抗衰老酶1-p53信号通路在吸入麻醉致原代海马神经元细胞凋亡中的神经保护作用
2020-01-08熊万霞赵曦宁
熊万霞,赵曦宁,王 颖
复旦大学附属中山医院麻醉科,上海 200032
吸入麻醉药作为天然分子,可广泛作用于多个位点,具有复杂的作用方式。除了常规的麻醉作用之外,吸入麻醉药还可能引起情绪、记忆和运动功能的改变,并对大脑产生持续的影响[1-3]。手术麻醉后,部分患者,尤其是老年患者,可出现学习、记忆、注意力和专注力等的障碍,而吸入麻醉药是引起认知功能障碍的可能原因之一。吸入麻醉药与手术创伤等因素能诱导围手术期氧化应激,引起细胞内活性氧自由基(reactive oxygen species,ROS)水平升高[4]。细胞内ROS及其他一系列分子相互作用可引起细胞膜电位及细胞内离子平衡的损害,最终促使细胞凋亡[5-6]。
临床常用的七氟醚(sevoflurane,Sev)、地氟醚及异氟醚能直接激活或增强γ-氨基丁酸(gamma-amino butyric acid,GABA)受体通道活性,诱导氯离子外流,导致神经元细胞膜的去极化,激活N-甲基-D-天冬氨酸(N-methyl-D-aspartic acid,NMDA)受体,使电压依赖性钙通道开放,引起钙离子内流,而细胞内钙超载可引起细胞凋亡[7-8]。有研究发现,吸入2%异氟醚6 h能上调促凋亡因子Bax水平,下调抗凋亡因子Bcl-2水平,并上调ROS水平,引起线粒体细胞色素C释放,半胱氨酸蛋白酶3(caspase-3)和caspase-9活性升高,最终诱导细胞凋亡。本课题组之前研究[9]表明,Sev联合一氧化二氮(N2O)麻醉可诱导老年大鼠海马神经元细胞的凋亡,并最终导致空间学习和记忆能力损害。
沉默信息调节因子2蛋白(silent information regulator 2,Sir2)是一组高度保守的烟酰胺腺苷酸核酸(nicotinamide adenine dinucleotide,NAD)依赖的Ⅲ类组蛋白去乙酰化酶。Sir2可通过抑制基因组转录,调节与能量代谢和促存活机制相关的蛋白表达,在能量限制引起的长寿效应中扮演重要角色[10]。目前发现的人类Sir2同源蛋白质共有7种,分别命名为抗衰老酶(sirtuin)1~7(SIRT1~SIRT7),其中以SIRT1的研究最为广泛。其作用主要依赖于其去乙酰化组蛋白和非组蛋白底物。目前,已在多种神经退行性疾病模型中发现SIRT1具有明确的神经保护作用,包括阿尔茨海默病(Alzheimer disease,AD)、肌萎缩侧索硬化(amyotrophic lateral sclerosis,ALS)、瓦勒变性(Wallerian degeneration)和多发性硬化症(multiple sclerosis)等[11-15]。本研究在以往研究的基础上进一步探讨Sev联合N2O吸入麻醉药对原代海马神经元细胞凋亡的影响及其可能机制。
1 材料与方法
1.1 实验动物及试剂 出生24 h内的SD大鼠,雌雄不限,购于上海斯莱克实验动物有限责任公司。
细胞培养液、1×PBS缓冲液、20×PBS缓冲液和DMEM购于HyClone公司;胎牛血清(fetal bovine serum,FBS)、D-hanks液、Nerobasal培养液和B27购于Gibco公司;多聚-L-赖氨酸和驴血清封闭液购于北京鼎国生物技术有限责任公司;0.25%胰蛋白酶和0.02%EDTA液购于上海吉诺生物科技有限公司;组织细胞蛋白裂解液、BCA蛋白质定量检测试剂盒、Tris-HCl、过硫酸铵(APS)、十二烷基硫酸钠(SDS)、四甲基乙二胺(TEMED)、丙烯酰胺、丽春红S、甘氨酸、Tween-20和DAPI购于上海碧云天生物技术有限公司;蛋白Marker购于Fermentas公司;脱脂奶粉购于上海光明食品厂;聚偏二氟乙烯膜(polyvinylidene fluoride,PVDF)购于Millipore公司;ECL高灵敏化学发光试剂盒购于Thermo公司;6孔细胞培养板、离心管和细胞培养皿购于Corning公司;Sev购于Baxter公司;N2O由复旦大学附属中山医院供应中心提供。SIRT1拮抗剂Sirtinol、Salermide和激动剂Resveratrol购自Sigma公司;兔抗鼠SIRT1、Bax、剪切型聚腺苷酸二磷酸核糖转移酶[cleaved poly (ADP-ribose)polymerase,,c-PARP)和β-actin抗体均购自Santa Cruz公司,兔抗鼠剪切型caspase-3(cleaved caspase-3,c-caspase-3)和磷酸化组蛋白H2A家族成员X(phosphorylated H2A histone family member X,γ-H2AX)抗体均购自Epitomics公司,兔抗鼠乙酰化p53[acetylated p53(lysine 381),Acetyl-p53]抗体购自Abcam公司。小鼠抗大鼠神经元特异性核蛋白(neuron-specific nuclear protein,NeuN)抗体购自Millipore公司。
1.2 原代海马神经元的分离培养和处理 将新生24 h内的SD大鼠用75%乙醇溶液全身消毒,断头处死,无菌条件下分层剪开头皮、颅骨,用弯镊暴露脑区视野,小心拔开大脑颞叶皮质,显露出新月状海马回。小心夹出海马组织,置于4 ℃预冷的D-hanks液中,仔细剔除微血管,并充分剪碎组织。加入同体积0.125%的胰蛋白酶溶液,瓶口用锡箔纸盖上,置于37 ℃培养箱内消化15 min,每3 min轻摇1次,过滤。加入FBS终止消化,轻轻吹打数分钟,至糊状,然后用400目网筛过滤,收集滤液。1 000 r/min室温离心5 min,弃上清液,用含10%FBS的DMEM/F12培养液重悬细胞,轻轻吹打,锥出蓝染色后快速计数。调整细胞终密度为(1~10)×105个/mL,接种于多聚赖氨酸包被的培养板中,置于37 ℃、含5%CO2的培养箱中培养。接种12 h后将培养液换成加入了2%B27和L-谷氨酰胺的Nerobasal培养液继续培养,随后每3 d半量换液1次。
采用定制的透明玻璃箱(20 cm×20 cm×10 cm),放置于37 ℃水浴箱中30 min,然后将培养2周的原代神经元置于玻璃箱中,内置气体监测仪探头,外接1.3%Sev联合50%N2O吸入麻醉+50%O2(对照组仅给予50%空氧混合气)处理2 h。或将原代神经元先分别予以100 μmol/L Resveratrol、50 μmol/L Salermide或50 μmol/L Sirtinol预处理24 h后,再吸入麻醉处理2 h。24 h后收集各组细胞,进行后续检测。
1.3 蛋白质印迹法检测SIRT1-p53信号通路相关蛋白表达 离心细胞,弃上清液,用预冷的PBS冲洗2次,吸尽上清液后,加入适量含PMSF的细胞蛋白裂解液,200 μL移液器反复吹打,冰上孵育30 min;冰上超声振荡;4 ℃,12 000×g离心30 min,将上清液移至新的EP管中。采用BCA法测定所提取蛋白的浓度后,取30 μg蛋白质,在10%十二烷基硫酸钠聚丙烯酰胺凝胶中进行电泳分离,随后在300~350 mA恒流条件下,转膜9 min,将电泳分离后的蛋白质条带转移至PVDF膜。然后,使用5%脱脂牛奶于室温封闭2 h,分别加入不同的一抗:兔抗鼠SIRT1抗体、兔抗鼠c-caspase-3抗体、兔抗鼠Bax抗体、兔抗鼠γ-H2AX抗体、兔抗鼠c-PARP抗体、兔抗鼠Acetyl-p53抗体和兔抗鼠β-actin抗体,4℃条件下孵育过夜。随后与相应的HRP标记山羊抗兔 IgG (Santa Cruz)、HRP 标记山羊抗鼠 IgG (Santa Cruz)在室温孵育1 h,使用ECL高灵敏化学发光试剂盒进行显影。对胶片进行曝光,并记录图片,然后进行灰度值分析。将各目的蛋白条带灰度值与内参蛋白(β-actin)条带灰度值的比值作为该蛋白的相对表达量。
1.4 免疫荧光法检测相关蛋白表达 将各处理组细胞在含2%甲醛的PBS中冰上固定10 min,用预冷PBS冲洗2次,每次5 min。然后,细胞在含0.1% Triton X-100和10%驴血清的PBS中进行内源性抗原封闭30 min。用PBS漂洗2次后,在每个35 mm培养皿中分别加入0.75~1 mL含兔抗鼠c-caspase-3抗体、小鼠抗大鼠NeuN抗体、兔抗鼠SIRT1抗体和兔抗鼠Acetyl-p53抗体的一抗工作液(用含10%驴血清的PBS稀释液按比例稀释NeuN 1∶500;c-caspase-3 1∶200;SIRT1 1∶100;Acetyl-p53 1∶250),室温孵育2 h。PBS洗涤3次后,每个35 mm培养皿中各加入0.75~1 mL荧光标记驴抗兔 IgG (Alexa Fluor-555)、荧光标记驴抗鼠 IgG (Alexa Fluor-488)的二抗工作液(用含10%驴血清的PBS稀释按1∶1 000比例稀释)室温孵育30 min。PBS洗涤3次后,加入稀释体积比例为1∶1 000的DAPI 染色液1 mL,室温孵育1 min。洗涤3次后,置于免疫荧光显微镜下观察染色结果并拍照。
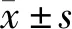
2 结 果
2.1 Sev联合N2O吸入麻醉诱导原代海马神经元细胞凋亡 蛋白质印迹法检测结果(图1A)显示,Sev联合N2O处理后原代海马神经元细胞内凋亡相关指标Bax、c-caspase-3和c-PARP表达水平显著升高(均P<0.001)。为了验证该结果,进一步采用免疫荧光实验检测c-caspase-3的表达情况,结果如图1B所示,神经元特异性蛋白NeuN在对照组和Sev联合N2O处理组均有表达,而Sev联合N2O处理后c-caspase-3的荧光染色比对照组明显增强。以上结果表明,Sev联合N2O麻醉处理可引起原代海马神经元细胞的凋亡。
2.2 Sev联合N2O吸入麻醉上调γ-H2AX蛋白水平 蛋白质印迹法检测结果(图2)显示,Sev联合N2O吸入麻醉处理后,原代海马神经元细胞中γ-H2AX的蛋白水平显著升高(P<0.001),证实吸入麻醉可引起原代海马神经元细胞的DNA断裂。
图1 Sev联合N2O吸入麻醉对原代海马神经元细胞凋亡的影响
A:蛋白质印迹法检测凋亡相关蛋白Bax、c-caspase-3和c-PARP的表达(β-actin为内参照);B:免疫荧光法检测c-caspase-3表达(NeuN为神经元特异性核蛋白).Sev+N2O:1.3%Sev联合50%N2O+50%O2吸入麻醉处理;对照组:仅给予50%O2+50%空气的混合气;***P<0.001

图2 Sev联合N2O吸入麻醉对DNA损伤标志蛋白γ-H2AX表达的影响
Sev+N2O:1.3%Sev联合50%N2O+50%O2吸入麻醉处理;对照组:仅给予50%O2+50%空气的混合气;***P<0.001
2.3 Sev联合N2O吸入麻醉上调SIRT1蛋白水平并促进p53去乙酰化 蛋白质印迹法检测结果(图3A)显示:Sev联合N2O处理可明显上调原代海马神经元细胞中的SIRT1蛋白水平(P<0.01)。此外,SIRT1作为一个去乙酰化酶,其活性作用的发挥依赖于蛋白底物的去乙酰化;而且p53蛋白是细胞凋亡的重要调节因子,也是SIRT1的主要作用底物之一。因此,本研究进一步检测了p53蛋白的乙酰化水平。结果显示Sev联合N2O处理后Acetyl-p53蛋白水平明显降低(P<0.01)。进一步使用免疫荧光实验验证Sev联合N2O吸入麻醉对SIRT1和Acetyl-p53蛋白水平的影响,结果显示Sev联合N2O处理可明显增强SIRT1的荧光染色(图3B),而Acetyl-p53的荧光强度明显减弱(图3C)。

图3 Sev联合N2O吸入麻醉对SIRT1及去乙酰化p53蛋白表达的影响
A:蛋白质印迹法检测 SIRT1和Acetyl-p53表达水平;B、C:免疫荧光实验检测SIRT1和Acetyl-p53表达.Sev+N2O:1.3%Sev联合50%N2O+50%O2吸入麻醉处理;对照组:仅给予50%O2+50%空气的混合气;**P<0.01与对照组相比
2.4 激活或抑制SIRT1通路对吸入麻醉引起原代海马神经元细胞凋亡的影响 蛋白质印迹法检测结果(图4A)表明:SIRT1拮抗剂salermide或sirtinol预处理可明显下调SIRT1蛋白的表达水平(P<0.001),并明显上调p53的乙酰化水平(P<0.001),同时抑制SIRT1后细胞凋亡相关指标Bax(P<0.001)、c-caspase-3(P<0.001)、c-PARP(P<0.001)和γ-H2AX(P<0.01)的蛋白水平进一步升高。与之相反的是,SIRT1激动剂resveratrol预处理可上调SIRT1蛋白水平(P<0.01),而抑制p53的乙酰化(P<0.05),并且凋亡相关指标Bax(P<0.05)、c-caspase-3(P<0.05)、c-PARP(P<0.05)和γ-H2AX(P<0.01)的表达水平也相应降低。
免疫荧光实验结果(图4B)进一步证实,SIRT1激动剂resveratrol预处理能减弱Sev联合N2O处理后神经元细胞的凋亡;而SIRT1拮抗剂salermide和sirtinol预处理则在吸入麻醉药引起细胞凋亡的基础上进一步增加凋亡相关指标的表达。以上结果提示,吸入麻醉引起的海马神经元细胞中SIRT1蛋白水平的反馈性升高具有神经保护作用,而抑制SIRT1表达将加重吸入麻醉导致的神经元细胞凋亡。
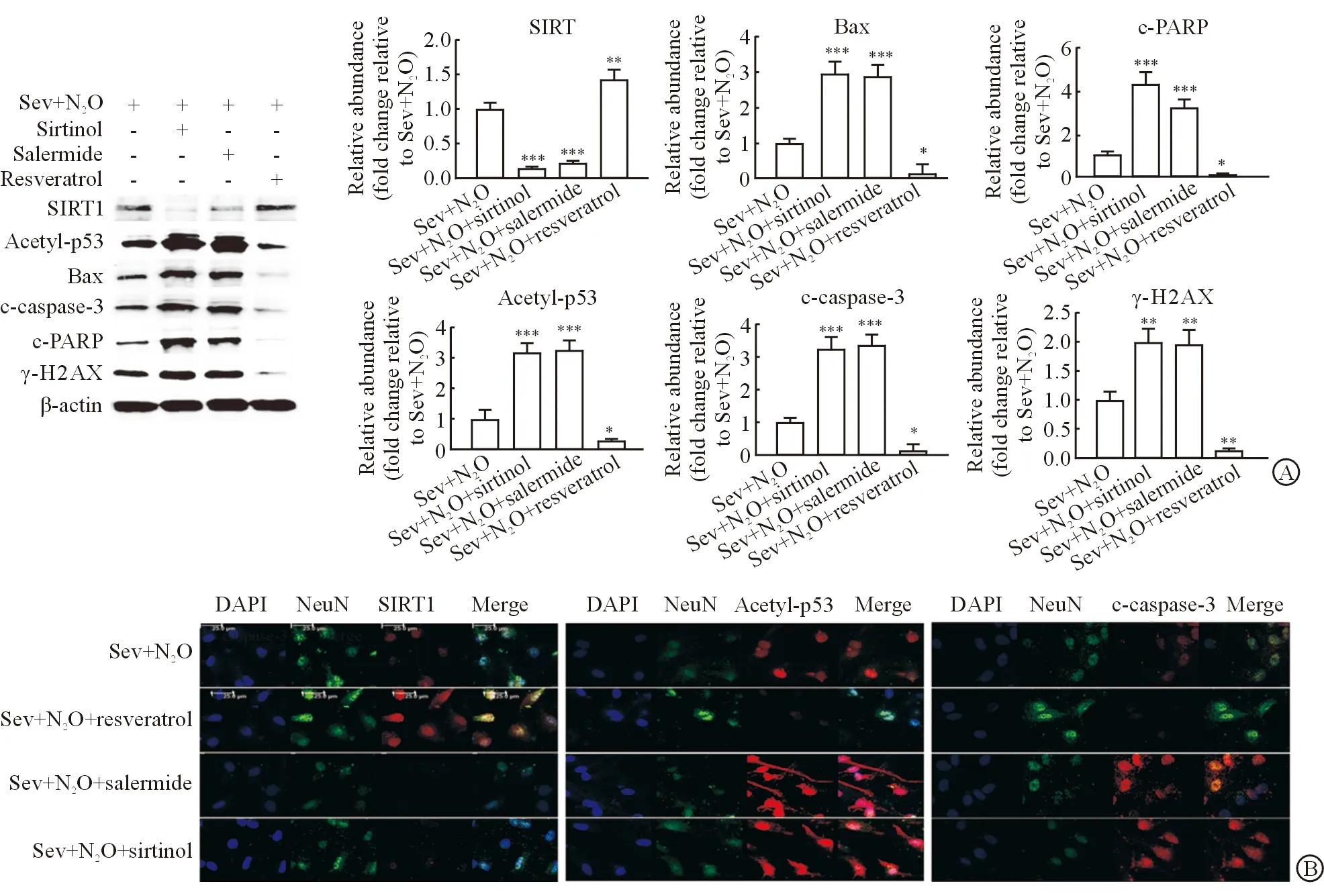
图4 激活或抑制SIRT1通路对Sev联合N2O麻醉诱导原代海马神经元细胞凋亡相关蛋白水平的影响
A:蛋白质印迹法检测SIRT1、Acetyl-p53、Bax、c-caspase-3、c-PARP和γ-H2AX 的表达;B:免疫荧光实验检测SIRT1、Acetyl-p53和c-caspase-3蛋白表达.Sev+N2O:1.3%Sev联合50%N2O+50%O2吸入麻醉处理;Sev+N2O+sirtinol:先用SIRT1拮抗剂sirtinol预处理,再用1.3%Sev联合50%N2O+50%O2吸入麻醉处理;Sev+N2O+salermide:先用SIRT1拮抗剂salermide预处理,再用1.3%Sev联合50%N2O+50%O2吸入麻醉处理;Sev+N2O+resveratrol:先用SIRT1激动剂resveratrol预处理,再用1.3%Sev联合50%N2O+50%O2吸入麻醉处理.*P<0.05,**P<0.01,***P<0.001与对照组(Sev+N2O)相比
3 讨 论
近年来研究发现Sir2蛋白家族在延长寿命方面具有一定的功能,是目前医学研究的热点之一[16]。SIRT1作为人类Sir2同源蛋白质之一,在发育期大脑中具有广泛的表达,并且中枢神经系统受损后被激活的SIRT1具有神经保护作用[17-18]。有研究[19]显示,SIRT1在AD患者的大脑皮质内存在明显的下降趋势,这可能与tau和Aβ蛋白的积聚及认知功能损害密切相关。另外,在AD动物模型中,过表达SIRT1能降低Aβ水平,而敲除SIRT1则能引起Aβ水平升高[20]。另一项关于Huntingtin疾病的研究[21]也显示,过表达SIRT1可减轻亨廷顿蛋白(huntingtin protein)介导的代谢异常,减少大脑萎缩,并改善运动功能。事实上,SIRT1抑制神经退行性变的功能是其具有延长寿命作用的机制之一;另外SIRT1还具有调节神经元分化,以及在海马缺血预处理中发挥神经保护的作用[22-23]。
组蛋白的乙酰化修饰对基因表达具有重要的调控作用,是学习与记忆的重要部分[24]。同时,在记忆的巩固中,组蛋白乙酰化通过调节染色质重塑也发挥了重要作用,而SIRT1在染色质重塑及组蛋白乙酰化过程中均扮演着重要角色[25]。本研究以原代海马神经元细胞为研究对象,结果显示Sev联合N2O吸入麻醉能促进原代海马神经元细胞的凋亡,同时也可引起SIRT1表达增加。鉴于SIRT1的神经保护作用,本研究进一步给予SIRT1抑制剂sirtinol、salermide或激动剂resveratrol共培养后进行吸入麻醉干预,以期双向验证Sev联合N2O吸入麻醉引起的SIRT1上调对其诱导的神经细胞凋亡的影响,结果初步阐明了上调SIRT1表达可以减弱Sev联合N2O吸入麻醉的促神经细胞凋亡作用,表明SIRT1具有潜在的神经保护作用。
目前已有的文献报道[26-27]显示,SIRT1能使p53在内的多种蛋白发生去乙酰化。p53蛋白在进化上相对保守,p53乙酰化修饰对其功能具有重要的调节作用,并且在p53依赖的细胞凋亡途径中发挥重要作用。当一些信号刺激引起DNA损伤时,一方面p53发生乙酰化,促进其下游促凋亡基因的激活或转录,进而引起细胞凋亡;另一方面,机体也会同时启动自我修复机制,上调SIRT1的表达,以维持p53的去乙酰化状态,使p53活性下降,导致依赖于p53的Bax等促凋亡基因的转录被抑制,从而保护细胞免受凋亡损害[28]。研究[29]显示,通过 SIRT1 对 p53 的去乙酰化可以抑制p53 介导的细胞凋亡而发挥其促神经分化与存活的作用,而SIRT1 基因敲除时将增加 p53 的乙酰化及神经元细胞的凋亡。由此可见,SIRT1 通过 p53 去乙酰化平衡在神经元生存中发挥了重要的作用。本研究中Sev联合N2O吸入麻醉处理在引起SIRT1表达水平上调的同时,也促进了p53的去乙酰化,并且进一步使用SIRT1的激动剂和拮抗剂证明Sev联合N2O吸入麻醉处理后神经细胞内上调的SIRT1通过去乙酰化p53发挥其抗凋亡作用,从而减缓吸入麻醉所致神经细胞凋亡。
本研究用从新生大鼠中提取的原代海马神经元细胞作为体外模型,验证了Sev联合N2O吸入麻醉可致神经元细胞凋亡及DNA损伤,但与此同时也可反馈性地促进SIRT1的表达,上调的SIRT1进一步促进p53的去乙酰化,从而缓解Sev联合N2O吸入麻醉诱导的神经元细胞凋亡。因此,本研究阐明了SIRT1-p53信号通路在Sev联合N2O吸入麻醉所致大鼠原代海马神经元细胞凋亡中具有神经保护作用。该结果为临床上减轻吸入麻醉药引起的神经细胞损伤提供了一个潜在的治疗靶点。
