沉默人乙酰转移酶样蛋白基因对结直肠癌CL187 细胞放射敏感性的影响
2020-03-03胡乐林陶慧慧杨阳丽陶欣荣唐小龙
胡乐林 陶慧慧 杨阳丽 陶欣荣 唐小龙
1(安徽理工大学医学院第一临床学院 淮南232001)
2(安徽理工大学第一附属医院肿瘤放疗科 淮南232001)
3(安徽理工大学医学院生物化学与细胞生物学教研室 淮南232001)
结直肠癌是我国最常见的消化系统恶性肿瘤。近年来,由于肥胖、饮食结构和生活方式的改变,我国结直肠癌的发病率持续上升。2015 年,我国新发病例376 300例,居中国人群恶性肿瘤发病的第5 位;死亡病例191 000 例,居中国人群恶性肿瘤死因的第5位[1]。不同于欧美国家,我国直肠癌的发病率高于结肠癌,且大多数直肠癌患者就诊时已处于局部进展期[2]。手术是局部进展期直肠癌的重要治疗方式,但是单纯的手术治疗疗效较差,且局部复发率较高[3]。
新辅助放化疗联合全直肠系膜切除术,是目前国际推荐的进展期直肠癌的标准治疗方案。新辅助放化疗可以提高直肠癌的手术切除率、肿瘤局控率及保肛率[4],但放疗抵抗严重制约了新辅助治疗的临床疗效,新辅助放化疗后只有约20%的患者达到病理完全缓解(Pathologic complete response,PCR)[5]。目前,国内外尚缺乏能够准确评判结直肠癌放射敏感性的分子靶标,因此,明确结直肠癌放疗抵抗的机制和探索结直肠癌放疗敏感性的增敏因素,对提高放疗患者的临床疗效至关重要。
有研究表明,多种组蛋白乙酰转移酶在放射敏感性调节中起重要作用。近年来还发现,多种组蛋白乙酰化酶抑制剂,如Anacardic acid、Garcinol、Curcumin等能增加多种肿瘤细胞的放疗敏 感 性[6-8]。 hALP (human acetytransferase-like protein)也称为NAT10 (N-Acetyltransferase 10),是一种乙酰转移酶,在染色体有丝分裂期hALP与hsSUN1(Sad1 unc-84 domain protein 1)共同作用促进组蛋白H2B 的Lys15 和H4 的Lys8、Lys12、Lys16 乙酰化[9]。hALP 是否参与放射敏感性的调节尚不明确。本研究通过转染SiRNA(Small interfering RNA)片段,观察沉默hALP基因后结直肠癌细胞放射敏感性的改变,为探讨结直肠癌放射抵抗机制提供理论基础。
1 材料与方法
1.1 数据库
在Oncomine microarray 数据库(http://www.oncomine.org)检测hALP mRNA 在配对结直肠癌标本中的表达。以“hALP 或NAT10”作为关键词,使用“Cancervs.Normal analysis”为初级过滤条件,选择“Colorectal cancervs.Normal analysis”作为分析类型。采用Studentt检验计算各组p值。Oncomine网站提供标准化、规范化的统计学计算。
1.2 细胞系和主要试剂
CL187细胞购自美国ATCC细胞库;DMEM培养液和胎牛血清购自杭州四季青生物技术有限公司;hALP SiRNA 和阴性对照SiRNA (Negative control,NC)由上海吉玛制药技术有限公司设计并化学合成;脂质体转染试剂Lipofectamine2000购自美国Life 公司;AnnexinV/PI (Propidium iodide)双染凋亡试剂盒购自北京庄盟生物科技有限公司;hALP 抗体、P53 抗体、BAX(BCL2 associated X)抗体、β-actin 抗体购自武汉三鹰生物科技有限公司;CCK8试剂盒购自合肥睿捷生物技术有限公司;ECL 显色试剂盒购自上海碧云天生物科技有限公司。
1.3 SiRNA序列
hALPSiRNA序列:5'-CAGCACCACUGCUGAGAAUAAGA-3'; NC 序列:5'-UUCUCCGAACGUGUCACGUTT-3'由上海吉玛生物技术有限公司合成。
1.4 细胞培养和照射条件
CL187 细胞生长在含10%胎牛血清的DMEM培养液中,在37oC、5%CO2培养箱中培养。细胞传代选择0.25%EDTA的胰酶消化。照射方法为采用瓦里安Triology 直线加速器6 MV X 射线照射,源皮距100 cm,照射野20 cm×20 cm,剂量率300 cGy/min。
1.5 转染SiRNA寡核苷酸
25 cm2培养瓶中接种0.8×106个细胞,培养24 h后使细胞能够达到80%的汇合。取退火复性的SiRNA 200 nmol/L,加无血清的DMEM 培养基200 μL,混匀。取脂质体14.25 μL,加入200 μL无血清的DMEM培养基混匀。在5 min中之内将以上两种溶液混合,室温静置20 min。加入待转染的细胞,在37oC、5%CO2培养箱中培养。转染6 h后,换上含有10% 胎牛血清(Fetal bovine serum,FCS) 的DMEM 培养基。转染48 h 后细胞接受射线照射,剂量8 Gy,24 h 后收集细胞利用蛋白印迹法检测蛋白的表达情况。
1.6 克隆存活分析实验
取合适数量的细胞接种于35 mm 的细胞培养皿中(以最终每个培养皿中有100 个大于50 个细胞的克隆为宜),在37oC、5%CO2培养箱中培养。细胞贴壁后给予射线照射,剂量分别为0 Gy、2 Gy、4 Gy、6 Gy、8 Gy。照射结束后,继续培养12~14 d。0.02%的亚甲蓝染色。计数每个培养皿中含50 个细胞以上的克隆数。克隆形成率=(克隆数/细胞接种数)×100%;细胞存活分数=实验组克隆形成率/对照组克隆形成率。单机多靶模型拟合剂量存活曲线。相对生物学效应=对照组细胞存活分数/实验组细胞存活分数。
1.7 细胞存活率检测
CL187细胞接受递增剂量的射线照射后,接种于96 孔板,每孔接10 000 个细胞,设3 个复孔。96 孔板置于培养箱中继续培养24 h 后每孔加入10 μL CCK8 溶液,置于培养箱内避光孵育2 h。酶标仪于450 nm 波长处测定吸光度值。细胞存活率=(干扰组吸光度值/对照组吸光度值)×100%。
1.8 细胞凋亡分析
将贴壁细胞用胰酶消化,尽量使细胞呈单个状脱落。加入含10%FCS 的DMEM 终止胰酶的作用,并将细胞吹打散,转移到离心管中,常温1 000 r/min,离心5 min。用4oC 预冷的1×磷酸缓冲盐溶液(Phosphate buffer saline,PBS)洗3 遍。每次洗之前,先用残留的PBS 将细胞沉淀弹开,防止细胞结块。用1×PBS重新悬浮细胞,并计数。取5×105细胞悬浮液,1 000 r/min,离心5 min后弃上清,加入500 μL 的1×Binding buffer 重悬细胞。加入5 μL 的Annexin V-FITC,轻轻混匀。在室温下,避光反应15 min。上机前1 min 再加入2 μL,PI,在1 h内进行流式细胞仪检测。
1.9 Western blotting检测
放 射 免 疫 沉 淀 测 定 法 (Radioimmunoprecipitation assay,RIPA)裂解液裂解细胞,提取细胞总蛋白。测蛋白浓度后,加SDSPAGE 蛋白上样缓冲液,95oC 变性5 min 后上样。按照每块胶15 mA 的电流进行恒流电泳,转至PVDF 膜,在5%脱脂奶粉的TBS 封闭液中室温封闭1 h,一抗4oC 孵育过夜。TBS-T 洗膜,每次5 min,洗4 次。二抗按比例稀释(山羊抗兔HRP 抗体:1︰5 000;山羊抗鼠HRP 抗体:1︰4 000),室温孵育1 h。弃掉二抗,用TBS-T洗膜,每次5 min,洗4 次。按说明书用ECL 试剂发光,X 光片压片、显影、定影,清水漂洗后自然晾干,观察实验结果并做好相关记录。
1.10 数据统计分析
所有实验至少重复3 次,应用SPSS 17.0 软件进行统计学分析。实验数值用xˉ±s表示,统计分析采用两样本独立t检验比较两组间的差异,双侧p<0.05认为统计学差异有意义。使用GraphPad Prism软件进行数据作图。
2 结果
2.1 临床数据库挖掘hALP在各种肿瘤组织中的表达情况
2.1.1 Oncomine分析hALP在肿瘤中的表达概况
利用Oncomine 数据库在数据集中查询hALP在肿瘤中的表达概况。红色方块表示hALP 过表达,蓝色方块表示hALP 低表达。表达水平基于基因的百分等级。根据p<0.000 1,表达异常倍数大于2倍的限定条件,选择正常组织与肿瘤组织的对比数据,结果如图1所示(图中左向箭头为蓝色方向,右向箭头为红色方向)。Oncomine数据库中共收集了360 个不同类型的研究结果,关于hALP 表达有统计学差异的研究结果有10 个,其中8 个研究显示hALP在结直肠肿瘤(包括结直肠癌和结直肠腺瘤)中高表达,1 个研究显示hALP 在乳腺癌中高表达,1个研究显示hALP在白血病中高表达。
2.1.2 Oncomine 分析hALP 在结直肠肿瘤组织表达水平
在Oncomine 数据库的10 个数据集共收集了hALP表达有统计学差异的研究结果21个,对研究结果进行Meta 分析(见图2),明确结直肠肿瘤(包括结直肠癌和结直肠腺瘤)临床标本hALP mRNA 的表达水平。红色方块表示结直肠肿瘤临床标本hALP mRNA 的表达比正常结肠组织标本高,红色强度越强提示hALP mRNA 表达水平越高。研究结果显示,中位秩=666,p=7.40×10−9,结直肠肿瘤癌组织hALP mRNA表达水平显著高于正常结肠组织标本。右边的表格描述的这些数据集的名称和每个数据集对应的患者样本数。Oncomine分析显示hALP在结直肠肿瘤组织表达水平显著高于正常结直肠组织标本。
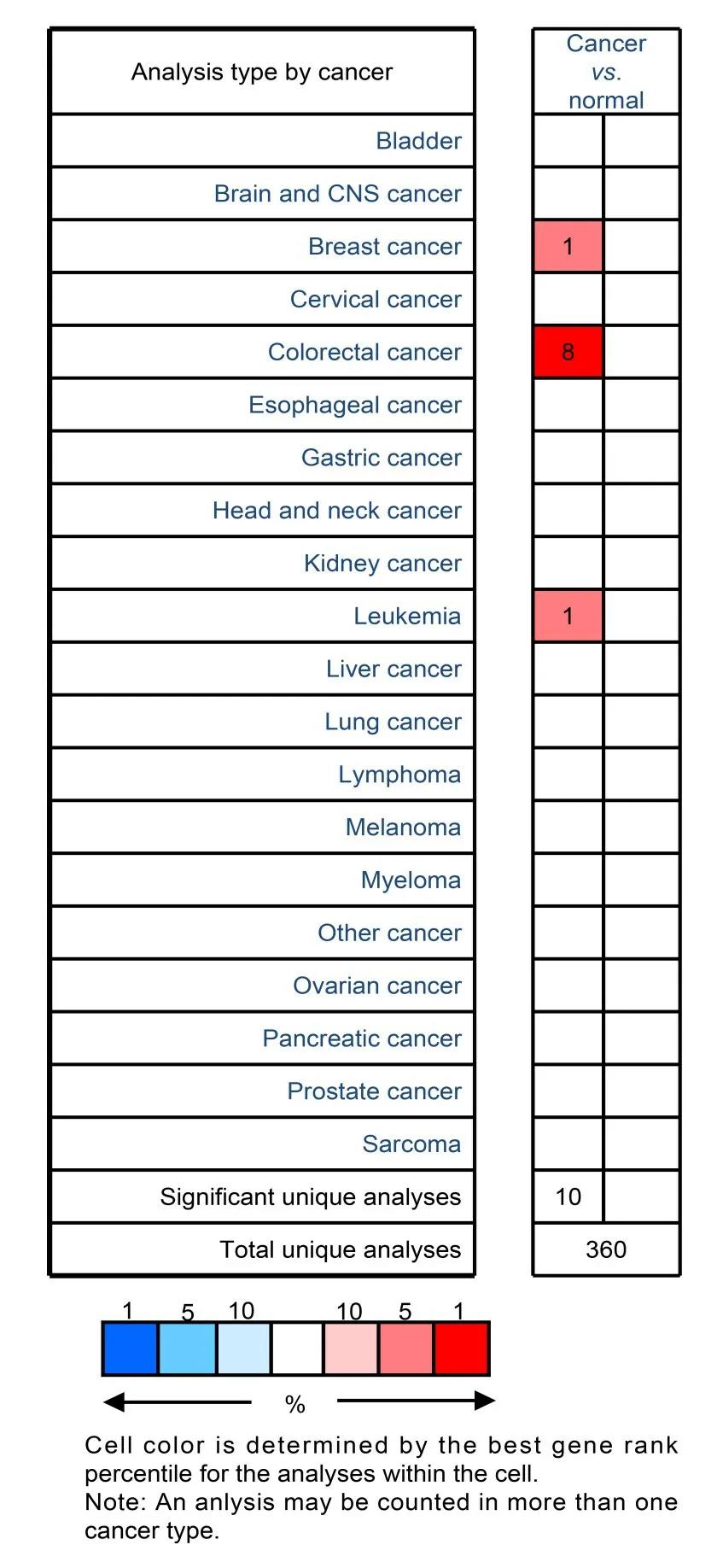
图1 Oncomine数据库分析hALP在肿瘤中的表达(彩色见网络版)Fig.1 Analysis of hALP expression in tumors using Oncomine database(color online)
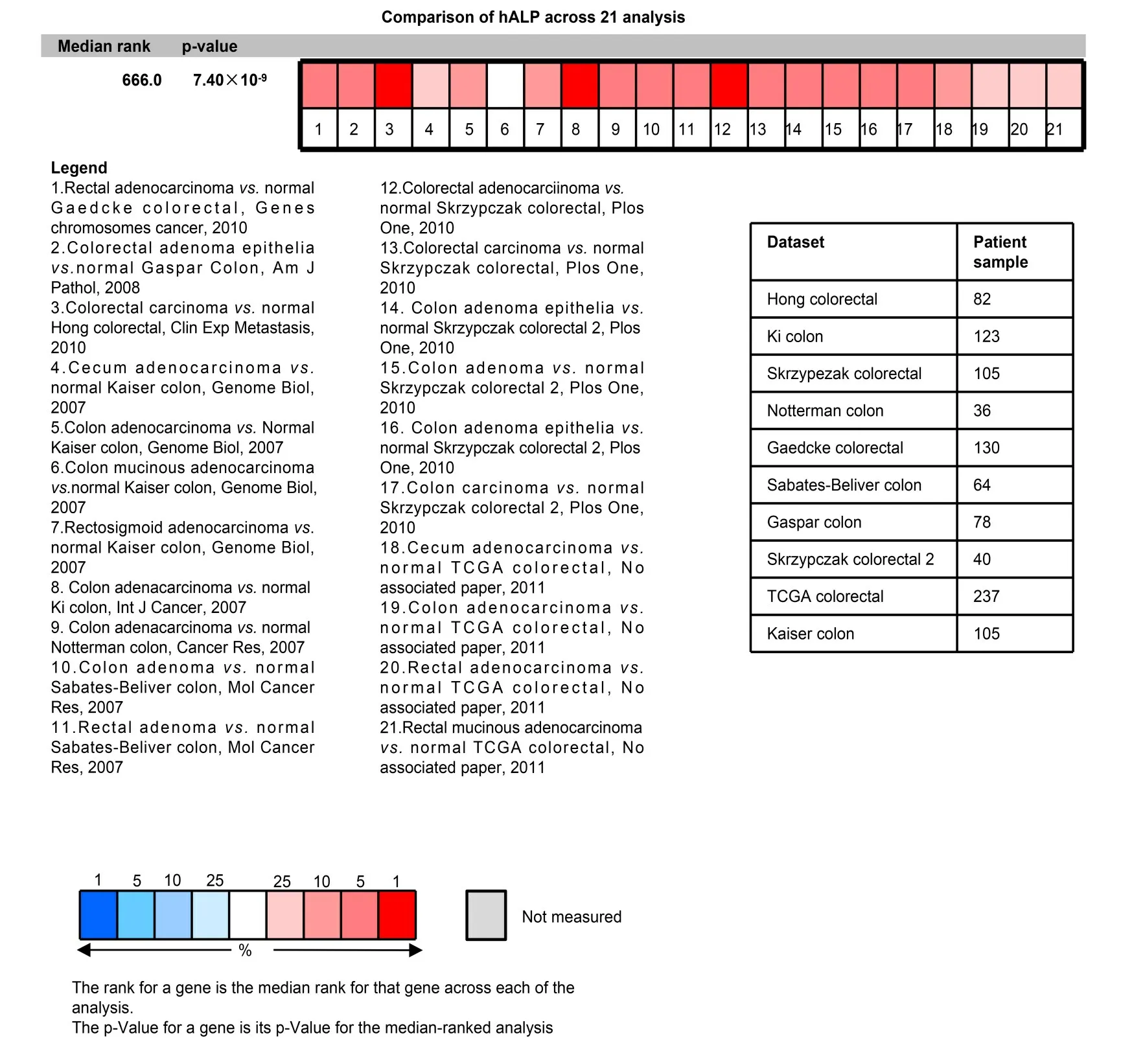
图2 Oncomine数据库分析结直肠肿瘤组织中hALP表达水平(彩色见网络版)Fig.2 Analysis of hALP expression in colorectal tumor sample using Oncomine database(color online)
2.2 结直肠癌CL187细胞沉默hALP基因后放射敏感性
CL187 细胞转染hALPSiRNA 或阴性对照SiRNA 后48 h,Western blotting 检测hALP 蛋白的表达情况。结果显示hALP SiRNA 靶向沉默hALP基因(图3(a)、(b))。沉默hALP基因后取合适数量的CL187细胞接种于35 mm的细胞培养皿中,并给予细胞射线照射,剂量分别为0 Gy、2 Gy、4 Gy、6 Gy、8 Gy。单机多靶模型拟合剂量存活曲线(图3(c))。计算RBE,即RBE=SF2(对照组)/SF2(沉默hALP基因组)。对照组,沉默hALP基因组的SF2分别是0.63、0.5。RBE为1.26。沉默hALP基因组结直肠癌CL187细胞放射敏感性增加。
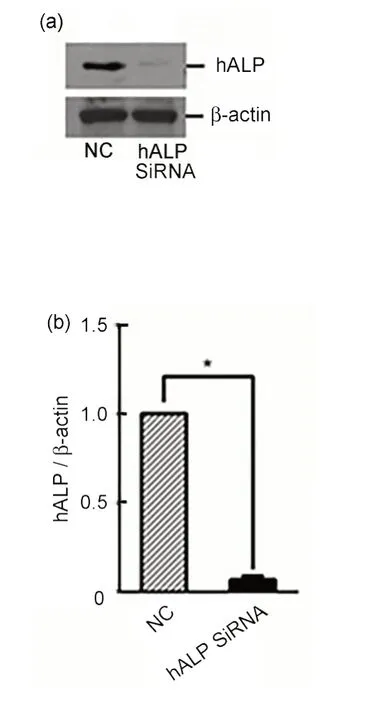
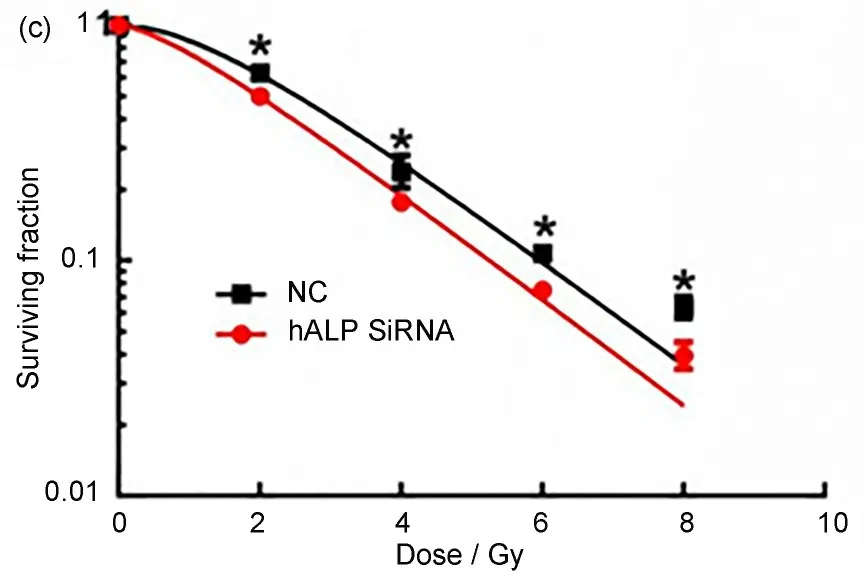
图3 (a)Western blotting检测hALP蛋白的表达情况;(b)Photoshop软件分析hALP和β-actin条带的灰度值;(c)克隆增殖实验计算细胞的存活分数,*与对照组比较,p<0.05Fig.3 (a)Western blotting was performed to assess the expression of hALP 48 h after transfection;(b)the expression of hALP was quantified using Photoshop software;(c)cell survival fraction was calculated by clonal proliferation assay,*compared with the control group,p<0.05
2.3 结直肠癌CL187 沉默hALP基因后接受射线照射细胞死亡比例
结直肠癌CL187 沉默hALP基因后,接受射线照射,剂量分别为0 Gy、2 Gy、4 Gy、6 Gy、8 Gy,照射后接种于96孔板继续培养24 h,利用CCK8试剂盒检测CL187 细胞的存活率,结果显示(图4)沉默hALP基因组CL187细胞接受递增剂量射线照射后细胞死亡比例较对照组显著增加(p<0.05),具体见图4。
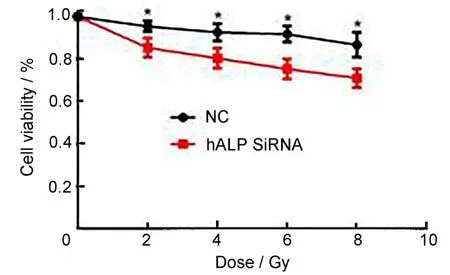
图4 CCK8试剂盒检测CL187细胞的存活率*与对照组相比,p<0.05Fig.4 CCK8 kit was used to detect the cell viability of CL187 cells,*compared with the control group,p<0.05
2.4 结直肠癌CL187沉默hALP基因后接受射线照射细胞凋亡比例
CL187 细胞沉默hALP基因后接受射线照射,剂量8 Gy,照射后24 h收集细胞,Annexin V和PI双染色,流式细胞检测术检测CL187 细胞的凋亡比例。结果如图5 所示,结直肠癌CL187 沉默hALP基因后接受射线照射,细胞凋亡比例从19.53%增高到36.49%(p<0.05)。
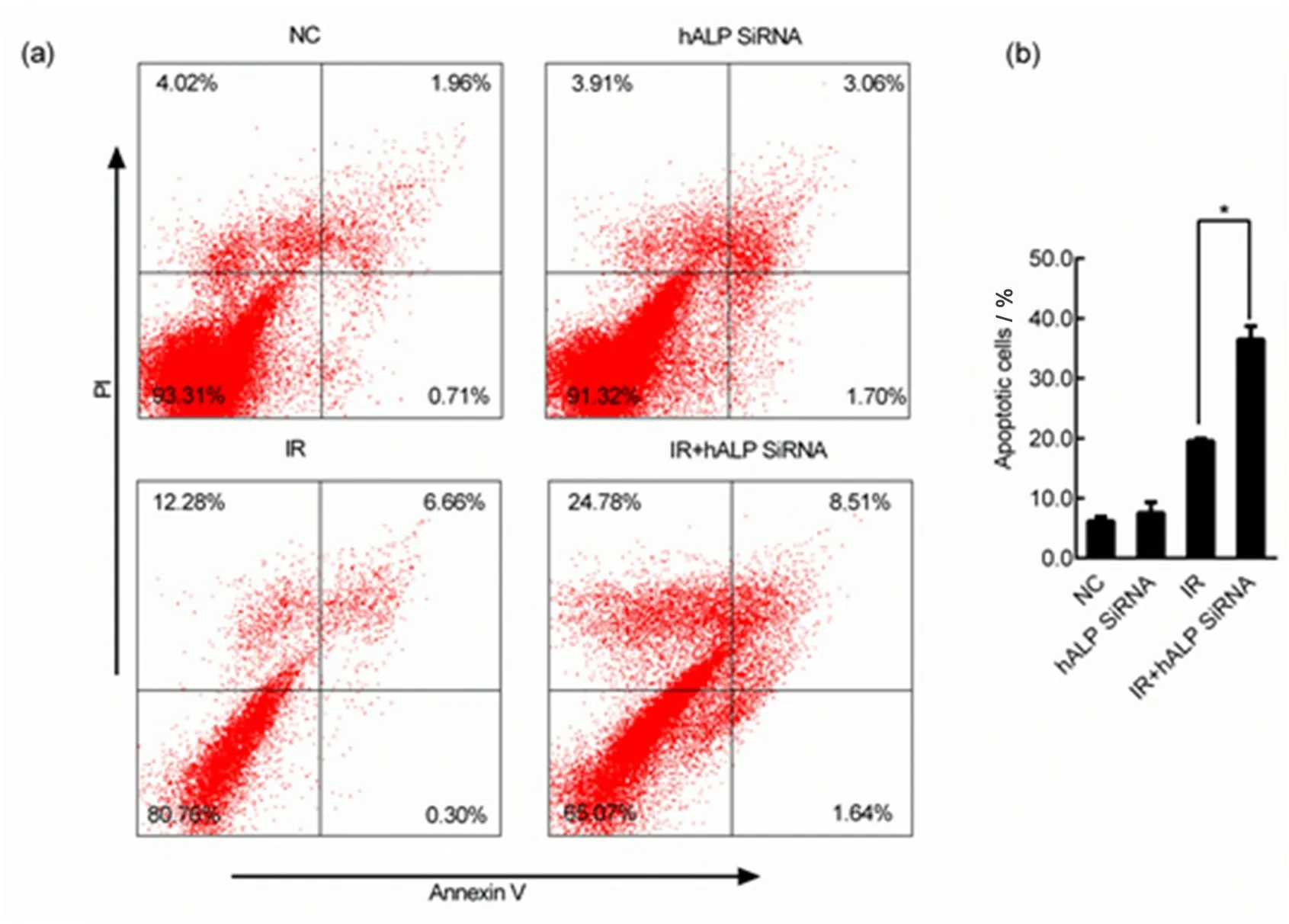
图5 (a)流式细胞分析术检测CL187细胞的凋亡情况;(b)统计学分析凋亡细胞百分比的改变*与对照组相比,p<0.05Fig.5 (a)Flow cytometry was used to detect the apoptosis of CL187 cells;(b)statistical analysis of changes in percentage of apoptotic cells,*compared with the control group,p<0.05
2.5 射线促进结直肠癌细胞hALP、P53、BAX 的表达
CL187 细胞接受0 Gy、2 Gy、4 Gy、8 Gy 递增剂量的射线照射后,置于培养箱继续培养24 h,利用Western blotting 技术检测hALP、P53、BAX蛋白的表达情况。结果显示,随着吸收剂量的增加,CL187 细胞内hALP、P53、BAX 蛋白的表达水平增高(图6)。
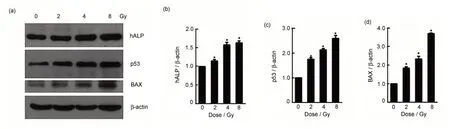
图6 (a)Western blotting检测hALP、P53、BAX和β-actin的蛋白表达;(b~d)Photoshop软件分析hALP、P53、BAX和β-actin条带的灰度值,以β-actin灰度值为对照,统计学分析hALP(b)、P53(c)和BAX(d)与β-actin灰度值的比,所有数值用x±s表示;*与对照组相比,p<0.05Fig.6 (a)The protein expressions of hALP,P53,BAX and β-actin were determined using western blotting,(b~d)expression of hALP(b),P53(c),and BAX(d)was quantified using Photoshop software.The data are normalized to the β-actin and presented as x±s,*compared with the control group,p<0.05
2.6 沉默hALP 基因后CL187 细胞接受射线照射,P53、BAX表达水平
CL187 细胞转染200 nmol/L 的hALP SiRNA 沉默hALP基因后,接受射线照射,剂量8 Gy,照射后细胞置于培养箱继续培养24 h,Western blotting检测hALP、P53、BAX 蛋白的表达情况。结果显示射线照射后P53、BAX 表达水平增高,沉默hALP 基因后射线引起P53、BAX 表达水平进一步增高(图7)。
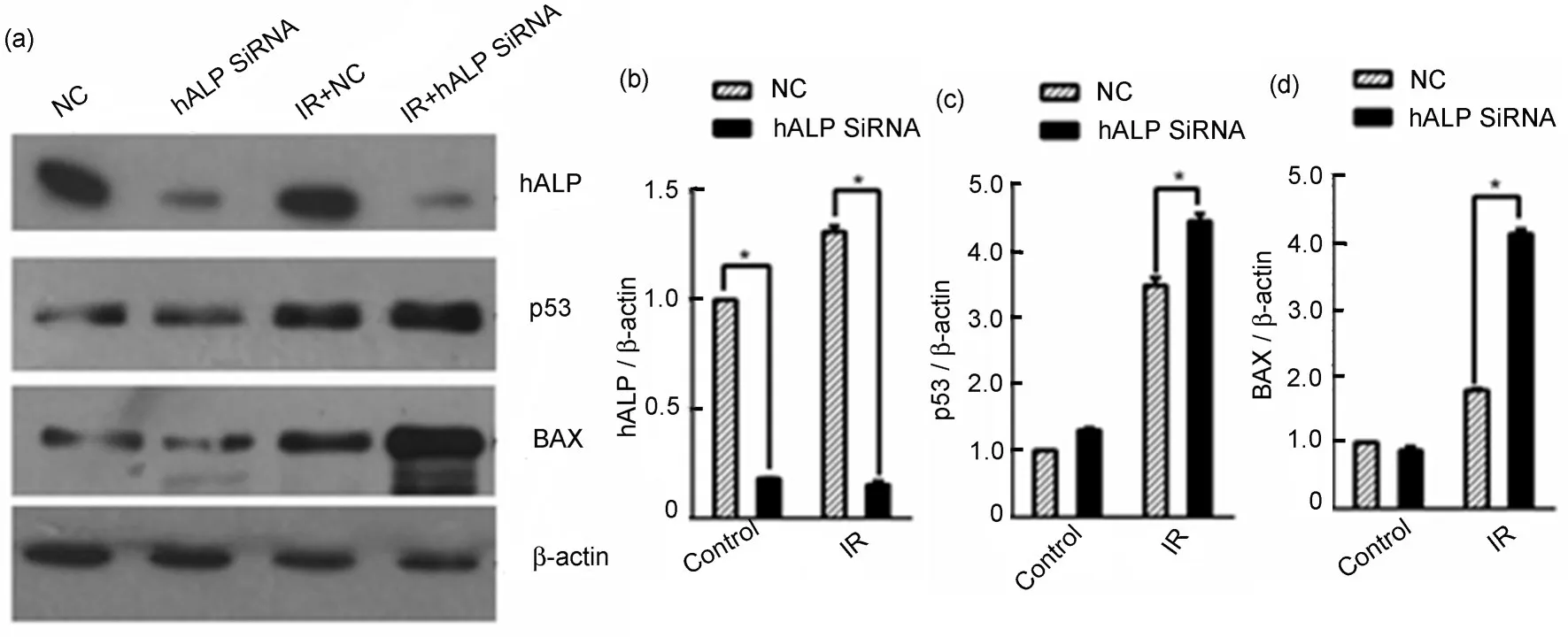
图7 (a)Western blotting检测hALP、P53、BAX和β-actin的蛋白表达;(b~d)Photoshop软件分析hALP(b)、P53(c)、BAX(d)和β-actin条带的灰度值;*与对照组相比,p<0.05Fig.7 (a)Western blotting was used to detect the protein expressions of hALP,P53,BAX and β-actin;(b~d)the expression of hALP(b),P53(c),and BAX(d)was quantified using Photoshop software;*compared with the control group,p<0.05
3 讨论
hALP基因位于11 号染色体(11p13),全长2.5 kb,编码834 个氨基酸,分子量94 kD。hALP蛋白是乙酰基转移酶家族成员之一。hALP蛋白包含N 端乙酰转移酶功能区、ATP酶功能区和C端赖氨酸富集区3 个保守结构域[10]。在细胞有丝分裂间期hALP 定位于核仁,在细胞分裂末期hALP 定位于中间体[11]。研究表明hALP的表达水平和亚细胞定位与多种肿瘤的发展、转移以及预后密切相关[12-15]。Shen 等[11]运用免疫组化和组织芯片技术对155例肉瘤,28例软组织良性肿瘤或软组织肿瘤样病变组织进行研究,发现hALP在多种软组织肉瘤中高表达,并且hALP的表达水平与肿瘤的组织分级密切相关。Zhang 等[12]对17 例肝癌患者的新鲜肝癌组织和癌旁组织进行Real-time PCR 分析,并对186例肝癌患者的肝癌组织和癌旁组织石蜡标本进行免疫组化分析,发现在肝癌组织中hALP的表达水平高于癌旁组织,hALP是肝癌预后不良的标志。Ma 等[14]使用Remodelin(hALP 的抑制剂)以及基因干扰技术降低hALP的表达,肝癌细胞的转移力和侵袭力都降低。韩斐等[16]应用免疫组化技术对80 例鼻腔鼻窦黏膜黑色素瘤患者的石蜡标本进行免疫组化分析,发现hALP的表达与肿瘤淋巴结转移有关。梁佩淇等[17]应用实时荧光定量PCR技术对48例急性髓细胞白血病患者及20例非恶性血液疾病患者或健康人的骨髓单个核细胞进行检测,发现急性髓细胞白血病患者hALP mRNA表达水平高于对照组,并且hALP mRNA的高表达与急性髓细胞白血病预后不良密切相关。Zhang等[15]对结直肠癌组织进行免疫组化分析时发现,在正常结直肠组织中hALP定位于核仁,在结直肠癌组织中hALP向胞浆和细胞膜等亚细胞器重新分布,hALP的亚细胞定位与结直肠癌的预后密切相关。尚不明确hALP 是否参与结直肠癌的发生发展,为探讨hALP 在结直肠癌的发生发展中的作用,我们通过对临床数据库挖掘,发现hALP在结直肠癌组织标本中表达量显著高于正常结直肠组织标本(图1、2),提示hALP 在结直肠癌的发生发展中起重要作用。hALP是否参与结直肠癌细胞的放射敏感性调控尚不明确。
SiRNA 是一种小的核苷酸片段,能通过碱基互补识别特异的靶mRNA,并与之结合并引起靶mRNA 的降解,从而抑制靶基因表达[18]。本研究利用hALP 的特异SiRNA 序列干扰hALP 的表达,观察其对细胞放射敏感性的影响。克隆形成实验发现,CL187 细胞沉默hALP基因组克隆形成率较其对照组明显降低(图3),表明抑制细胞内hALP蛋白的表达后,对射线照射诱导结直肠癌细胞克隆形成率下降有促进作用。CCK8试剂盒检测细胞存活率分析发现,CL187 细胞沉默hALP基因组接受递增剂量射线照射后细胞死亡比例较对照组显著增加(图4),这与图3 的结果一致,证明沉默hALP基因增加结直肠癌CL187细胞的放射敏感性。
肿瘤细胞的放射敏感性受多种因素的影响,细胞凋亡是影响肿瘤细胞放射敏感性的重要因素[19]。为探讨沉默hALP基因是否通过影响结直肠癌CL187 细胞凋亡的比例而影响肿瘤的放射敏感性,本研究利用流式细胞分析术检测发现,hALP沉默组CL187 细胞接受射线照射引起的细胞凋亡比例增加(图5),提示hALP基因影响结直肠癌CL187 细胞凋亡参与肿瘤的放射敏感性的调控。P53 和BAX 蛋白在细胞凋亡中起重要的调控作用[20]。hALP是否通过P53、BAX调控细胞凋亡参与了结直肠癌细胞的放射敏感性的调控尚不明确。本研究利用Western blotting 证实,随着剂量的增加,hALP 和凋亡相关蛋白P53、BAX 表达水平增高(图6),提示hALP 极有可能通过影响P53、BAX 的表达,参与了结直肠癌细胞的放射敏感性的调控。为进一步明确hALP 是否通过P53、BAX调控细胞凋亡参与了结直肠癌细胞的放射敏感性的调控,CL187 细胞沉默hALP基因后接受射线照射(8 Gy),Western blotting 检 测hALP、P53、BAX 蛋白的表达情况。结果显示射线照射后P53、BAX 表达水平增高,沉默hALP基因后射线引起P53、BAX表达水平进一步增高(图7)。这些研究表明,P53、BAX 在hALP 对结直肠癌细胞的放射敏感性的调控中起重要作用。
综上所述,我们的研究结果表明,hALP 可能通过P53、BAX调控细胞凋亡参与结直肠癌细胞的放射敏感性的调控;但hALP对结直肠癌放射抗拒调控网络中的作用及其具体分子机制仍不清楚,需要进一步研究。研究结直肠癌放疗敏感性的调节机制,有助于阐明hALP在结直肠癌放射敏感性中的地位,有利于在hALP差异表达的结直肠癌患者中筛选放射治疗的潜在获益人群。
