蛾类性信息素受体研究进展
2020-03-04王桂荣
曹 松, 刘 杨, 王桂荣
(中国农业科学院植物保护研究所, 植物病虫害生物学国家重点实验室, 北京 100193)
雄蛾通过识别同种雌蛾释放的特异的性信息素来寻找合适的配偶,并完成后代繁衍。蛾类的性信息素一般是由雌蛾(仅少数雄蛾)特定的腺体合成并释放到体外的一类挥发性的化合物,常为不同比例的多种化合物构成的混合物(Byer, 2006),这类混合物能引起同种雄蛾特定的行为或生理反应(Karlson and Butenandt, 1959; Karlson and Lüscher, 1959; Roelofs and Cardé, 1977),同时对种间生殖隔离的形成具有重要作用(Grootetal., 2006; Mingetal., 2007; Baker, 2008; Smadja and Butlin, 2009)。1959年,科学家从家蚕Bombyxmori中鉴定了蛾类的第一种性信息素成分——蚕蛾醇(Butenandtetal., 1959)。在过去60年中,超过1 600种蛾类的性信息素成分得到鉴定(Grootetal., 2016)。根据化学结构,蛾类的性信息素可分为4种类型:Ⅰ,Ⅱ,Ⅲ及0型。Ⅰ型性信息素通常是由10~18个碳原子构成的包含0~3个不饱和双键的酯类、醇类或醛类化合物(Millar, 2000; Andoetal., 2004; Föstedtetal., 2016),如家蚕的性信息素成分蚕蛾醇E10,Z12-16∶OH(Butenandtetal., 1959)。这类化合物约占蛾类性信息素的75%。典型的Ⅱ类性信息素是由17~25个碳原子构成的长链不饱和碳氢化合物或环氧衍生物,常包含1~3个不饱和双键和0~2个环氧基(Millar, 2000; Andoetal., 2004; Grantetal., 2012; Föstedtetal., 2016),如冬尺蠖蛾Operophterabrumata的性信息素成分1, 3Z, 6Z, 9Z-19∶Hy(Roelofsetal., 1982)。Ⅱ类性信息素约占蛾类性信息素的15%。Ⅲ类性信息素的结构与Ⅰ和Ⅱ类性信息素不同,典型的Ⅲ类性信息素通常包含1或2个甲基侧链分支,并且甲基支链被奇数个碳原子隔开(Föstedtetal., 2016),如舞毒蛾Lymantriadispar的性信息素成分(7R, 8S)-7,8环氧-2-甲基十八烷(Bierletal., 1970)。0型性信息素主要是一类由7~9个碳原子构成的短链醇类或酮类化合物,这类化合物结构比较简单,更类似于植物挥发物(Föstedtetal., 2016),例如高山毛顶蛾Eriocraniasemipurpurella的性信息素成分(S,Z)-6-nonen-2-ol(Kozlovetal., 1996)。目前这类化合物仅在鳞翅目古老的单孔亚目(Monotrysia)及毛翅目(Trichoptera)中有报道,因此也常被看作是蛾类性信息素的起源。
雄蛾主要通过触角上灵敏的嗅觉系统来识别性信息素(Vogt and Riddiford, 1981)。昆虫触角上分布着不同类型的嗅觉感器,通常情况下,每根感器下包含1~4个不等数量的嗅觉受体神经元(olfactory receptor neuron, ORN),性信息素受体(pheromone receptor, PR)表达在毛形感器内ORN的树突膜上。蛾类性信息素识别是一个复杂的过程,需要多种蛋白和神经元细胞参与。首先,性信息素分子通过毛形感器表面的孔洞进入感器内部,脂溶性的性信息素分子被性信息素结合蛋白(pheromone binding protein, PBP)捕获结合,并运输至PR,PR被激活后打开离子通道,将化学信号转化为电信号,电信号被OSN的轴突传递到触角叶内的嗅小球,信号在触角叶中完成加工,而后被投射神经元(projection neuron, PN)传递到更高级的脑中枢。信号经过高级脑中枢的整合、处理,指导雄蛾对性信息素做出电生理或行为反应。在外周嗅觉分子水平上,除了PR(Sakuraietal., 2015; Changetal., 2017)和PBP(Große-Wildeetal., 2006; Allen and Wanner, 2011; Zhu Getal., 2016)以外,感受神经元膜蛋白(sensory neuron membrane protein, SNMP)(Pregitzeretal., 2014; Blankenburgetal., 2019)和性信息素降解酶(pheromone-degrading enzyme, PDE)(Rybczynskietal., 1989; Ishida and Leal, 2005; Durandetal., 2011; Chooetal., 2013)等多种蛋白也参与了蛾类性信息素的识别过程。另外,有研究发现果蝇Drosophila的离子型受体(ionotropic receptor, IR)也参与了性信息素的识别过程(Kohetal., 2014; Heetal., 2019),但是蛾类IR是否直接参与性信息素的识别还未见报道。
尽管多种嗅觉相关蛋白参与了蛾类性信息素的识别过程,但是PR作为与性信息素直接结合并产生信号转导的蛋白,决定了ORN对配体的选择性和特异性。本文综述蛾类PR的鉴定、结构特征、表达模式、功能研究以及进化等方面的研究进展,以期为蛾类PR的功能和进化研究提供参考,同时为开发蛾类害虫基于性信息素受体的绿色防控措施提供理论依据。
1 蛾类昆虫性信息素受体的鉴定及结构特征
1991年,科学家从脊椎动物褐家鼠Ratfmnorvegicus的嗅觉上皮细胞中发现第一个气味受体(odorant receptor, OR)基因(Buck and Axel, 1991),它属于G蛋白偶联受体(G protein-coupled receptor, GPCR)超家族成员。1999年,通过全基因组测序的方法,研究人员在果蝇中首次鉴定到昆虫的OR基因(Clyneetal., 1999; Gao and Chess, 1999; Vosshalletal., 1999),为昆虫OR的鉴定和研究提供了理论基础。昆虫的OR一般由350~450个氨基酸编码,是一类七次跨膜蛋白(seven-transmembrane domain protein),但它并不属于GPCR家族成员,因为它们具有与GPCR相反的拓扑结构,昆虫OR的N端在胞内而C端在胞外(Bentonetal., 2006; Lundinetal., 2007)。此外,昆虫特异的OR需要和一个种间高度保守且广泛表达的气味受体共受体(odorant receptor coreceptor, Orco)(Larssonetal., 2004; Jonesetal., 2005; Neuhausetal., 2005)形成异源OR-Orco多聚复合体来行使功能(Neuhausetal., 2005; Satoetal., 2008; Wicheretal., 2008),特异的OR决定复合体的功能。蛾类PR属于昆虫OR基因家族,与昆虫其他OR具有相同的结构特征。
蛾类的PR基因最先在烟芽夜蛾Heliothisvirescens(Kriegeretal., 2002, 2004)和家蚕(Sakuraietal., 2004; Kriegeretal., 2005)中通过触角cDNA文库分析和基因组分析的方法被鉴定。在随后几年内,研究人员通过同源克隆的方法成功地在多种蛾类,包括粘虫Mythimnaseparata、小菜蛾Plutellaxylostella和瓜绢螟Diaphaniaindica(Mitsunoetal., 2008)、多音天蚕Antheraeapolyphemus和柞蚕A.pernyi(Forstneretal., 2009)以及苹淡褐卷蛾Epiphyaspostvittana(Jordanetal., 2009)、秆野螟属Ostrinia几种蛾类(Miuraetal., 2009, 2010; Wanneretal., 2010)、棉铃虫Helicoverpaarmigera、烟青虫H.assulta(Zhangetal., 2010)和脐橙螟Amyeloistransitella(Xuetal., 2012)等蛾类中鉴定到PR基因,这表明蛾类PR基因具有序列高度保守性。此外,系统发育分析结果表明这些典型的PR基因都聚集在进化树的同一分支,形成了蛾类PR的特定分支。这也是鉴定蛾类PR基因的一个重要依据。
近年来,随着测序技术和生物信息工具的发展和应用,目前已有65种蛾类昆虫的PR基因得到了鉴定(表1)。
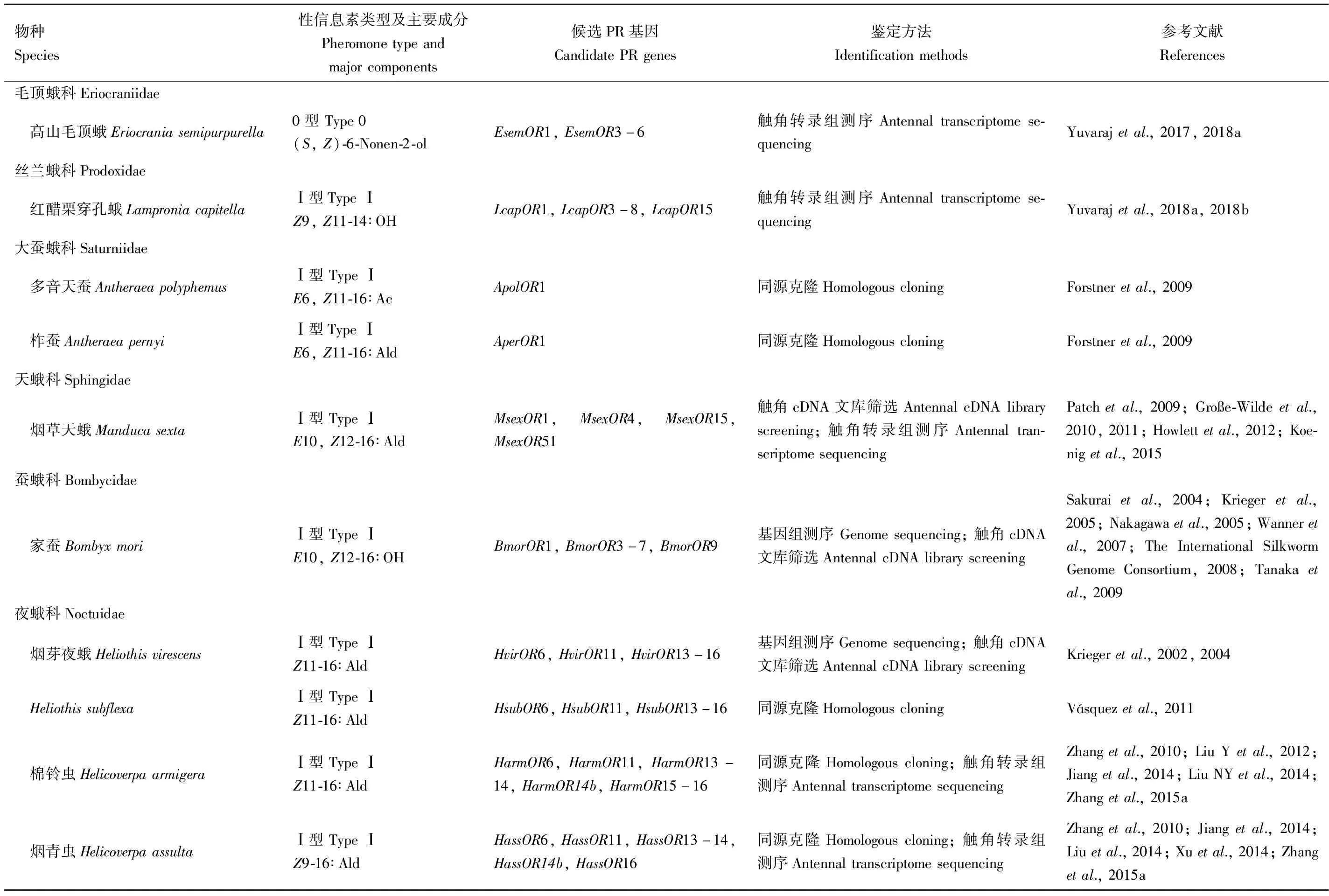
表1 已报道的65种蛾类的PR基因
2 蛾类昆虫性信息素受体基因的表达模式
通过研究PR基因的表达模式,如表达时间、表达位置,在感器中的定位等信息,我们可以推测PR的功能,为该受体的功能研究提供理论基础。研究人员一般通过RT-PCR或qRT-PCR对目标PR基因进行表达模式研究。由于性信息素是主要由雌蛾释放用于吸引同种雄蛾的化合物,而雄蛾依靠其触角上的嗅觉系统来识别性信息素,因此在大多数情况下,PR基因在雄蛾触角中特异或偏好表达(Sakuraietal., 2004; Kriegeretal., 2005; Nakagawaetal., 2005; Wanneretal., 2007)。但是,也存在一些特殊情况。例如,烟芽夜蛾的HvirOR6和家蚕的BmorOR9等在雌雄成虫触角中的表达量无明显差异(Kriegeretal., 2004; Wanneretal., 2007),而且烟芽夜蛾雌蛾触角对HvirOR6的配体Z9-14∶Ald有电生理反应(Zielonkaetal., 2018)。还有些PR基因在雌成虫触角中的表达量更高,例如红醋栗穿孔蛾Lamproniacapitella的LcapOR3(Yuvarajetal., 2018b)、烟草天蛾Manducasexta的MsexOR15(Koenigetal., 2015)、苹果蠹蛾Cydiapomonella的CpomOR21和CpomOR22(Walkeretal., 2016)、小菜蛾PxylOR8(Liu Yetal., 2018)等。这些在雌成虫触角中高表达的PR基因可能参与了雌成虫间的种内识别、种内竞争等特殊行为,避免处于高交配竞争或产卵不适宜的环境,通过减少生态资源竞争来提高自身交配率及后代的生存率(Holdcraftetal., 2016)。此外,PR基因也在成虫非触角组织中表达,这种特殊的表达模式暗示PR基因可能具有其他功能。例如,在烟芽夜蛾中识别主要性信息素成分的HvirOR13和识别次要性信息素成分的HvirOR6在成虫腹部及雌成虫产卵器中表达(Kriegeretal., 2004; Widmayeretal., 2009; Vogeletal., 2010),表明其可能参与了成虫交配时对性信息素的识别,而雌成虫可能通过识别自身释放的性信息素来反馈调节性信息素的释放量(Widmayeretal., 2009)。
除了在成虫组织中表达以外,有研究表明PR基因也在幼虫和蛹中表达。例如,家蚕BmorOR1在雄成虫羽化前4 d的蛹期开始表达,随后表达量逐渐增加(Sakuraietal., 2004),烟芽夜蛾所有PR基因在雄成虫羽化前1 d均有表达,部分基因在羽化前5 d就开始表达(Kriegeretal., 2009)。烟芽夜蛾HvirOR6和HvirOR13在1龄幼虫的头部及5龄幼虫头部和触角中均有表达,而且幼虫触角感器对性信息素成分有电生理反应(Zielonkaetal., 2016)。在之前的报道中,研究人员就发现海灰翅夜蛾Spodopteralittoralis、甜菜夜蛾S.exigua以及小菜蛾幼虫均能识别雌蛾释放的性信息素而且性信息素还能增加幼虫对食物的趋性行为(Poivetetal., 2012; Jinetal., 2015; Zhu Jetal., 2016),这表明性信息素可以作为幼虫寻找食物资源的一个重要化学线索。但是,研究人员并未在海灰翅夜蛾幼虫触角中发现PR基因的表达(Poivetetal., 2012, 2013),而甜菜夜蛾和小菜蛾幼虫的触角中是否表达PR基因目前还未见报道。
PR基因表达在触角长毛形感器内的ORN中,在通常情况下每根感器下包含2个不同的ORNs。通过原位杂交技术,研究人员可以确定表达在同一根长毛形感器中的PR基因。例如,研究发现家蚕BmorOR1和BmorOR3在邻近的ORNs中表达(Kriegeretal., 2005),烟芽夜蛾HvirOR11和HvirOR13在邻近的ORNs中表达(Kriegeretal., 2009)。
3 蛾类昆虫性信息素受体的功能研究
3.1 性信息素受体功能的研究方法
2004年,研究人员借助爪蟾卵母细胞表达系统完成了第一个蛾类PR即家蚕BmorOR1的功能研究(Sakuraietal., 2004)。由于爪蟾卵母细胞具有个体大、周期短、操作简单、结果稳定等优点,使它成为PR功能研究最常用的异源表达系统(表2)。当然,研究人员也通过其他异源表达系统研究蛾类PR的功能,主要包括HEK293(human embryonic kidney 293)细胞系(Yuvarajetal., 2018b)、CHO(Chinese hamster ovary)细胞系(Wicheretal., 2017)、果蝇“空神经元”系统(Montagnéetal., 2012)、转基因家蚕系统(Sakuraietal., 2011)、家蚕巨型囊泡(giant vesicles)系统(Hamadaetal., 2014)和草地贪夜蛾S.frugiperdaSf9细胞系(Xuetal., 2014)等(表2)。这些异源表达系统都成功地用于蛾类PR的功能研究,而且通常情况下,通过不同的表达系统得到的结果是一致的(表2),说明了这些异源表达系统用于蛾类PR的功能研究是可行的。

表2 已完成功能研究的性信息素受体的配体及研究方法
然而,在某些情况下,使用不同的表达系统鉴定的PR功能存在明显的差异。例如,苹果褐卷蛾Epiphyaspostvittana的EposOR1属于PR分支,当它在Sf9细胞系中表达时不能被性信息素成分激活,而对几种植物挥发物有电生理反应(Jordanetal., 2009)。当EposOR1在HEK293细胞系和爪蟾卵母细胞中表达时,却表现出了完全相反的功能,它能被性信息素成分激活但对前面报道的植物挥发物没有反应(Corcoran, 2011)。同样地,苹果蠹蛾的CpomOR3在转基因果蝇和HEK293细胞中表达时仅对植物挥发物梨酯有电生理反应(Bengtssonetal., 2014; Cattaneoetal., 2017),对性信息素成分没有反应,但当它在爪蟾卵母细胞中表达时,不仅能被梨酯激活,也能被性信息素所激活(Wanetal., 2019)。烟青虫HassOR13在果蝇“空神经元”表达系统和烟青虫触角ORN中的结合谱比爪蟾卵母细胞中的结合谱更窄,表现出更强的特异性(Wangetal., 2016)。Hou等(2019)和Yuvaraj等(2017)使用HEK293细胞系和爪蟾卵母细胞两种表达系统研究了高山毛顶蛾PR的功能并系统地比较了两种表达系统鉴定PR的功能差异,结果表明,在不同的表达系统中,EsemOR3和EsemOR5的选择特异性和结合特异性均存在差异;此外,EsemOR4在HEK293表达系统中对测试的性信息素成分均无电生理反应,而在爪蟾卵母细胞表达系统中特异地对主要性信息素成分有反应(Yuvarajetal., 2017; Houetal., 2019)。这种同一个PR在不同的异源表达系统中表现出明显的功能差异,可能与不同异源表达系统固有的因素有关,也可能与不同的表达系统中气味的刺激方式差异相关。另外,与昆虫体内环境相比,由于异源表达系统存在一些固有的缺陷同样会限制我们对PR功能的评估,例如,鉴定PR功能时所用的气味浓度和昆虫在自然环境中所接触的气味浓度是否相符,筛选PR配体时所使用的气味种类和数量,异源表达系统中缺少性信息素识别过程中的重要蛋白,如PBP,SNMP和ODE等。因此,当我们鉴定PR功能时,不能简单地根据体外筛选得到的配体来定义其体内功能,还需要借助体内功能研究方法,对其功能进行进一步确定。
目前,研究蛾类PR功能的体内研究方法主要包括RNA干扰(RNA interference, RNAi)技术(Linetal., 2015; Zhang QHetal., 2017),TALENs(transcription activator-like effector nucleases)基因编辑技术(Sakuraietal., 2015)以及CRISPR/Cas9(clustered regularly interspaced short palindromic repeats associated nuclease Cas9)基因编辑技术(Changetal., 2017)等。由于蛾类昆虫基因的RNAi效果差,不同基因干扰效率差异大(Tereniusetal., 2011),所以目前使用RNAi技术研究蛾类PR功能的报道较少,而CRISPR/Cas9作为一种新兴基因编辑技术,凭借其效率高、价格低、操作简单等优点,在今后PR功能研究中必将占据重要地位。但是,我们也应该注意到,如果单独运用基因敲除技术研究PR的功能会存在一定的盲目性。例如,当一个PR突变体的交配率下降时,并不能清楚地解释这个PR是如何导致交配率下降的。尽管可以通过电生理技术比较突变体和野生型对性信息素的反应差异,但仍难以解释这些差异是由这个PR直接还是间接导致的。有研究发现,位于同一根感器下的神经元之间存在侧向抑制现象并能影响昆虫嗅觉行为(Suetal., 2012; Zhang Yetal., 2019),所以,一个神经元上的受体被敲除时,很可能影响邻近神经元的功能。此外,由于受体和配体之间是组合编码的(Careyetal., 2010; de Fouchieretal., 2017),受体之间还可能存在功能补偿作用。因此,为了更全面地研究和理解PR的功能,需要将体外的功能鉴定方法和体内的研究方法充分地结合起来。一般情况下,体外功能研究方法周期较短,操作相对简单,所以我们可以先通过体外异源表达系统对目标PR的配体进行大范围地筛选,明确其候选配体,推测其在昆虫体内的功能。然后在昆虫体内将目标PR基因进行敲除,并针对其配体设计试验验证该PR的体内功能。例如,Liu Y等(2013)通过爪蟾卵母细胞表达系统确定了棉铃虫HarmOR16是一种性信息素拮抗剂Z11-16∶OH的受体,但是并不清楚它分布在哪种类型的长毛形感器及其生态学作用。随后,Chang等(2017)通过CRISPR/Cas9技术将HarmOR16在棉铃虫中进行敲除,并借助电生理及行为学实验,明确了该受体与棉铃虫选择最优的交配时间有关。
3.2 蛾类典型性信息素受体的功能研究
通常情况下,蛾类典型的PR基因比较保守,在进化树中聚集于同一分支,这也是蛾类典型PR基因鉴定的一个重要标准。表2总结了截至2019年12月,已经研究功能的蛾类PR的配体及研究方法等信息。
根据目前已研究功能的典型PR对性信息素的结合特异性,将其分为以下几类:(1)结合谱特异的PR。这类PR通常是主要性信息素的受体。例如,家蚕主要性信息素成分E10,Z12-16∶OH的受体BmorOR1(Sakuraietal., 2004),红醋栗穿孔蛾主要性信息素成分Z9,Z11-14∶OH的受体LcapOR8(Yuvarajetal., 2018b)等(表2)。特异的结合谱有助于更准确地识别同种雌蛾释放的性信息素而不受其他成分的干扰。(2)结合谱宽的PR。除了结合谱特异的PR以外,大多数PR都能被多种结构类似的性信息素成分或类似物所激活,具有较宽的配体结合谱。例如,黄地老虎Agrotissegetum所有的PR(Zhang and Löfstedt, 2013),豆秆野螟Ostriniascapulalis除了未鉴定到配体的OscaOR6-8(Miuraetal., 2009, 2010)等,均能被至少两种性信息素或类似物所激活。这类PR基因可能是对性信息素改变的一种预适应,即使性信息素成分发生了轻微的变化,雄成虫同样能追踪到雌成虫(Fleischer and Krieger, 2018)。(3)未鉴定到配体的PR。除了上面两类PR以外,还有一些PR不能被测试的化合物所激活,这类PR很常见,在目前已鉴定PR功能的蛾类中普遍存在(表2)。不过,虽然目前未鉴定到这类PR对应的配体,但是并不能说明它们完全没有作用。Zhang等发现冬尺蠖蛾ObruOR1能被II类性信息素成分1,3Z,6Z,9Z-19∶Hy所激活,而其在黄地老虎中的同源基因AsegOR3同样能被这类化合物所激活(Zhang DDetal., 2016)。系统发育分析结果表明,ObruOR1和AsegOR3与烟芽夜蛾HvirOR11,棉铃虫HarmOR11,烟青虫HassOR11以及甜菜夜蛾SexiOR11聚集在进化树的同一分支,而这些OR11在功能研究中均未鉴定到对应的配体,这说明这些OR11很可能同样识别这一类化合物。但是这些化合物的来源及其对夜蛾科昆虫的生态学意义还需进一步研究。另外,Cattaneo等(2017)借助HEK293细胞系及果蝇空神经元表达系统研究苹果蠹蛾CpomOR1的功能,发现CpomOR1在两种表达系统中均不能被测试的化合物激活。但是,Garczynski等(2017)使用CRISPR/Cas9技术将该受体进行基因敲除,发现突变体雌虫产卵量及卵孵化率均显著下降,尽管尚不清楚CpomOR1的作用机理。
3.3 蛾类其他性信息素受体的功能研究
尽管科研人员已经在蛾类昆虫中鉴定了大量的典型PR,并且对其中一部分的功能进行了深入地研究,但是仍然存在一些问题没有解决。例如,在几个已进行OR鉴定和分析的物种中未鉴定到典型的PR基因,如黄野螟Heortiavitessoides(Chengetal., 2019)及松毛虫属Dendrolimus的3种松毛虫(Zhang SFetal., 2014, 2017a, 2017b)。此外,在多个已完成典型PR功能研究的物种中,未找到其主要性信息素成分的受体,如斜纹夜蛾Spodopteralitura(Zhangetal., 2015b)与海灰翅夜蛾(Montagnéetal., 2012; de Fouchieretal., 2015)的主要性信息素成分Z9,E11-14∶Ac的受体,苹果蠹蛾主要性信息素成分E8,E10-12∶OH的受体(Bengtssonetal., 2014; Cattaneoetal., 2017; Garczynsketal., 2017; Wanetal., 2019)以及苹果褐卷蛾主要性信息素成分E11-14∶Ac(Jordanetal., 2009; Corcoran, 2011)的受体等。这些问题都暗示了蛾类昆虫可能还存在其他的PR基因。
近些年,研究人员在几种蛾类昆虫中发现一些位于典型PR分支以外的PR基因,这些PR同样起着识别性信息素的作用。Bastin-Héline等(2019)通过表达量及功能筛选,找到了海灰翅夜蛾一个独立于传统PR分支的候选PR基因,通过基因敲除并借助行为、电生理等研究证明了SlitOR5为其主要性信息素的受体。Li等同样通过OR基因的表达量,找到一簇在灰茶尺蠖Ectropisgrisescens雄蛾触角中高表达的OR基因,但是这个OR基因并不在传统的PR分支中,通过体外功能研究,发现这个分支中的一个OR能特异地识别其主要性信息素成分(Li ZJetal., 2017)。Yuvaraj等(2017)通过触角转录组鉴定了一种单孔亚目蛾类高山毛顶蛾的OR基因,通过功能研究找到3个识别其性信息素的PR基因,这3个PR基因构成了一个独立的PR分支(图1)。
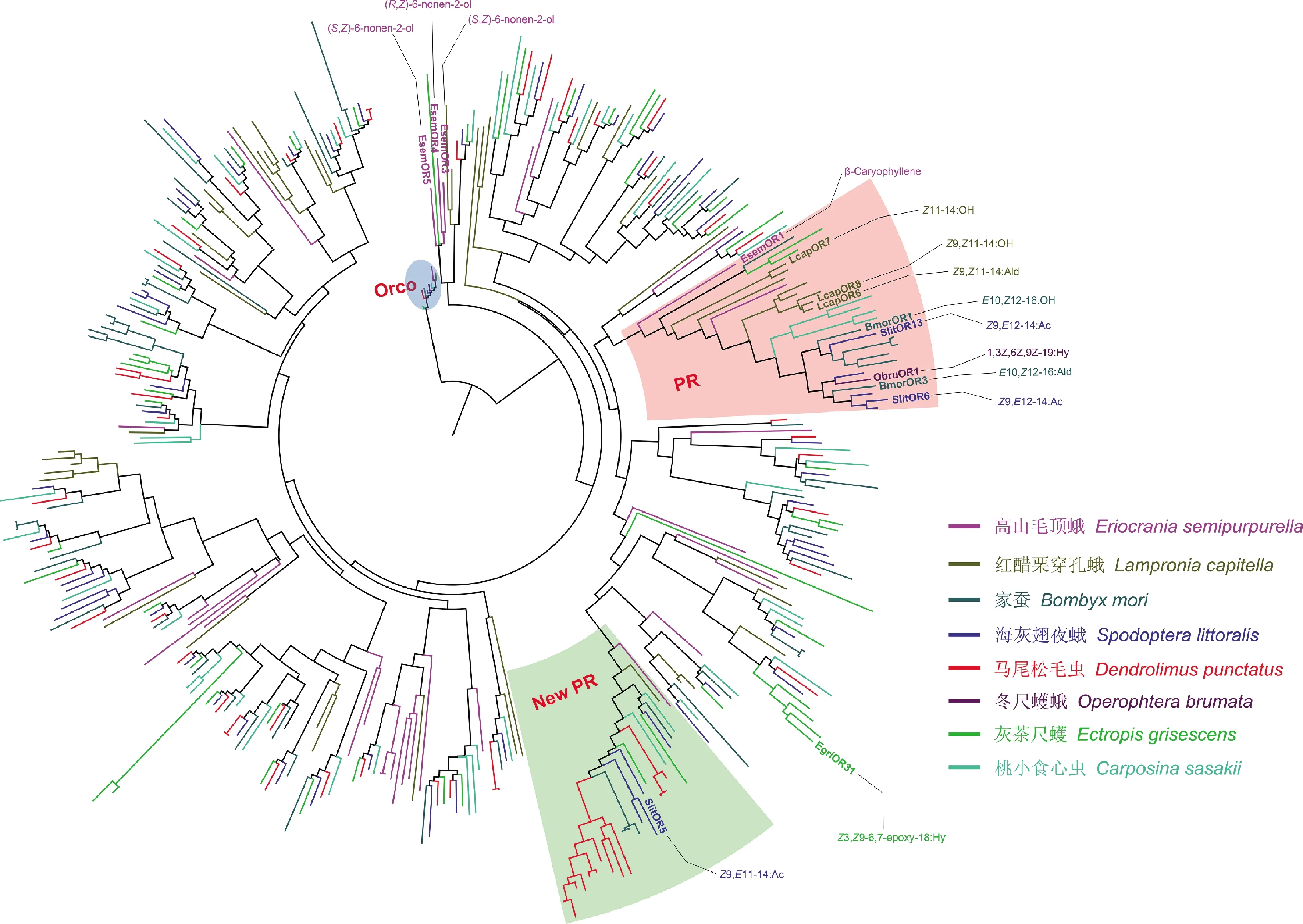
图1 蛾类气味受体的进化分析
目前人们对于蛾类非典型PR的研究报道较少,主要集中在上述海灰翅夜蛾等3个物种中。但是随着更多蛾类的PR基因得到鉴定和功能得到研究,我们对蛾类PR的认识也必将更加全面,更加深入。
4 蛾类昆虫性信息素受体的进化
雌蛾释放的性信息素作为种内异性交流的主要方式,也使种间产生了生殖隔离。通常情况下,不同种雌蛾释放的性信息素成分存在很大差异。为了能在复杂的环境中找到合适的配偶,雄蛾必须进化出一套与其高度特异的信息素相对应的PR基因。为了研究不同种蛾类PR的进化关系,我们使用RAxML v8(Stamatakis, 2014)的JTT氨基酸替换模型构建进化树。进行1 000次bootstrap统计学检测,选取7种蛾类的OR基因及冬尺蠖蛾的ObruOR1和ObruOrco进行系统发育分析(图1)。这7种蛾类包括高山毛顶蛾、红醋栗穿孔蛾、家蚕、海灰翅夜蛾、马尾松毛虫Dendrolimuspunctatus、灰茶尺蠖以及桃小食心虫Carposinasasakii。这些蛾类物种均具有独特的特征:高山毛顶蛾的性信息素为0型性信息素,红醋栗穿孔蛾是性信息素为I型的单孔亚目蛾类,家蚕为蛾类昆虫的模式昆虫,海灰翅夜蛾中发现了新的PR基因分支,马尾松毛虫不包含传统的PR基因,灰茶尺蠖的性信息素为Ⅱ型性信息素,并已鉴定到独立于传统PR分支的PR基因,桃小食心虫的性信息素类型属于其他类型,冬尺蠖蛾的性信息素为Ⅱ型性信息素,并已鉴定到位于传统PR分支上的PR基因。
从进化树我们可以发现,尽管这些蛾类的亲缘关系较远,但是它们的Orco基因仍聚在进化树的同一分支,具有高度保守性。相对来说,不同蛾类PR基因的进化关系较远。根据目前已鉴定功能的PR基因的系统发育关系,我们发现除了传统的PR分支以外,还包含3个PR分支,高山毛顶蛾的3个PR基因构成了一个感受0型性信息素成分的PR分支,海灰翅夜蛾中新发现的一个识别其主要性信息素成分的PR基因构成了一个新的PR分支以及灰茶尺蠖识别其主要性信息素的PR基因形成的一个分支(图1)。
值得注意的是,在传统的PR分支中,既包括识别I型性信息素的PR又包括识别Ⅱ型性信息素的PR,同时还包括一个识别重要植物源气味β-caryophyllene的OR(EsemOR1)。另外,性信息素为其他类型的桃小食心虫的4个候选PR也分布在此分支中,尽管目前它们的功能还没有得到鉴定。这说明识别不同类型性信息素的PR基因在进化上并没有完全分离,可能由相同的祖先基因(ancestral gene)进化而来。但奇怪的是,马尾松毛虫的性信息素的类型为I型,但是它的PR基因并不分布在传统PR分支中,这说明了马尾松毛虫PR基因的进化与其性信息素种类以及物种进化并没有直接联系。冬尺蠖蛾ObruOR1与识别I型性信息素的PR基因的进化关系,同样支持这一观点(图1,粉色分支)。
海灰翅夜蛾作为一种典型利用I型性信息素的蛾类昆虫,它识别主要性信息素的PR基因编码蛋白,SlitOR5分布在一个远离传统的PR分支的分支中(图1,绿色分支)。我们发现,这个分支包含多个马尾松毛虫的OR基因,同时还包括其他除了冬尺蠖蛾(进化树仅包括报道的两个基因)以外所有蛾类的OR基因,这说明了这个新的PR分支并不是海灰翅夜蛾所独有的,很可能与传统的PR分支一样,在OR进化过程中分化并独立出来。尽管处于这个分支中其他蛾类的OR基因的功能还未得到验证,但我们发现,对于某一种蛾类来说,它的PR基因在两个分支中的分布存在一定偏向性。例如,马尾松毛虫所有的候选PR基因都分布在此分支,而家蚕和红醋栗穿孔蛾候选PR基因则更多分布在传统PR分支中。当然,如果想明确两个PR分支的进化关系,还需要更多的实验证据支持。
灰茶尺蠖的一个识别其主要性信息素的候选PR既不属于传统的PR分支中也不属于新鉴定的PR分支(图1)。在系统发育树中,这一簇基因与新的PR分支关系更近,但是目前还不能确定它们是否属于同一分支。如果它们是属于同一分支,那么对这几个物种而言,PR基因更多存在于新的分支中,这个结果可能在一定程度上颠覆我们对蛾类PR基因进化及性信息素识别机制的认识。
另外一个PR分支由高山毛顶蛾的3个PR基因构成,这3个基因主要分布在蛾类昆虫识别植物源气味的OR基因分支中。由于高山毛顶蛾的性信息素结构更类似于植物挥发物,因此存在这样的进化关系也很好理解。此外,我们注意到不管在传统PR分支还是新的PR分支中,它们最外缘都是高山毛顶蛾的OR基因(图1),这与高山毛顶蛾在蛾类中的进化关系一致,说明了其他蛾类的PR可能是由高山毛顶蛾的OR进化而来。
对于亲缘关系较远的蛾类,它们PR基因的进化关系较远(图1),PR的功能也存在很大差异(表2)。但对于一些高度近缘的蛾类,它们的性信息素成分和PR基因都非常相似,因此它们必须借助与其近缘种同源的PR基因,通过基因表达量差异,基因功能分化等方式来区分同种雌蛾与近缘种雌蛾所释放的性信息素成分。
欧洲玉米螟Ostrinianubilalis(ECB)包含两个亚种,分别为E型和Z型,Z型的性信息素为Z11-14∶Ac(Z11)和E11-14∶Ac(E11),主要组分为Z11-14∶Ac。而亚洲玉米螟Ostriniafurnacalis(ACB)的性信息素组分为1∶1比例的Z12-14∶Ac(Z12)和E12-14∶Ac(E12)。尽管它们性信息素组分的结构非常相似,但是它们的雄成虫仅特异地识别同种雌成虫释放的性信息素,这暗示了两个近缘物种识别性信息素的过程存在差异。对其PR的功能研究结果表明,亚洲玉米螟的ACBOR3特异地识别其性信息素成分E12和Z12,而欧洲玉米螟Z型的直系同源基因ECB(Z)OR3则对同种雌蛾产生的性信息素E11有强烈反应,对亚洲玉米螟的性信息素E12和Z12的反应很弱。通过受体选择压分析,结合单点突变和功能研究,研究人员发现ECB(Z)OR3的氨基酸序列第148位的丙氨酸(A)突变导致了两个受体的功能分化(Learyetal., 2012)。与此类似,近缘物种棉铃虫和烟青虫性信息素组分均为Z11-16∶Ald和Z9-16∶Ald,但是两者的比例却完全相反,分别为97∶3和3∶97(Li RTetal., 2017)。两个物种PR的表达模式及其体外功能研究结果表明,OR13和OR14b分别作为棉铃虫和烟青虫识别其主要性信息素Z11-16∶Ald和Z9-16∶Ald的受体,它们在两个物种中的表达量也发生了变化(Liu Yetal., 2013; Yang Ketal., 2017)。此外,OR14b在两个物种中的功能也发生了分化,HassOR14b特异地识别烟青虫的主要性信息素成分Z9-16∶Ald,而HarmOR14b则主要识别棉铃虫的次要性信息素成分Z9-14∶Ald和Z9-16∶Ald。通过定点突变结合功能分析的方法,研究人员发现两个氨基酸位点的突变导致了OR14b在两个物种间的功能分化(Yang Ketal., 2017)。
这些研究表明,近缘种间直系同源PR的氨基酸位点突变导致了PR的功能发生了分化,进而引起近缘种对同种性信息素的行为差异。这种通过氨基酸突变而导致其对性信息素结合能力发生改变的PR,是与其高度特异的性信息素及物种分化是相适应的,也是驱动新物种形成的重要动力。
5 展望
在过去15年中,蛾类昆虫PR的研究已取得了诸多进展,17个科65种蛾类的PR基因得到了鉴定,30种蛾类的PR功能得到研究(表1和2),但仍存在很多问题亟需解决。在今后的PR研究中,以下方面的研究可以作为研究的重要方向:
(1)目前PR的功能研究主要集中在夜蛾科昆虫中,对其他科昆虫特别是非I型性信息素蛾类的PR功能研究较少,严重限制我们对PR基因进化的理解,在今后研究中可鉴定更多非I型性信息素蛾类的PR及其功能,增加对PR基因进化的认识。
(2)缺乏对于一些具有特殊表达模式的PR基因的功能认知,如在雌蛾触角中高表达,在成虫其他组织或幼虫、蛹中也有表达的PR,它们可能具有一些与其表达量相对应的特殊功能,通过对这些特殊PR的功能进行解析拓宽对PR功能的认识。
(3)蛾类的PR新分支均是2017年之后才开始报道,目前研究人员对这类基因的关注度较低,功能研究则更少。以后在对蛾类昆虫PR基因鉴定和功能研究时,也需要更多关注新PR分支的PR基因,特别是未鉴定到传统PR分支PR的蛾类,这些研究必将进一步促进揭示PR进化与蛾类进化的关系。
(4)PR基因在蛾类昆虫的性信息素识别过程中发挥核心作用,但是其他嗅觉相关蛋白,包括PBP,IR以及SNMP等同样发挥重要作用。目前对这些嗅觉蛋白单独的功能研究报道较多,但是对它们如何协同发挥作用的研究非常缺乏,这也限制了我们对蛾类昆虫性信息素识别分子机制的认知。今后可对PR与其他嗅觉相关蛋白,特别是PR与PBP和SNMP1的互作关系进行研究,加强对PR作用机制的理解。
(5)昆虫的PR属于OR亚家族,目前OR的三级结构的研究仅限于对Orco的结构研究,但是对PR与Orco形成异聚复合体的结构研究仍是空白,极大地限制了我们通过蛋白结构对PR配体结合的关键氨基酸位点及PR功能的预测。因此,尝试解析PR和Orco形成复合体的结构,有助于理解PR结构和功能的关系,以及PR功能分化和物种进化的关系。
(6)尽管目前很多蛾类PR的功能得到了鉴定,但是基于PR功能指导开发蛾类害虫绿色防控技术的报道却很少。在今后的研究中,通过已鉴定PR的功能设计更加高效的蛾类害虫绿色防控措施将是本领域研究的热点。首先,由于很多蛾类昆虫的性信息素成分并不稳定,在田间应用时严重影响使用效果和防控时间。因此,可以从PR的功能入手,筛选能激活相同PR且结构更加稳定的配体,从而增加行为调控剂的作用时长,保证作用效果。另外,PR对配体的识别具有一定的灵敏度,我们可以使用PR高敏感的配体替换低敏感的配体,在保证不影响防控效果的同时降低生产成本。当然,也可以通过种植转基因植物,释放转基因目标昆虫等方法对害虫的目标PR进行干扰或敲除,影响蛾类害虫的交配和种群繁衍,达到防控目的。
