漂浮草垫面积主要集中在200 m2以下, 水深主要集中在50—100 cm, 夏季漂浮草垫主要集中在离岸距离10 m以内, 冬季漂浮草垫主要集中在离岸距离21—30 m内, 夏季和冬季的漂浮草垫中植物的最长根长都主要集中在40—120 cm。漂浮草垫下水体中总氮(TN)、总磷(TP)、溶解性总磷(TDP)都明显高于开阔水域的(P<0.05), 而开阔水域的溶氧(DO)明显高于草垫下水体的(P<0.05)。
(4)这些结果表明洱海漂浮草垫还处于形成的早期阶段, 在水位上升过程中菰比其他挺水植物更容易形成漂浮草垫的原理和机制则需要更进一步的研究。
