D-氨基酸脱氢酶在不同宿主中发酵产酶条件优化
2020-01-03
(湖南福来格生物技术有限公司,湖南 长沙 422010)
D-氨基酸脱氢酶(D-amino acid dehydrogenase,DAADH)属于氧化还原酶类,是一种细菌膜蛋白,能以NADH或NADPH为辅酶可逆的催化D-氨基酸氧化成相应的亚氨基酸,再水解为酮酸和氨[1-2]。DAADH广泛存在于芽孢杆菌属、链球菌属等微生物体内。1985年,Ohshima[3]在球形芽孢杆菌内发现了氨基酸脱氢酶。随着研究的不断深入,来自枯草芽孢杆菌、地衣芽孢杆菌、蜡样芽孢杆菌、嗜盐链球菌不同来源的氨基酸脱氢酶陆续被发现。DAADH在制药工业和医学领域具有广泛的应用前景[4-5],还被应用于生物传感器的设计,可检测溶液中尤其是血清中某些游离D-氨基酸的含量[6],DAADH也被用于工业氨基酸的合成[7]。由于DAADH具有立体特异性[8],可将它们用于合成光学纯的食品、医药等领域的D-氨基酸。人工合成的D-氨基酸[9]除了具有天然氨基酸的大部分功能外,还具有天然氨基酸所不具备的优良性能,在药物合成(医药和农药)、食品、化妆品等方面具有特殊用途[10]。
本实验室筛选到能高表达DAADH的大肠杆菌和毕赤酵母。大肠杆菌的细胞壁含有D-氨基酸,DAADH会分解D-氨基酸,从而使大肠杆菌破胞,导致清液酶活升高,不利于下游提取纯化,也限制了细胞内酶活提升,但大肠杆菌发酵时间短。毕赤酵母属于真菌,真菌细胞壁中不含D-氨基酸,DAADH不会使毕赤酵母产生破胞问题,但发酵时间长。笔者旨在通过对大肠杆菌产DAADH和毕赤酵母产DAADH的发酵工艺条件进行优化,为工业生产提供参考依据。
1 材料与方法
1.1 菌 种
大肠杆菌BL21(DE3)、毕赤酵母X-33,由本实验室构建并保藏。
1.2 培养基
大肠杆菌摇瓶培养基:5 g/L酵母粉,10 g/L蛋白胨,10 g/L氯化钠。
大肠杆菌发酵罐培养基:10 g/L甘油,10 g/L酵母粉,7.5 g/L蛋白胨,7.5 g/L磷酸氢二钾,7.5 g/L磷酸二氢钾,2 g/L柠檬酸,10 g/L硫酸铵, 10 g/L氯化钠,0.5 g/L七水硫酸镁, 1 mL/L微量元素,初始pH 7.0。
酵母摇瓶培养基:YPD培养基,10 g/L酵母粉,20 g/L蛋白胨,20 g/L葡萄糖。
酵母发酵罐培养基:15 g/L硫酸镁,2.5 g/L硫酸钙,1.5 g/L NaCl,50 g/L甘油,6 g/L KOH,10 mL/L磷酸,3 mL/L微量元素,1.0 mL/L生物素。初始发酵pH 5.0。
1.3 控制工艺
1.3.1 大肠杆菌发酵工艺控制
发酵液3.0 L,初始空气流量200 L/h,罐体压力0.015 MPa左右,转速200 r/min,根据溶氧值将搅拌转速升至800 r/min,转速调满后增加空气流量至350~400 L/h,调整罐压为0.05 MPa。氨水控制pH 7.0,溶氧反弹后补加浓度为700 g/L的葡萄糖,诱导前比生长速率控制在0.2~0.3,OD600达到40左右缓慢降温诱导,诱导后比生长速率控制在0.1~0.2,逐步降低,直至45 h放罐。
1.3.2 酵母发酵工艺控制
发酵液2.5 L,初始空气流量200 L/h,罐体压力0.015 MPa左右,转速200 r/min,溶氧串级至800 r/min,转速调满后增加空气流量至350~400 L/h,调整罐压为0.05 MPa。溶氧反弹后补加甘油,甘油补加6~7 h后,控制罐内溶氧为30%左右,保持2 h,再单独补加甲醇。补加甲醇采用反向关联补料,即溶氧高于30%即补料,溶氧稳定30%左右,发酵时间为114 h。
1.4 检测方法
1.4.1 发酵液OD600值检测
取一定量发酵液,稀释适当倍数(保证读数在0.2~0.8),以去离子水为对照,测定OD600的吸光值。
1.4.2 DAADH酶活检测
DAADH酶活包括混液酶活和清液酶活。混液酶活指均匀取的发酵液的酶活,其中包括菌体;清液酶活是发酵液离心后的上清液酶活。由于DAADH的提取工艺是离心收集菌体后破碎提取,因此上清液酶活越低越好。在一定反应条件下,单位酶量每分钟消耗1 μmol的NADPH为一个单位。25 ℃液态酶活力计算公式为
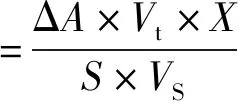
式中:ΔA表示每分钟吸光度的变化值,即为斜率;S表示NADPH的摩尔消光系数,通过制作标准曲线求得;Vt表示反应液的总体积,mL;Vs表示样品酶液的体积,mL;X表示样品酶液的稀释倍数。
2 结果与讨论
微生物的发酵工艺会影响工程菌的重组蛋白表达量,对产酶影响较大。在工程菌构建完成后,外源蛋白的表达量往往达不到工业化生产所要求的水平,因此需要对其发酵工艺进行优化[11]。微生物发酵过程的机理复杂,并受到微生物内部代谢调控机制和外界环境(例如培养基组成与配比、发酵温度和pH)等因素的影响。对于工程菌来说,菌种对质粒的稳定性要求则更加严苛。利用工程菌进行高密度发酵,它的生长和外源蛋白的表达水平与目的蛋白本身及其遗传特性有很大关系(如外源基因拷贝数)。
本实验室使用的发酵培养基配方能满足菌种的生长和产酶需要,因此主要从产酶的诱导条件入手进行优化。
2.1 大肠杆菌产DAADH发酵工艺优化
2.1.1 大肠杆菌产DAADH诱导温度优化
微生物生长和产酶过程中对温度比较敏感,温度会影响微生物代谢过程中相关酶的活性。微生物生长最适温度与诱导最适温度往往不同,一般采用前期高温培养,后期低温诱导策略[12]。一定范围内,高温能加快菌体生长代谢,缩短培养时间,但随着温度升高,酶活下降,菌体容易衰老,蛋白表达量降低。适当的诱导温度还可以有效地减少包涵体的产生[13]。
分别选取22,25,28,32 ℃ 4个温度进行实验。诱导温度对大肠杆菌产DAADH的影响如图1所示。
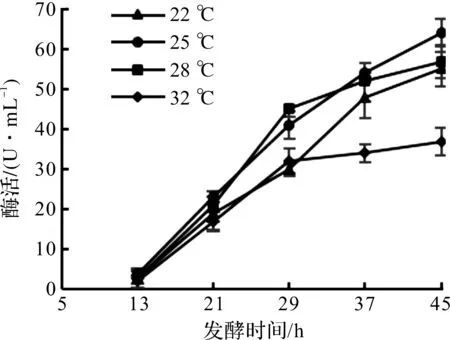
图1 诱导温度对大肠杆菌产DAADH的影响Fig.1 Effect of induction temperature on DAADH production by Escherichia coli
由图1可知:在32 ℃条件下,前期还可以正常产酶,29 h后酶活增长速度趋于平缓。理论上温度高,会有利于微生物代谢,但合成目的蛋白质时,速度过快会造成蛋白折叠异常,从而没有活性,所以高温度往往不利于产酶。22,25,28 ℃ 3个温度产酶趋势比较一致,随着发酵进行,酶活逐渐增加,发酵后期的增长速度放慢。25 ℃会更有优势,45 h放罐酶活达到了64 U/mL。
2.1.2 大肠杆菌产DAADH诱导pH优化
发酵过程中随着微生物代谢进行,发酵液pH值会随之改变[14]。大肠杆菌碳源分解代谢主要经过柠檬酸循环,代谢产物大部分为有机酸,会使发酵液pH值降低,因此需要加碱来维持pH值恒定。本实验用氨水来调节pH值,分别选定pH 6.0,6.5,7.0,7.5 这4个条件进行试验,诱导pH对大肠杆菌产DAADH的影响如图2所示。
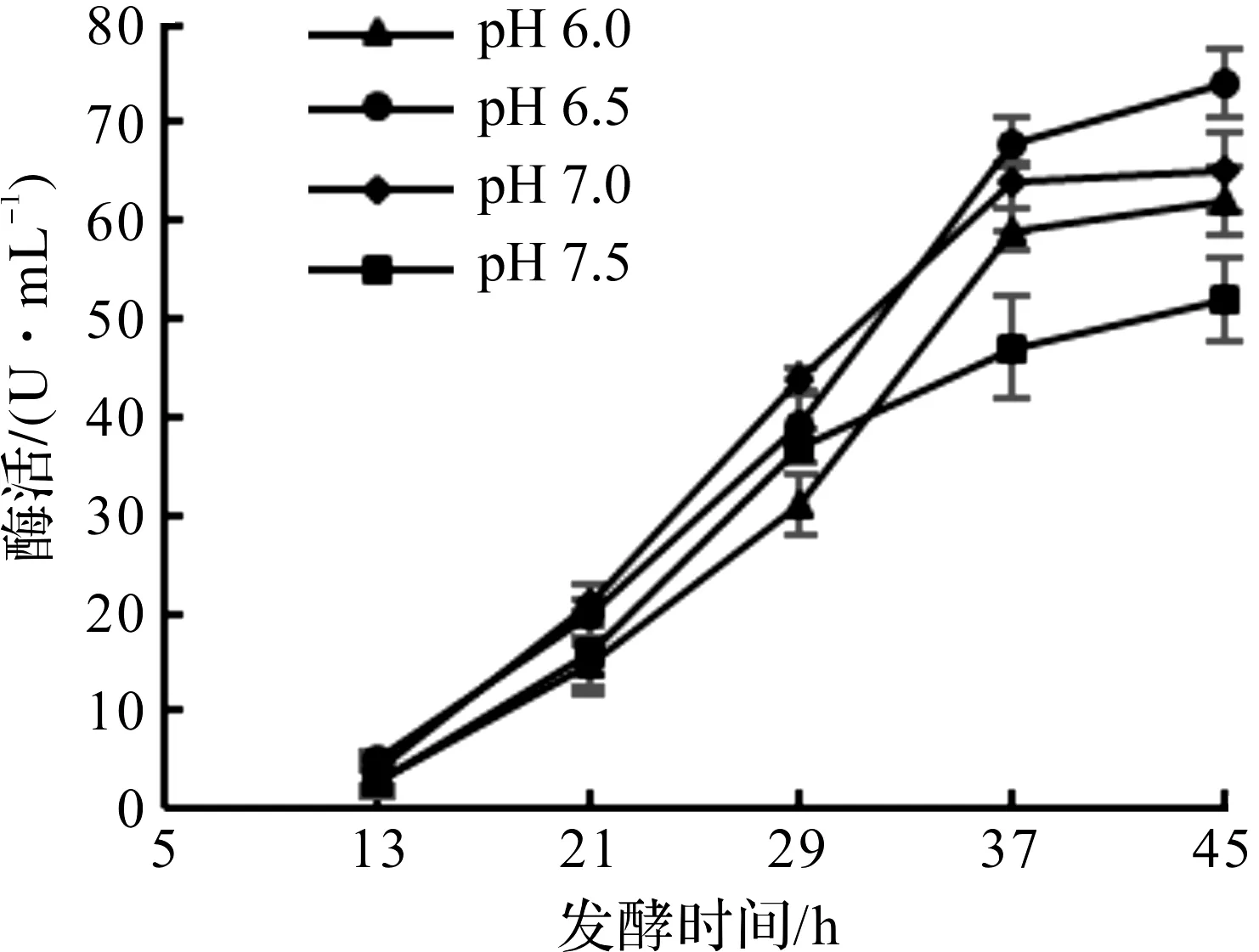
图2 诱导pH对大肠杆菌产DAADH的影响Fig.2 Effect of induced pH on DAADH production by Escherichia coli
由图2可知:当pH为6.5时,酶活为74.0 U/mL,比其他3个pH条件下的酶活高,而pH为7.5时则最低,放罐酶活只有52.0 U/mL,比pH为6.5时低了30%。
2.1.3 诱导大肠杆菌产DAADH起始OD600优化
对数生长期的微生物活力较高,环境适应能力较强,对重组大肠杆菌进行诱导一般选在这一时期[15]。实验分别选取诱导OD600值为30,35,40,45,起始诱导OD600对大肠杆菌产DAADH的影响如图3所示。
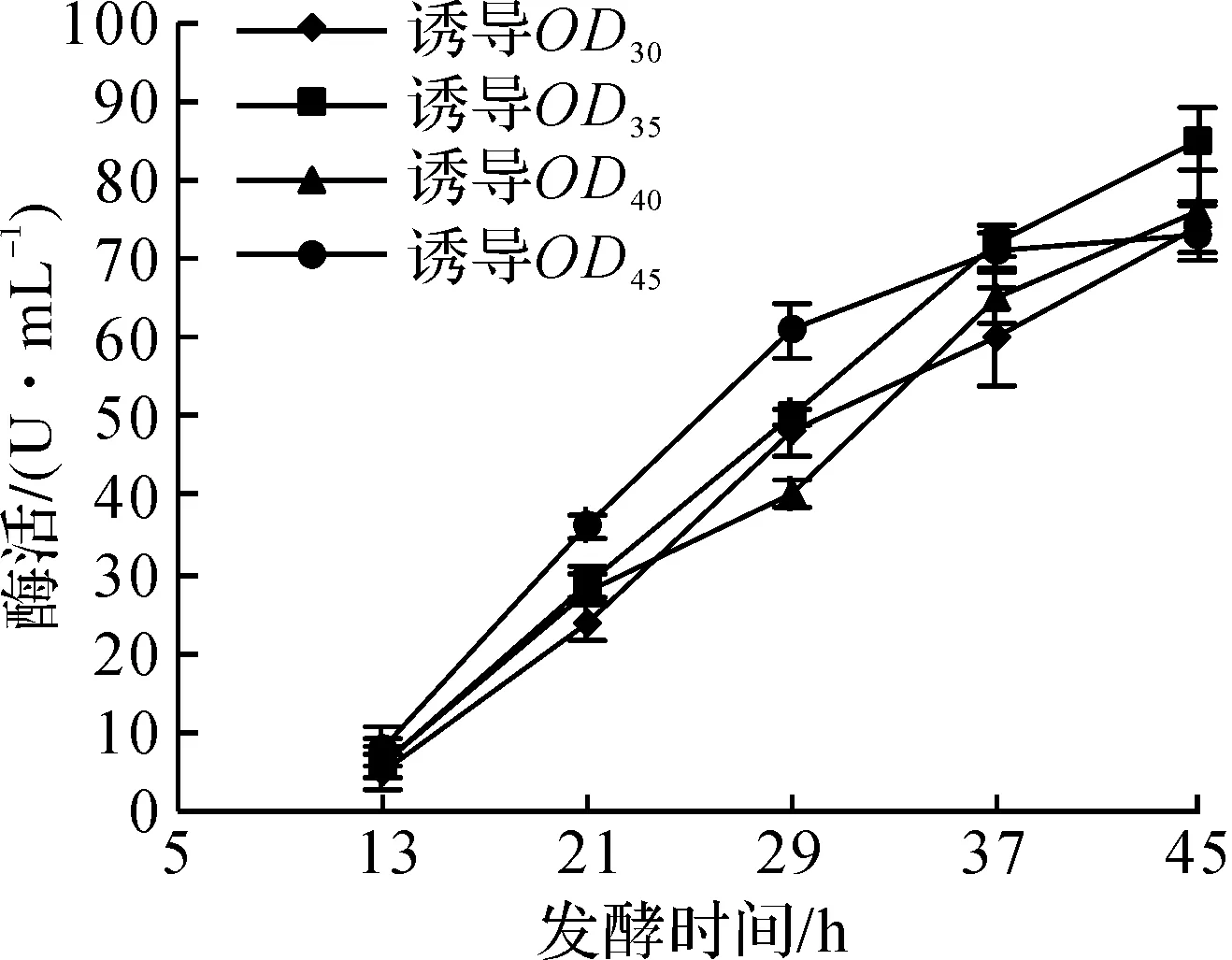
图3 起始诱导OD600对大肠杆菌产DAADH的影响Fig.3 Effect of induced initial OD600 on DAADH production by Escherichia coli
由图3可知:当诱导OD600为35时,酶活最高,达到了85.0 U/mL,比OD600为30,40,45时分别高了13.0%,10.6%,14.1%。结合实验和生产情况来看,由于种子、接种量以及罐子的差异,大肠杆菌生长状况并不完全相同,因此诱导OD600的值只是一个范围。根据实验结果,诱导OD600的值为35左右为最佳。
2.1.4 大肠杆菌产DAADH发酵时间优化
大肠杆菌的细胞壁由肽聚糖构成,肽聚糖结构中含有D-氨基酸,DAADH能特异性的将D-氨基酸氧化成相应的亚氨基酸,再水解为酮酸和氨,从而裂解大肠杆菌的细胞壁,使细胞失去支撑从而破胞。酶活越高,破胞问题越严重,为解决这一问题,需要让大肠杆菌在一定时间内大量产酶,并在产酶速率放缓以后的合适时间内放罐。
在连续监控3批发酵的发酵液混液酶活和离心清液酶活后,以DAADH酶活为纵坐标,以发酵时间为横坐标作图,大肠杆菌产DAADH的发酵时间优化如图4所示。
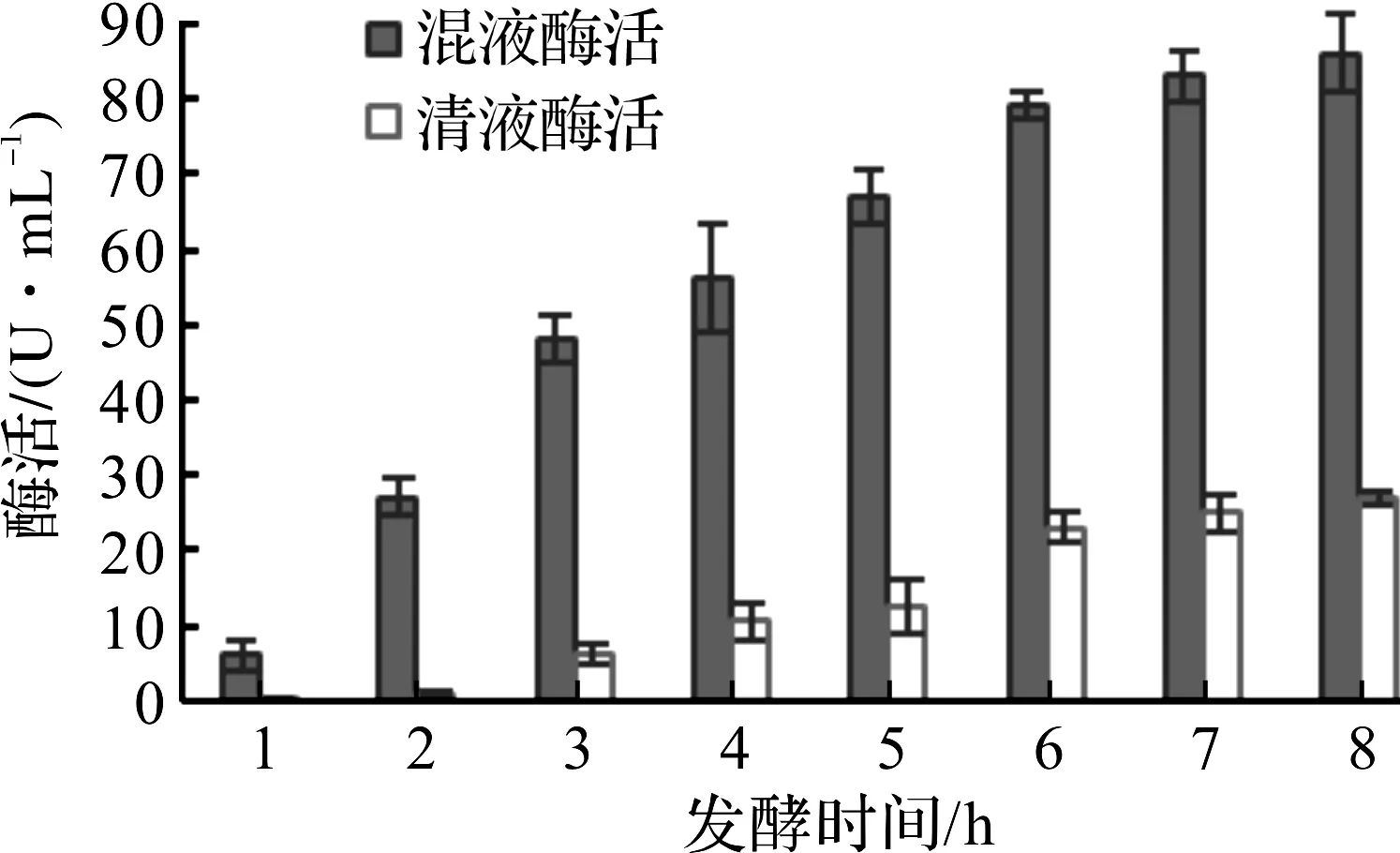
图4 大肠杆菌产DAADH的发酵时间优化Fig.4 Optimization of fermentation time for DAADH production by Escherichia coli
由图4可知:在35,38,41 h时,混液酶活分别为67.2,79.4,83.0 U/mL,离心清液酶活占发酵混液酶活的比例分别为18.6%、24%、35%。35 h清液酶活低,但混液酶活也不高,41 h混液酶活高,但清液酶活占了35%,38 h混液酶活高,清液酶活也在20%左右,因此在38 h左右放罐比较合适。
2.2 毕赤酵母产DAADH发酵工艺优化
大肠杆菌细胞壁含有D-氨基酸,破胞问题势必会存在,而且随着酶活的提高,破胞问题也会加重,虽然提前放罐能在一定程度上缓解破胞问题带来的负面影响,但同时也会造成资源和设备浪费,导致成本偏高。毕赤酵母属于真菌,真菌细胞壁中主要成分为几丁质、纤维素、葡聚糖、甘露聚糖等,这些多糖都是单糖的聚合物,不存在DAADH破坏细胞壁而导致细胞破裂的情况[16]。
2.2.1 毕赤酵母产DAADH诱导温度优化
毕赤酵母是甲醇营养型酵母中的一类能够利用甲醇作为唯一碳源和能源的酵母菌。它的最佳生长温度范围是28~30 ℃,在诱导产酶阶段,在20~30 ℃范围都可以正常产酶。诱导温度对毕赤酵母产DAADH的影响如图5所示。
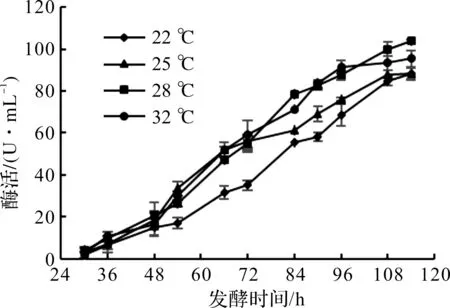
图5 诱导温度对毕赤酵母产DAADH的影响Fig.5 Effect of induction temperature on DAADH production by Pichia pastoris
由图5可知:毕赤酵母在诱导阶段酶活一直呈上升趋势,前期增速较慢,中期酶活提高迅速,后期趋于平缓。在发酵过程中,22 ℃条件下酶活比其他3个温度条件酶活低,说明低温不利于毕赤酵母产DAADH。而在28 ℃条件下,114 h放罐酶活最高,为103.7 U/mL,因此选择诱导温度为28 ℃。
2.2.2 毕赤酵母产DAADH诱导pH优化
pH值会改变细胞膜的通透性,影响生物半透膜的正常结构和功能,使其通透性增加,胞内物质流出细胞[17],从而影响毕赤酵母的正常代谢和DAADH的合成。诱导pH对毕赤酵母产DAADH的影响如图6所示。
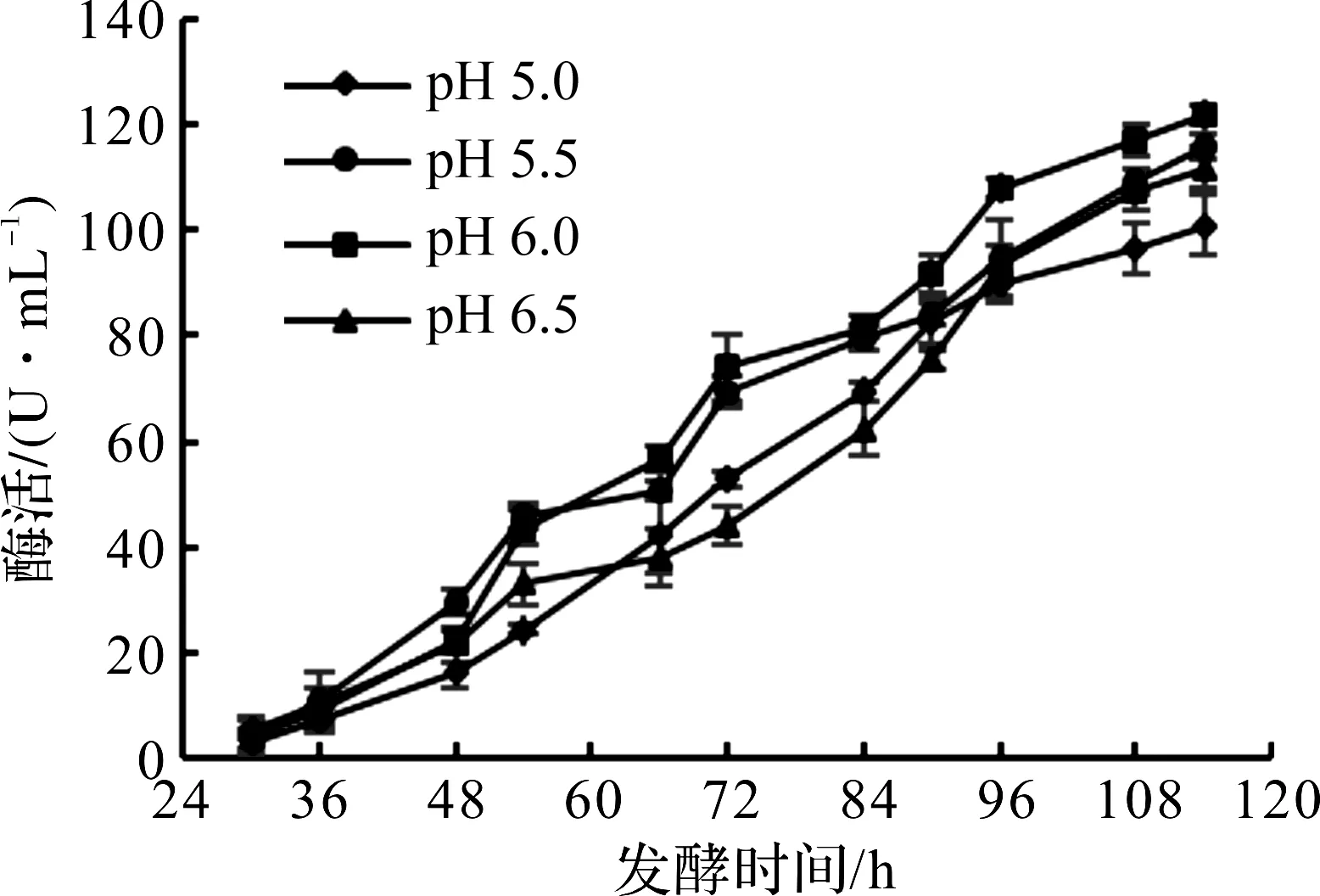
图6 诱导pH对毕赤酵母产DAADH的影响Fig.6 Effects of induced pH on DAADH production by Pichia pastoris
由图6可知:pH 6.0的酶活为121.7 U/mL,此时酶活最高,比pH 5.0,5.5,6.5时分别高17.3%,4.9%,8.3%。
2.2.3 毕赤酵母产DAADH溶氧优化
重组毕赤酵母是甲醇营养型工程菌,以AOX1为启动子,当甲醇作为唯一碳源时,毕赤酵母能被甲醇诱导启动外源蛋白的表达。毕赤酵母的发酵产酶分为两个阶段[18],第一阶段主要是菌体生长,以甘油为碳源,第二阶段以甲醇为碳源,并诱导AOX基因驱动外源基因表达。由于过高浓度的甘油和甲醇都会抑制毕赤酵母细胞的正常生长和产物的顺利表达,所以需要通过合适的补料策略控制甘油和甲醇的流加,达到显著提高外源蛋白表达量的目的。分批补料发酵模式简单、高效,是毕赤酵母细胞高密度发酵最常用发酵模式。在甲醇诱导阶段[19],由于重组菌的物质和能量代谢、比生长速率及生理状态的改变,重组菌对碳源的需求量也会产生很大的改变。溶氧值对毕赤酵母产DAADH的影响如图7所示。
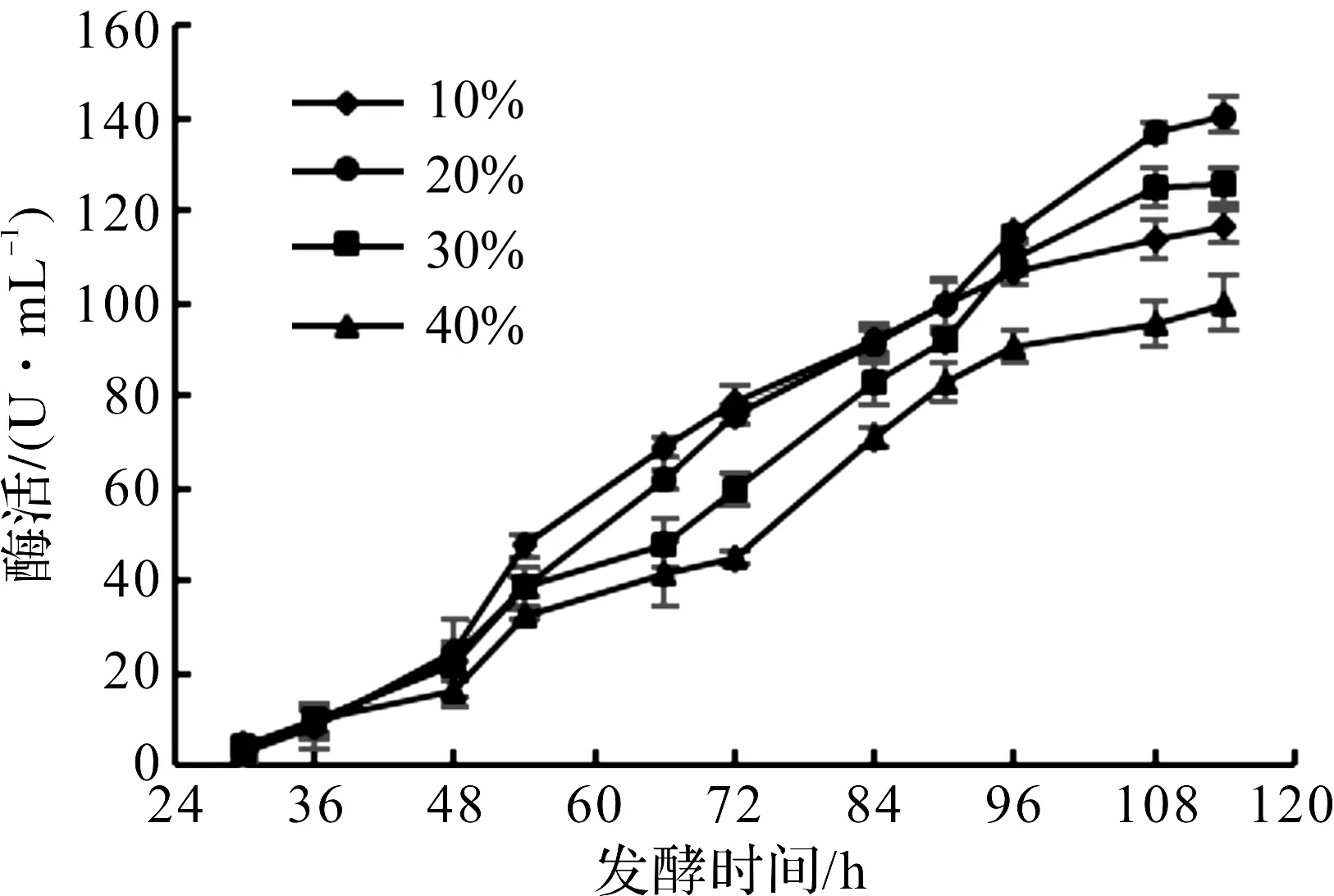
图7 溶氧值对毕赤酵母产DAADH的影响Fig.7 Effect of dissolved oxygen value on DAADH production by Pichia pastoris
由图7可知:当溶氧控制在10%时,前期酶活增长较快,后期酶活增长趋于平缓;当溶氧控制在40%时,整个发酵过程中酶活都是最低的。所以甲醇加量过高过低都不利于毕赤酵母合成DAADH,甲醇浓度过高会毒害菌体,影响代谢;甲醇浓度过低会使醇氧化酶与目的蛋白减少[20],也不利于产酶。当关联补料控制溶氧为20%时,酶活最高,达到140.5 U/mL,综上毕赤酵母诱导阶段溶氧控制为20%左右。
3 结 论
通过对大肠杆菌产DAADH诱导温度、诱导pH、诱导起始OD600值和发酵时间进行优化,在25 ℃、pH 6.5和OD600值为35的条件下诱导,培养45 h酶活为86.0 U/mL,优化前为64.1 U/mL,提高了34.4%。大肠杆菌细胞壁中的D-氨基酸会被DAADH分解从而导致细胞破裂,酶活越高,破胞现象越严重。经过优化确定38 h左右放罐,此时DAADH混液酶活为79.0 U/mL左右,清液酶活占20%。毕赤酵母属于真菌,不存在DAADH破坏细胞壁而导致细胞破裂的情况。将连接目的基因的重组质粒转化到毕赤酵母菌中,对毕赤酵母产DAADH诱导温度、诱导pH和溶氧值进行优化,确定诱导条件为28 ℃、pH 6.0、溶氧值为20%,放罐时DAADH酶活为140.5 U/mL,比优化前提高了58.9%。大肠杆菌和毕赤酵母发酵产DAADH,酶活都能达到较高水平。大肠杆菌发酵时间短,设备利用率高,但存在破胞问题,不利于下游酶的提取;毕赤酵母不存在DAADH导致的破胞问题,且市面上NADPH售价高,而酵母菌体中含有大量的NADPH,对需要NADPH参与循环的反应,用酵母菌体破碎后直接参与反应,能免去额外投加NADPH的费用,但酵母的DAADH酶活是大肠杆菌的1.8倍,发酵时间却是大肠杆菌的2.5倍,设备利用率不高,可以根据用途选择大肠杆菌或者毕赤酵母来进行发酵。
