蒙古寒蝉末龄若虫与成虫复眼的形态、组织结构和超微结构
2020-03-04兰英,魏琮
兰 英, 魏 琮
(西北农林科技大学植物保护学院, 植保资源与病虫害治理教育部重点实验室, 陕西杨凌 712100)
节肢动物广泛地分布在陆地或水生栖息地中(Alfsnesetal., 2017)。复眼是节肢动物的重要视觉器官,在觅食、求偶、感知光线、定位和导航等行为中具有重要作用(Nilsson, 1989; Srinivasanetal., 1999; Floreano and Zufferey, 2010; Chapman, 2013; Lehmannetal., 2017)。在昆虫纲中,绝大多数成虫和半变态类若虫都具有1对发达的复眼(Chapman, 2013);复眼的形态结构与昆虫对光亮的适应调控密切相关,因此是解析昆虫对光亮感受行为及相关生理反应机制研究的基础(蒋月丽等, 2016)。
昆虫的复眼由数目不等的小眼构成,每个小眼由角膜、晶锥、视杆、基膜等组成(Nilsson 1989; Oakley, 2003)。生活在不同环境中的昆虫,复眼表面感器形态及小眼数目和结构存在差异。例如,一些善于飞行的蝇类,其复眼表面具有浓密的刚毛(Stumm-Tegethoff and Dicke, 1974);稻赤斑黑沫蝉Callitettixversicolor小眼之间随机分布一些稀疏的感觉毛(Jia and Liang, 2015);一些夜行性昆虫复眼的表面则分布许多微小的乳头状凸起(Bernhardetal., 1970);红火蚁Solenopsisinvicta小眼的数目及感觉毛的形状与数量在不同性别及品级之间存在显著差异,可能与不同个体的生活习性和活动环境有关(范凡等, 2008)。根据成像方式,昆虫复眼可被分为并列像眼和重叠像眼两种类型(Völkeletal., 2003; Nation, 2008; Mehdipouretal., 2015)。通常在日间活动的昆虫,如善于飞行的蜜蜂和蜻蜓等,其复眼为并列像眼(Chapman, 2013);这类昆虫对光的敏感性较低,但视觉的分辨率较高(Horridge, 1977; Land, 1997)。在并列像眼中,复眼内部的视杆与晶锥紧密连接,小眼内小网膜细胞中的色素颗粒可随光亮变化而发生移动来调节进光量,从而适应不同的光环境(闫海燕等, 2006)。夜间活动的昆虫,如蛾类和一些甲虫,其复眼为重叠像眼;该类昆虫对光具有较高的敏感性,但分辨率较低;其复眼内部的视杆与晶锥未直接相连,存在一透明区域,也可以通过色素细胞及小网膜细胞内的色素颗粒移动来调节进光量(Horridge, 1977; Nilsson, 1983; Land and Nilsson, 2012)。昆虫复眼的结构与其行为之间关系错综复杂,因此它们的复眼被认为是研究生物视觉生理和行为反应的最佳模型(Evangelinetal., 2015)。
半翅目(Hemiptera)被分为头喙亚目(Auchenorrhyncha)、胸喙亚目(Sternorrhyncha)、鞘喙亚目(Coleorrhyncha)及异翅亚目(Heteroptera)(Cryan and Urban, 2012; Lietal., 2017)。以往少量的半翅目昆虫复眼研究大多集中在异翅亚目(Settembrini, 1984; Fischeretal., 2000; Reisenman, 2002; Mishraetal., 2019),其他类群复眼的研究一直比较有限。关于头喙亚目昆虫复眼的研究非常缺乏,截至目前仅有数种沫蝉总科(Cercopoidea)昆虫的复眼形态结构得到研究(Ramachandran, 1963; Keskinen and Meyer-Rochow, 2004; Jia and Liang, 2015; Ranierietal., 2016)。头喙亚目的蝉总科(Cicadoidea)若虫长期在土壤中营固定生活(最长可达17年之久)(周尧等, 1997),以刺吸式口器吸食寄主植物幼根汁液,经5个龄期发育后出土羽化(Houetal., 2015),成虫则在树冠生活,取食寄主枝干汁液(周尧等, 1997)。蝉总科昆虫与头喙亚目的沫蝉总科和角蝉总科昆虫相比,其成虫与若虫在个体发育中发生了非常显著的生态位改变(由黑暗环境转移到明亮的环境),但关于该总科昆虫的视觉器官形态和结构研究一直缺乏。
本研究采用光学显微镜和扫描及透射电子显微镜技术,比较研究了中国广泛分布的蒙古寒蝉Meimunamongolica不同发育阶段的末龄(5龄)若虫与成虫复眼的形态、组织结构和超微结构,旨在明晰其复眼结构特征及其在出土羽化前后可能出现的变化,以便为该类群的视觉生理和相关行为研究奠定基础。
1 材料与方法
1.1 标本采集
蒙古寒蝉M.mongolica的成虫及若虫均于2019年8月采集于陕西杨凌西北农林科技大学校园(34°16′N, 108°4′E)。成虫采用网捕法采集,采获后带回实验室解剖研究。若虫通过在寄主植物油松Pinustabuliformis和杜梨Pyrusxerophila树冠下的土壤中挖取获得,采获后立即连同适量土壤一起放入铝盒中(黑暗环境)带回实验室,通过体型大小、体色、头宽确定末龄若虫(即5龄若虫)(Houetal., 2015),备用。
1.2 扫描电子显微镜样品制备及观察
解剖观察之前,先对蒙古寒蝉成虫和具有不同颜色复眼的末龄若虫(各9头)进行冷冻麻醉处理约10 min,然后在Nikon SMZ 1500光学显微镜下,将复眼连同头部一同取下,用2%多聚甲醛和2.5%戊二醛混合固定液(10%多聚甲醛∶25%戊二醛∶0.2 mol/L磷酸缓冲溶液∶蒸馏水=2∶1∶5∶2, v/v; pH 7.2)在4℃固定24 h。固定后,用0.1 mol/L磷酸缓冲液(pH 7.2)清洗样品3次,每次10 min。然后分别在30%, 50%, 70%, 80%和90%乙醇中进行梯度脱水各10 min;再用100%乙醇脱水3次,每次30 min。脱水后,用乙酸异戊酯置换2次,各30 min;之后经过CO2临界点干燥后,用双面导电胶将样品固定到样品台上,放入真空喷涂仪中喷金5 min后,置于Hitachi S-3400N型扫描电镜下观察并拍照。
1.3 透射电子显微镜样品制备及观察
将蒙古寒蝉成虫和具有不同颜色复眼的末龄若虫(各9头)头部迅速斩断并剖下完整复眼,置于2%多聚甲醛和2.5%戊二醛混合溶液(10%多聚甲醛∶25%戊二醛∶0.2 mol/L磷酸缓冲溶液∶蒸馏水=2∶1∶5∶2, v/v; pH 7.2)中在4℃固定24 h。固定后的样品用磷酸缓冲液(0.1 mol/L, pH 7.2)漂洗5次,每次15 min。漂洗后的样品用1%四氧化锇(OsO4)固定液在4℃下避光固定2 h,再用磷酸缓冲液漂洗5次,每次15 min;然后用30%, 50%, 70%, 80%和90%乙醇逐级脱水,每次10 min;再用100%的乙醇脱水3次,每次20 min;随后,用3个不同体积比例的无水乙醇和EPON 812树脂混合物(乙醇∶EPON 812=3∶1, 1∶1和1∶3, v/v)依次渗透2, 4和12 h,再用纯EPON 812树脂渗透2次,每次24 h。渗透好的样品用纯EPON 812树脂进行包埋,将包埋好的样品置于在30℃烘箱中聚合24 h,再在60℃下聚合48 h。
聚合完成后,对包埋胶粒进行适当修整,然后在Leica EM UC7切片机上进行半薄切片(约2 μm)并定位,再用钻石刀进行超薄切片(约70 nm);切片再分别用2%醋酸铀和4%柠檬酸铅染色(分别为20和10 min);将切片分别置于HITACHI H-7650 型透射电镜下,在80 kV下对切片进行观察并拍照。
1.4 光学显微镜观察
将麻醉的蒙古寒蝉成虫及具有不同颜色复眼的末龄若虫(各9头)用昆虫针固定于泡沫板上,置于Nikon SMZ 1500光学显微镜下观察其复眼形态,并采用QIMAGING RETIGA2000RCCD进行拍照,用Montage软件叠加合成图片。另外,将透射电镜制备的样品块在Leica EM UC7切片机上进行半薄切片(厚度约2 μm),然后将半薄切片置于载玻片上,用0.5%的甲苯胺蓝染液染色,随后在连有CCD成像系统的Olympus BX-51型光学显微镜下进行观察并拍照。
在Nikon SMZ 1500光学显微镜下将蒙古寒蝉成虫及具有不同颜色复眼的末龄若虫(各9头)头部取下,放入磷酸缓冲液(0.1 mol/L, pH 7.2)内,用镊子和解剖剪取下完整复眼,再用磷酸缓冲液(0.1 mol/L, pH 7.2)清洗3次,每次10 min;然后用超声波清洗3次(每次4~5 s),除去附着在复眼表面的灰尘等杂物;随后用2%多聚甲醛和2.5%戊二醛混合溶液(10%多聚甲醛∶25%戊二醛∶0.2 mol/L磷酸缓冲溶液∶蒸馏水=2∶1∶5∶2, v/v; pH 7.2)在4℃固定24 h;将固定后的样品用30%,50%, 70%, 80%, 95%和100%乙醇进行梯度脱水每次40 min;之后用透明剂透明30 min;最后浸蜡3 h,选择合适角度对样品进行包埋,包埋完成后即可进行切片(厚度约为4 μm)。通过初检选择切取角度合适、质量过关的切片,进行贴片、烤片及复水,然后用苏木精-伊红双染色法(HE)染色,封片后在连有CCD成像系统的Olympus BX-51型光学显微镜下观察并拍照。
本研究中复眼小眼的结构及术语根据Friedrich等(2011)。
2 结果
2.1 复眼外部形态
蒙古寒蝉末龄若虫与成虫的复眼均为半球形(图1; 图2: A-D)。末龄若虫的复眼在出土羽化前逐渐由白色变为红色至深褐色(图1: A-C),成虫的复眼则为浅褐色(图1: D)。末龄若虫复眼表面覆盖一层坚硬的外膜(羽化后成为蝉蜕的一部分),白色复眼的腹侧分布有稀疏的毛形感器、锥形感器和刺形感器:毛形感器较纤细,端向渐细,顶端较钝(图2: E, F);锥形感器较短小,直立,顶端较钝,基部无臼状窝(图2: G);刺形感器短小,直立,顶端较尖,基部毛窝较宽(图2: H)。解剖发现,末龄若虫白色复眼的坚硬外膜与内部组织未发生分离(难以用昆虫针及镊子将外膜与内部组织剥离),复眼表面未分化形成形态和边界清晰的小眼面(图2: A);末龄若虫红色和深褐色复眼表面的外膜可用昆虫针和镊子剥离;去除外膜后的复眼表面光滑,无任何感器,但具有明显的小眼,呈不规则的六边形或五边形;红色复眼的小眼面比深褐色复眼的小眼面形状更不规则(图2: I, J)。成虫复眼表面光滑,由排列整齐的等边六边形小眼组成,小眼面微外凸,无任何感器(图2: K)。

图1 蒙古寒蝉末龄若虫与成虫的头部及复眼

图2 蒙古寒蝉末龄若虫与成虫的复眼外部形态及超微结构
2.2 复眼的组织结构
若虫白色复眼内部由大量细胞团构成,未形成小眼结构(图3: A)。这些细胞团分化明显,近心端细胞层中的细胞较小且密集(图3: A红色箭头所示, C);远心端细胞层中的细胞较大,排列松散(图3: A, 黑色箭头所示)。角膜(cornea)下方排列着众多角膜形成细胞,表明旧的角膜与新的角膜即将分离(图3: B, 无尾箭头所示);在视叶与近心端的细胞团之间分布一些柱状细胞(图3: D, 矩形框所示)。
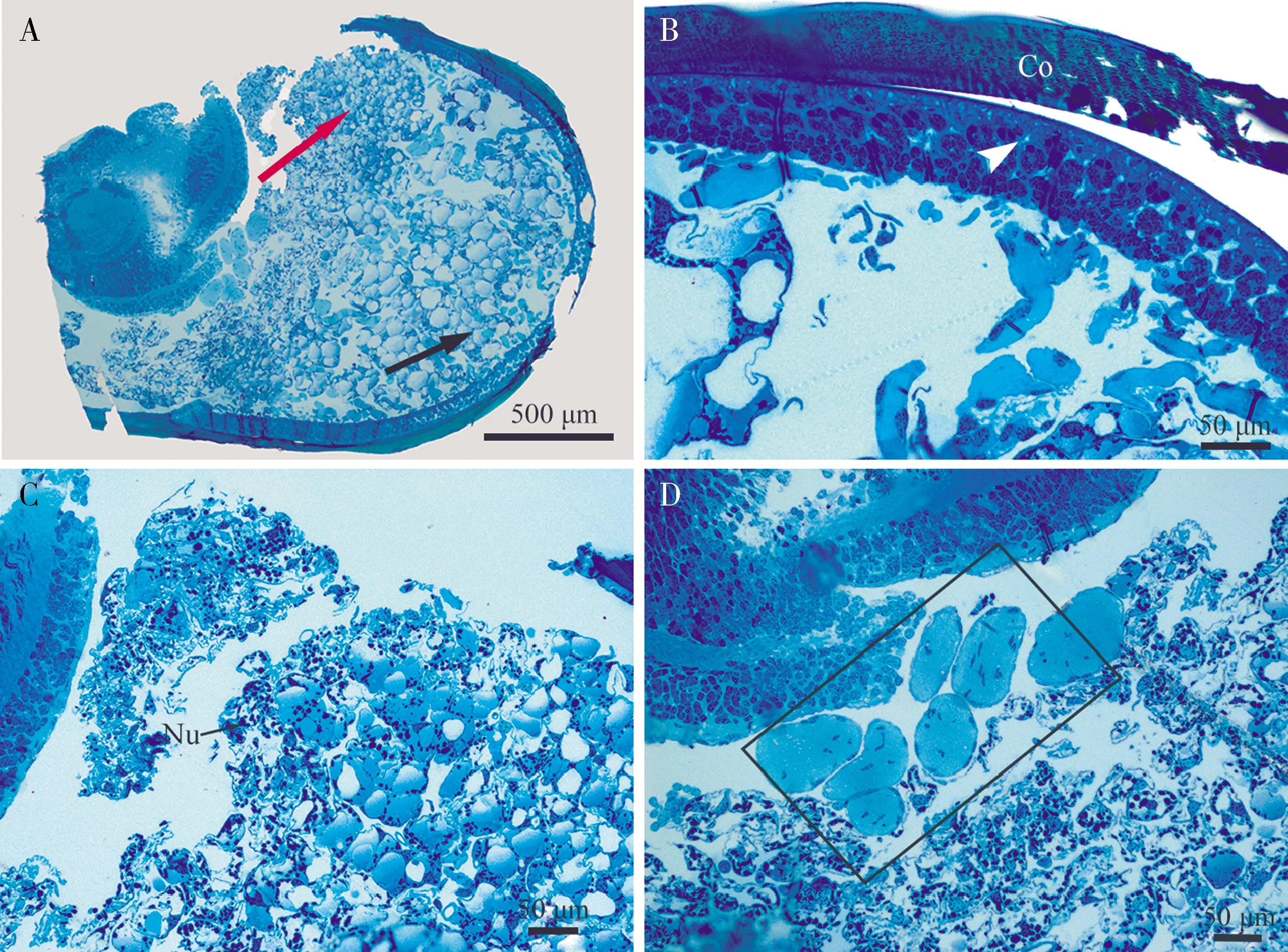
图3 蒙古寒蝉末龄若虫白色复眼组织结构观察
成虫复眼的小眼和若虫深褐色复眼的小眼都由角膜、晶锥(crystalline cone)、小网膜细胞(retinula cells)和基膜(basal membrane)组成(图4: B, E)。角膜向外凸起,向内与晶锥紧密接触(图4: A, D)。晶锥呈倒圆锥形,晶锥细胞(cone cells)的细胞核分布在晶锥细胞的最远端(图4: B, C, E, F)。在晶锥下方,小网膜细胞和视杆(rhabdom)构成感光器;小网膜细胞的细胞核分布在远心端和中间部分(图4: B, C, E, G)。在若虫深褐色复眼中,晶锥与视杆远端直接相连,但次级色素细胞(secondary pigment cells)的细胞核都集中分布在晶锥近心端和视杆远端之间(图4: B, C)。在成虫复眼中,晶锥的基部与视杆远端也直接相连,次级色素细胞的细胞核大都分布在晶锥近心端周围(图4: E, F)。
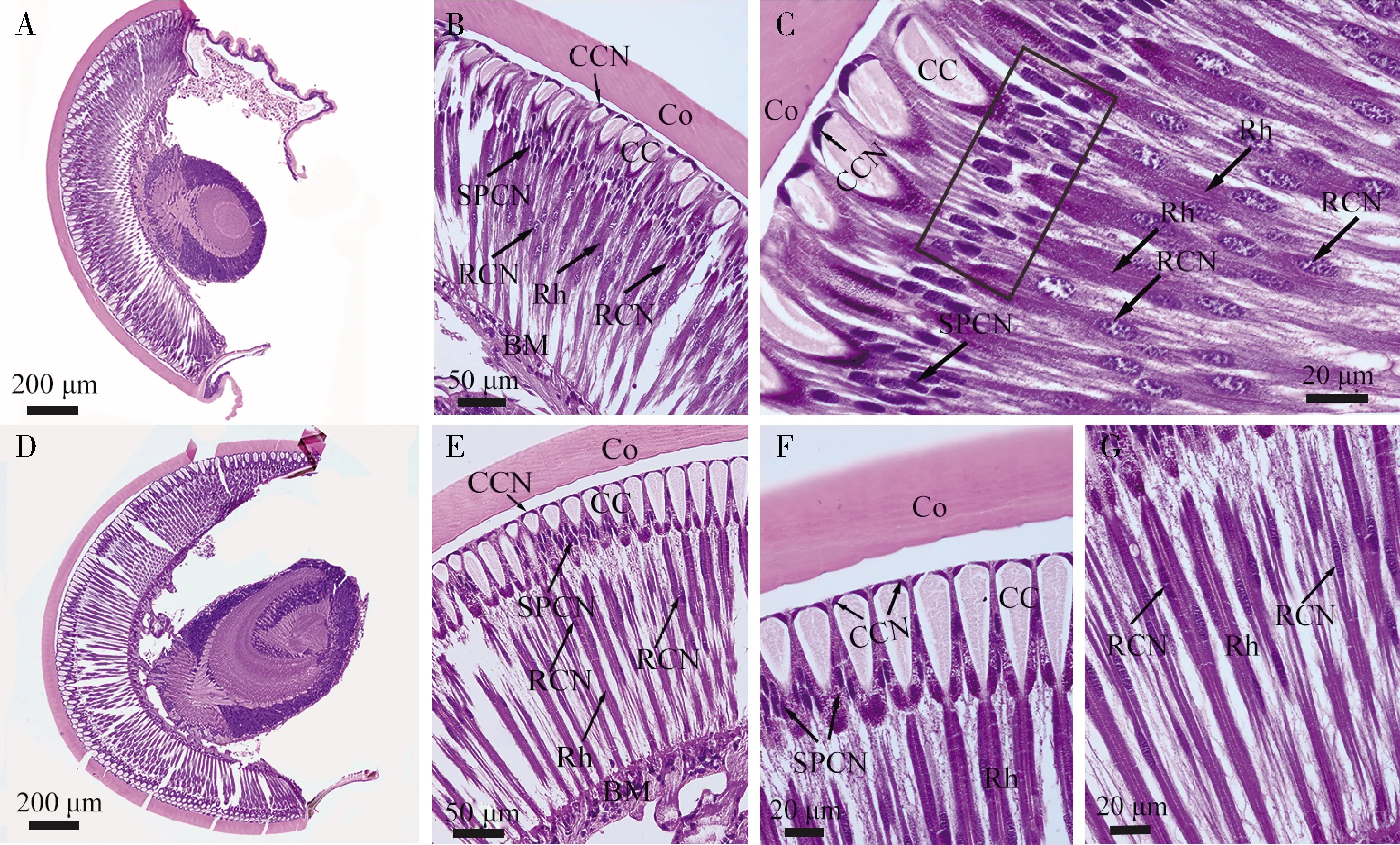
图4 蒙古寒蝉末龄若虫深褐色复眼及成虫复眼组织结构观察
2.3 复眼的超微结构
若虫白色复眼的角膜呈分层构造,外层排列较疏松,内层排列紧密(图5: A)。角膜生成细胞与角膜紧密相连,细胞膜在连接处特化成微绒毛状(图5: B, 箭头所示)。角膜生成细胞内含有高尔基体和线粒体(图5: B)。角膜生成细胞的细胞核分布在角膜生成细胞的基部,成簇存在(分别为“5-细胞核簇”和“8-细胞核簇”);细胞核周围分布有大量囊泡(vesicle)(图5: C, D)。

图5 蒙古寒蝉末龄若虫白色复眼超微结构
若虫深褐色复眼和成虫复眼的小眼晶锥均由4个晶锥细胞组成,为典型的晶锥眼;晶锥横切面呈圆形,周围包裹着1对初级色素细胞。初级色素细胞内含有色素颗粒,但若虫小眼初级色素细胞中的色素颗粒密度明显大于成虫。初级色素细胞外围分布着多个次级色素细胞,并填满相邻细胞间的空间(图6: A; 图7: A)。晶锥细胞向下延伸并插入小网膜细胞的空隙中,每个小眼基部都包含8个小网膜细胞(R1-R8);小网膜细胞的细胞膜的内表面特化形成微绒毛,微绒毛伸展方向一致,聚合形成视小杆,逐渐向晶锥中心延伸(图6: B; 图7: B)。视杆近心端的横切面近圆形,晶锥在视杆的中段消失(图6: C, D; 图7: C, D)。在若虫的深褐色复眼中,视杆为半融合型,在靠近小网膜细胞R8内表面处形成了瞳孔(图6:C, D);但成虫复眼的视小杆沿小眼纵轴聚合形成闭合的视杆(图7: D)。若虫深褐色复眼和成虫复眼的小网膜细胞内均有色素颗粒分布,但若虫小网膜细胞中的色素颗粒分布均匀且密度较大(图6: B, C),成虫小网膜细胞内的色素颗粒多集中在靠近视杆周围的栅栏层(palisade)附近(图7: B, C)。
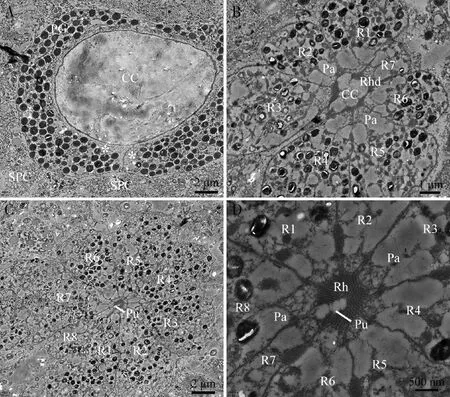
图6 蒙古寒蝉末龄若虫深褐色复眼的小眼超微结构

图7 蒙古寒蝉成虫小眼超微结构
3 讨论
昆虫复眼的形态和结构不仅与昆虫的种类及性别相关,也可能与其生存环境和习性密切相关(Jander and Jander, 2002)。有些昆虫复眼表面常具有刚毛等特殊构造,例如西方蜜蜂Apismellifera的复眼表面具有浓密刺毛,有助于觅食时准确判断取食场所和太阳之间的方向角度(Neese, 1965);东方胡蜂Vespaorientalis的复眼表面分布大量的圆形凸起,具有抗光线反射的作用(Litinetskyetal., 2002);而一些夜行性昆虫复眼表面具有许多小的乳头状凸起,也具有抗光线反射、增加入射光强度的作用,以便于夜间活动(Bernhardetal., 1970);直翅目昆虫双斑蟋Gryllusbimaculatus的复眼表面具有少量的梨状或卵圆形感器,主要作用为感受环境中的机械压力(那宇鹏等, 2015)。本研究结果表明,蒙古寒蝉末龄若虫和成虫复眼属于并列像眼;末龄若虫复眼表面具有毛形感器、锥形感器和刺形感器(图1: E-H),但成虫复眼表面无感器。这应是蝉总科昆虫在从若虫发育为成虫过程中对生态位显著改变的一种适应——长期在地下漆黑环境营固定生活的若虫可通过复眼表面的感器感受土壤砂砾或寄主植物根系的机械压力,并产生应激行为;成虫则脱离地下漆黑、密闭狭窄空间而生活于地上植物树冠。
小眼是复眼的结构和功能单元,每个小眼都是一个高度分化的独立视觉单元(蒋月丽等, 2016);小眼数量越多、夹角越小,复眼成像越清晰(陆苗等, 2013)。本研究发现蒙古寒蝉末龄若虫白色复眼的表面尚未形成小眼面,内部也仅由大量细胞团构成,也未分化形成小眼结构。据此可以推断,具有白色复眼的蒙古寒蝉末龄若虫尚处于该龄期的早期发育阶段,此阶段及更早的低龄若虫不具备视觉能力,但可通过复眼表面的感器及其他部位的感器感受外界机械压力或其他环境信号。此外,本研究发现末龄若虫红色和深褐色复眼表面的外膜已与内部细胞完全分化并可轻易被剥离;去除外膜后的复眼表面无任何感器,却已形成明显的小眼面,而红色复眼小眼面的形状、排列与深褐色复眼的小眼面相比更不规则。这些差异表明红色复眼期是其复眼的主要发育阶段,而深褐色复眼期为末龄若虫的发育晚期,其复眼仍在进一步发育。蒙古寒蝉末龄若虫复眼颜色及小眼形态、结构变化应与该龄期若虫即将出土羽化密切相关;关于若虫红色复眼的组织、超微结构及发育调控机制有待进一步深入研究。
并列像眼的主要特征是小眼的视杆与晶锥紧密相连,可通过小眼色素细胞移动对进光量进行调节;这类复眼对光的敏感性较低,但具有较高的分辨率(Warrant and McIntyre, 1993)。例如,在暗适应条件下,菜粉蝶Pierisrapae复眼中的色素颗粒集中在视网膜远心端而靠近晶锥,但在光适应条件下,色素颗粒聚集在视杆周围,从而通过色素颗粒的移动来调节进光量,以适应不同光强度的变化(Ribi, 1978)。夜间活动的昆虫,如蛾类和一些甲虫,其复眼多为重叠像眼,对光具有较高的敏感性,但分辨率较低,其复眼内部的视杆与晶锥之间有间隙而未直接相连,当昆虫由光亮地方移动到黑暗处活动时,其复眼可以通过色素细胞及小网膜细胞内的色素颗粒移动来调节进光量(Horridge, 1977; Nilsson, 1983; Land and Nilsson, 2012)。本研究发现,蒙古寒蝉成虫复眼和末龄若虫深褐色复眼的小网膜细胞和色素细胞内均含有色素颗粒,但在色素颗粒的分布上有明显差异:末龄若虫深褐色复眼的初级色素细胞及小网膜细胞内的色素颗粒大于成虫的色素颗粒,而且小网膜细胞内的色素颗粒分布较均匀;成虫视网膜细胞内色素颗粒密度相对较小,且多集中分布在栅栏层周围。这些差异也应与成虫和若虫的生态位显著分化密切相关——若虫生活于光强度恒定的黑暗环境中,而成虫的生境则与此相反,因此需要通过复眼色素细胞及小网膜细胞内的色素颗粒移动来调节进光量,以适应生境的光线强弱变化。
Fischer等(2000)研究发现半翅目中所有异翅亚目昆虫复眼的视杆都为开放型,而头喙亚目和胸喙亚目昆虫复眼的视杆都为闭合型。本研究发现,蒙古寒蝉成虫复眼的视杆为闭合型,但若虫深褐色复眼的视杆在中心形成了明显的瞳孔。吴卫国等(1990)关于直翅目蝗虫和螽斯成虫复眼的研究表明,瞳孔对小眼的进光量具有调节作用。其他相关研究表明,一些昆虫小眼的瞳孔闭合还具有昼夜节律变化,如染锥猎蝽Triatomainfestans(Reisenmanetal., 2002)、黄粉虫Tenebriomolitor(Wada and Schneider, 1968)和蝇类(Ro and Nilsson, 1994)的复眼。深褐色复眼期是蒙古寒蝉末龄若虫在地下生活的最后阶段,该时期的若虫都生活在非常靠近地表的浅层土壤中,其瞳孔应与即将发生的羽化行为密切相关——可通过视杆的扩张及收缩调节作用来适应出土羽化过程中的光线强度改变,但瞳孔的闭合是否存在昼夜节律仍然未知。瞳孔在蝉科昆虫若虫复眼中是否普遍存在,其他蝉科昆虫成虫的复眼是否具有瞳孔,这些问题都有待进一步研究予以明晰。
综上所述,本研究对蒙古寒蝉不同发育阶段的末龄若虫与成虫的复眼进行了形态、组织结构和超微结构比较研究。相关结果为蝉总科昆虫复眼的发育及功能变化与生境的相关性研究提供了新的重要信息,同时也为进一步探讨蝉总科与半翅目其他类群的系统演化提供了功能形态学方面的新证据。
