硫辛酸对草鱼脂肪细胞脂质含量及脂代谢相关基因表达的影响
2020-01-15黄陈翠黄小城边晨晨
黄陈翠,孙 健,吉 红,黄小城,边晨晨
(西北农林科技大学动物科技学院水产动物营养与饲料研究室,陕西咸阳,712100)
草鱼(Ctenopharyngodonidellus)是我国淡水养殖的四大家鱼之一,由于生长迅速,饲料来源广等优点,已成为我国主要的淡水养殖对象[1]。2018年我国草鱼的年产量超过500万吨,其产量位居淡水养殖品种之首。同时,草鱼作为食用鱼类和水生植物治理鱼类在全球广泛养殖[2]。科学家将草鱼作为潜在的草食性模式动物,展开了一系列营养、免疫、增殖放流相关的研究[3-5]。另一方面,草鱼摄食较高脂质水平日粮时,虽可快速生长并缩短上市周期,但同时会导致其腹腔脂肪过度蓄积,危害其健康,这一问题亟待解决[6,7]。
硫辛酸作为一种集抗氧化、减脂、提高免疫力等功能于一身的化合物,较多的研究报道其可以缓解高脂日粮对小鼠造成的负面影响,降低血脂、胆固醇等[8,9]。过去的研究发现,日粮中添加硫辛酸可以通过Nrf2-Keap1通路增强草鱼抗氧化能力,缓解脂质过氧化对草鱼的毒性作用[10],且适宜剂量的硫辛酸可以提高草鱼的免疫功能和抗病性[11]。另外,硫辛酸可以通过激活AMPK-ATGL通路增强脂质水解,同时调控PPARα-CPT1促进脂肪酸β-氧化供能,从而减少草鱼腹腔脂肪蓄积[12]。腹腔脂肪组织是草鱼主要蓄积脂肪的部位,而脂肪组织主要由各类脂肪细胞组成,因此可以通过减少脂肪细胞脂质含量的方法来解决脂肪组织的脂质蓄积问题。然而,硫辛酸能否调控草鱼脂肪细胞的脂代谢并减少脂质含量尚不清楚。因此,本试验在体外研究硫辛酸对草鱼成熟脂肪细胞脂代谢的影响,旨在丰富硫辛酸作为新型饲料添加剂在淡水鱼类日粮中的应用,同时也为淡水鱼类健康养殖提供新思路。
1 材料方法
1.1 试验材料
主要试剂:DMEMF12培养基(Hyclone)、PBS缓冲液、无脂肪酸牛血清白蛋白(Sigma)、I型胶原酶(Sigma)、硫辛酸(Sigma)、甘油三酯试剂盒(普利莱基因技术有限公司)、甘油试剂盒(南京建成生物工程研究所)、非酯化脂肪酸(NEFA)试剂盒(南京建成生物工程研究所)、RNAiso Plus (TaKaRa)、分析纯的氯仿、异丙醇、无水乙醇、反转录试剂盒(TaKaRa)、SYBR®Premix Ex TaqTMII (TaKaRa)等。
主要试验器材:剪刀镊子等解剖器材、200目过滤网、移液枪、酶标仪、实时定量-PCR仪等。
1.2 试验方法
1.2.1 草鱼脂肪细胞的分离
从杨凌康乐市场购买体重1 kg左右健康无病的草鱼后暂养1 d。重击头部致晕后剪断鳃弓放血,用洗洁精清洗鱼体3遍至洁净。无菌环境下分离腹腔脂肪组织,首先用75%酒精擦拭体表3遍消毒灭菌,用解剖工具缓慢剖开草鱼腹腔,分离腹腔脂肪组织。将分离的脂肪组织放入盛装无血清DMEMF12培养基的烧杯中,称重记录。用PBS洗涤脂肪组织3次后,加入等体积的0.1% I型胶原酶酶液消化脂肪组织,消化过程中连续剪碎脂肪组织并搅拌,消化时间为30 min。用有血清培养基终止消化,200目过滤网过滤消化液至50 mL离心管,700g离心10 min。上层漂浮细胞为草鱼成熟脂肪细胞,轻轻吸至另一洁净的50 mL离心管并用PBS缓冲液洗涤三次后,用2%牛血清白蛋白缓冲液重悬备用。
1.2.2 草鱼脂肪细胞的培养与处理
硫辛酸处理草鱼脂肪细胞的方法参考文献[13]中的HUFA处理脂肪细胞方法。硫辛酸处理细胞的浓度参考文献[12],试验用含有不同浓度(0、50和200 μmol/L)硫辛酸的2%牛血清白蛋白缓冲液重悬草鱼脂肪细胞,细胞浓度为6×105/mL,将细胞均匀地接种在24孔培养板中,放入28 ℃ 5% CO2培养箱中孵育6 h后取出细胞,立即放于冰盒上终止反应,4 ℃离心机700g离心10 min,用移液枪缓慢收集上层脂肪细胞至新的离心管,另外收集下层培养液。其中每个处理12个重复,每三个重复的样品混合为一个样本。
1.2.3 甘油三酯、甘油和非酯化脂肪酸检测方法
将收集的脂肪细胞用PBS洗涤两次后,加入400 μL细胞裂解液,将细胞裂解后,用于检测草鱼脂肪细胞甘油三酯含量及蛋白浓度。培养液用于检测脂肪细胞甘油和非酯化脂肪酸的释放量。
草鱼脂肪细胞甘油三酯含量和蛋白浓度,培养基甘油和非酯化脂肪酸含量的检测方法分别按照甘油三酯试剂盒(普利莱基因技术有限公司)、甘油试剂盒(南京建成生物工程研究所)、非酯化脂肪酸(NEFA)试剂盒(南京建成生物工程研究所)说明书执行。
1.2.4 RNA提取和实时定量PCR
按照文献[14]中草鱼成熟脂肪细胞总RNA的提取方法进行总RNA的提取和浓度测定,每个处理12个重复,每3个重复的样品混合为一个样本。使用反转录试剂盒(TaKaRa)反转得到cDNA备用。试验采用CFX96实时定量PCR检测系统,定量引物见表1,反应体积为20 μL,包含10 μL 2×SYBR® Premix Ex TaqTMⅡ,7.8 μL 双蒸水,1 μL cDNA 和各0.6 μL的引物。整个反应过程是95 ℃,30 s,然后95 ℃,5 s,最后60 ℃,30 s,40个循环。反应结束后,利用溶解曲线对产物的唯一性进行分析。β-actin作为内参,相对定量法比较CT2-ΔΔCt,计算基因表达量。

表1 实时定量PCR引物序列Tab.1 Primer sequences used for real-time PCR analysis of mRNA expression
注:TG:甘油三酯;NEFA:非酯化脂肪酸;ATGL:脂肪甘油三酯脂酶;HSL:激素敏感性甘油三酯脂肪酶;MGL:单酰甘油脂肪酶;PPARα:过氧化物酶体增殖物激活受体α;CPT1:肉毒碱棕榈酰基转移酶1;LPL:脂蛋白酯酶;CD36:脂肪酸转移酶;Leptin:瘦素;TNFα:肿瘤坏死因子-α;SREBP-1c:固醇调节元件结合蛋白1c;FAS:脂肪酸合成酶;ACC:乙酰辅酶A羧化酶;DGAT:甘油二酯酰基转移酶;PPARγ:过氧化物酶体增殖物激活受体γ;GyK:甘油激酶
1.3 数据分析
试验数据采用SPSS 21.0 软件分析。试验结果用平均值或者平均值±标准差表示。使用SPSS的单因素方差分析法(one-way ANOVA)和Duncan多重比较分析甘油三酯、甘油和非酯化脂肪酸的统计学差异。基因表达数据采用SPSS的独立样本T检验进行分析。差异水平为P<0.05。使用GraphPad Prism 5 (Graphpad Software,San Diego,CA)软件作图。
2 结果
2.1 硫辛酸对草鱼脂肪细胞甘油三酯含量的影响
硫辛酸对草鱼脂肪细胞甘油三酯含量的影响结果见图1。50和200 μmol/L硫辛酸孵育草鱼脂肪细胞6 h均可显著降低细胞甘油三酯含量。

图1 硫辛酸对草鱼脂肪细胞甘油三酯含量的影响Fig.1 The effect of α-lipoic acid on triglyceride content of adipocyte in C.idellus 数据以平均值±标准差表示,不同字母上标表示差异显著,下同
2.2 硫辛酸对草鱼脂肪细胞甘油和非酯化脂肪酸释放量的影响
硫辛酸对草鱼脂肪细胞甘油和非酯化脂肪酸释放量的影响结果见图2。50 μmol/L硫辛酸对草鱼脂肪细胞甘油和非酯化脂肪酸(NEFA)释放量无显著影响,200 μmol/L硫辛酸极显著增加草鱼脂肪细胞甘油和NEFA释放量。

图2 硫辛酸对草鱼脂肪细胞甘油和非酯化脂肪酸释放量的影响Fig.2 The effect of α-lipoic acid on the release of glyoerol and non-esterified fatty acid (NEFA) of adipocyte in C.idellusA:培养基甘油含量;B:培养基非酯化脂肪酸(NEFA)含量。
2.3 硫辛酸对草鱼脂肪细胞脂代谢相关基因表达的影响
2.3.1 硫辛酸对脂肪分解相关基因表达的影响
如图3A所示,200 μmol/L硫辛酸处理草鱼脂肪细胞6 h后,显著增加HSL的mRNA表达,对ATGL和MGL的mRNA表达无影响。硫辛酸对脂肪酸β-氧化相关基因的表达结果见图3B,200 μmol/L硫辛酸显著增加草鱼脂肪细胞PPARα、CPT1和CD36的mRNA表达,对LPL的mRNA表达无显著影响。另外,200 μmol/L硫辛酸处理草鱼脂肪细胞6 h对脂肪细胞因子Leptin和TNFα的mRNA表达无显著影响(图3C)。
2.3.2 硫辛酸对脂合成基因表达的影响
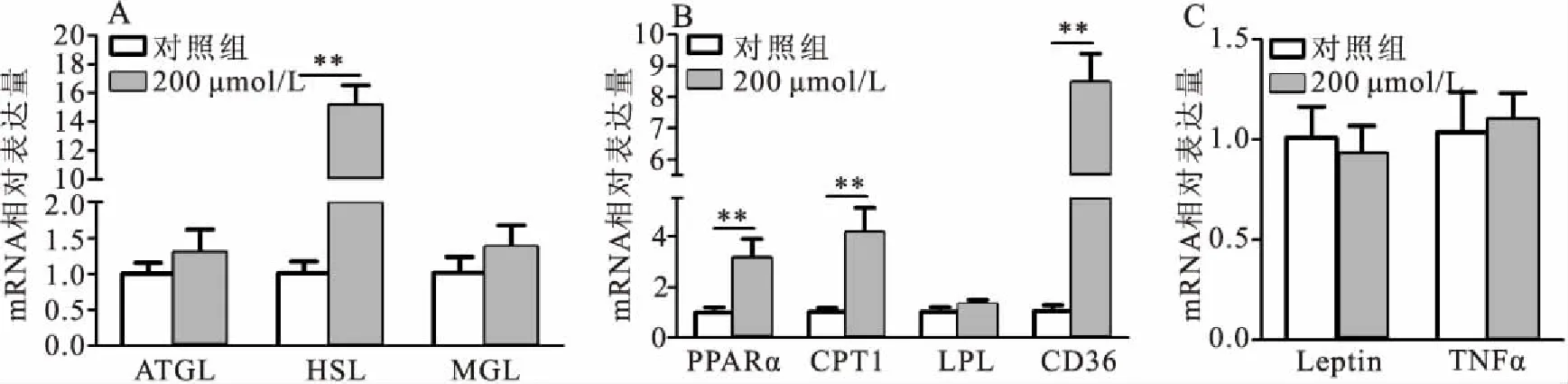
图3 硫辛酸对草鱼脂肪细胞脂肪分解相关基因表达的影响Fig.3 The effect of α-lipoic acid on mRNA expression of the genes related to lipolysis of adipocyte in C.idellus 数据以平均值±标准差表示,“**”表示差异极显著,下同

图4 硫辛酸对草鱼脂肪细胞脂合成相关基因表达的影响Fig.4 The effect of α-lipoic acid on mRNA expression of the genes related to lipogenesis of adipocyte in C.idellus
如图4所示,200 μmol/L硫辛酸极显著增加ACC和PPARγ的mRNA表达,对SREBP-1c、FAS、DGAT2和GyK的mRNA表达无显著影响。
3 讨论
硫辛酸是一种具有抗氧化、提高免疫力、调节营养代谢等功能的自然化合物,作为一种新型饲料添加剂已在陆上动物养殖中得到了较为广泛的研究。日粮中添加硫辛酸对哺乳动物的降脂功能已经得到确认,有研究指出,在高脂或高胆固醇日粮中添加硫辛酸可以降低总血脂、甘油三酯和胆固醇含量[15,16]。Jia等[17]研究发现日粮中添加硫辛酸也能显著降低肉鸡血清甘油三酯(TG)含量和增加非酯化脂肪酸(NEFA)的含量。另外,硫辛酸可以增强肉鸡肝脏和脂肪组织的脂质代谢,离体情况下增加其脂肪细胞的甘油释放量[18]。Shi等[12]研究发现硫辛酸可显著降低脂质蓄积肝细胞的甘油三酯含量。在3T3前体脂肪细胞的研究发现,硫辛酸可以抑制脂肪细胞早期分化,在分化后7天,硫辛酸显著降低了脂质含量[19]。目前尚无有关硫辛酸在鱼类脂肪细胞上的研究,本研究发现200 μmol/L硫辛酸处理草鱼脂肪细胞处理6 h后,细胞甘油三酯含量下降,且草鱼成熟脂肪细胞的甘油和NEFA的释放量显著增加。相似的结果在3T3脂肪细胞中也有发现,250 μmol/L硫辛酸处理孵育成熟的3T3脂肪细胞6 h后,处理组比对照组甘油释放量显著上升且差异最大,100 μmol/L硫辛酸处理3 h后非酯化脂肪酸显著高于对照组[20]。为了进一步探究硫辛酸对草鱼脂肪细胞脂代谢的调控作用机制,本试验对脂质代谢的相关基因表达情况进行了检测。
甘油三酯的水解是脂肪分解的重要组成部分,该过程必须通过脂肪甘油三酯脂酶(ATGL)、激素敏感性甘油三酯脂肪酶(HSL)和单酰甘油脂肪酶(MGL)三类脂肪酶的连续作用才会产生甘油和非酯化脂肪酸[21]。本研究发现200 μmol/L硫辛酸处理显著增强HSL mRNA的表达,对ATGL和MGL mRNA的表达无影响。之前的研究发现250 μmol/L硫辛酸显著上调脂质蓄积草鱼肝细胞ATGL和HSL mRNA的表达,并降低肝细胞TG含量[12]。与本研究结果不同的原因可能是由于硫辛酸对不同类型细胞的脂质水解的调控不同,即这种差异与脂肪细胞和肝细胞的不同有关。硫辛酸处理3T3脂肪细胞得到和本研究类似的结果,硫辛酸通过激活HSL磷酸化促进脂肪分解,对ATGL和G0S2的蛋白表达无影响[20]。另一方面,本研究结果表明硫辛酸可以显著增强草鱼脂肪细胞脂肪酸β-氧化相关基因(PPARα和CPT1)mRNA表达,这一结果和前人的研究结果相同[8,12]。
脂质代谢是一个动态调节的过程,表观的脂质含量是脂质分解代谢和合成代谢的综合体现。本研究发现硫辛酸可以上调草鱼脂肪细胞的脂质合成部分基因(ACC和PPARγ)mRNA的表达,对其他脂质合成基因表达无显著影响。乙酰辅酶A羧化酶(ACC)是脂肪酸合成的重要限速步骤,关系到甘油三酯(TG)的底物浓度[22]。与本研究结果相反,前人研究报道饲喂硫辛酸日粮会降低乙酰辅酶A羧化酶(ACC)和脂肪酸合成酶(FAS)的基因表达,抑制小鼠的脂质合成[23]。硫辛酸能够降低小鼠肝脏脂质含量,也是通过抑制肝脏脂肪酸合成[24]。草鱼脂肪细胞的过氧化物酶体增殖物激活受体γ(PPARγ)可以刺激甘油激酶(Gyk)的活性,而Gyk可以直接使脂质水解产生的甘油成为TG合成的直接原料-3-磷酸甘油[25]。而本研究发现硫辛酸增加草鱼成熟脂肪细胞PPARγ mRNA表达却并没有导致Gyk mRNA表达上升。实际上,PPARγ在人类的内脏脂肪细胞和肝细胞中除了调节脂质合成外,激活后可调节脂肪酸氧化酶系,促进脂质的氧化代谢[26]。本研究中PPARγ的mRNA表达上调可能是由于硫辛酸激活了PPARγ促进脂质氧化代谢的功能。
