杂色角孔海胆形态学观察、亲缘关系及性腺营养成分分析*
2019-12-06赵文飞黄小芳常亚青
赵文飞 李 喆 黄小芳 张 杨 丁 君 常亚青
杂色角孔海胆形态学观察、亲缘关系及性腺营养成分分析*
赵文飞 李 喆 黄小芳 张 杨 丁 君①常亚青①
(大连海洋大学 农业农村部北方海水增养殖重点实验室 大连 116023)
本研究以杂色角孔海胆()为研究材料,观察其形态,基于线粒体16S rDNA基因部分序列分析杂色角孔海胆与其他棘皮动物的亲缘关系,同时分析杂色角孔海胆性腺中的营养成分,为杂色角孔海胆种质改良和资源评估积累数据。结果显示,杂色角孔海胆的主要形态特征具有口器较小,每个步带板的外缘有3~4个小的圆形凹孔,管足壁内的骨片为C形。通过Mega 5.0对其他棘皮动物和得到的杂色角孔海胆的线粒体16S rDNA基因部分序列进行比较,基于距离法(NJ)和最大似然法(MP)分别构建系统树。结果显示,杂色角孔海胆与疏棘角孔海胆(的亲缘关系最近,其次是刻肋海胆属的3种海胆;杂色角孔海胆性腺中水分为86.1%,灰分为3.0%,粗脂肪为2.0%,蛋白质为8.01%;杂色角孔海胆性腺中的氨基酸种类共有16种,总含量为38.71 g/100 g,EAA/TAA为36.99%,EAA/NEAA为71.84%,DAA/TAA为37.17%;脂肪酸共检测到15种,含量最高的是棕榈酸C16:0、硬脂酸C18:0、EPA C20:5和花生四烯酸C20:4,含量最低的为二高-γ-亚麻酸C20:3,较其他海胆,其DHA含量较高。研究结果将为杂色角孔海胆的生物学研究提供基本资料,可用于经济海胆开发应用,同时也可为远缘杂交育种及经济海胆的种质鉴定提供理论依据。
杂色角孔海胆;形态学;性腺营养成分;线粒体;16S rDNA
海胆属于棘皮动物门(Echinodermata)、游在亚门(Eleutherozea)、海胆纲(Echinoidea),是常见的一类海洋无脊椎动物,具有较高的食用、药用和科研价值。全世界现存的海胆大约有850种,已被较好利用的经济种类不超过30种,可食用型海胆仅10余种。目前,我国主要的经济海胆种类有球海胆科的中间球海胆 ()、马粪海胆()和光棘球海胆();长海胆科的紫海胆();疣海胆科的海刺猬()和毒棘海胆科的白棘三列海胆()(常亚青等, 2004; 左然涛等, 2016)。
杂色角孔海胆()隶属棘皮动物门、海胆纲、拱齿目、刻肋海胆科、角孔海胆属,是我国南海普遍种。海胆的性腺部分可食用,在亚洲、地中海国家以及智利等西半球国家被认为是一种珍贵的美味。亚太地区的人们多年来也一直使用海胆性腺作为改善一般身体状况、治疗多种疾病的药物。杂色角孔海胆在海南或南海地方百姓食用的人较少,在香港有人食用杂色角孔海胆(Chen, 2010)。目前角孔海胆属的海胆均未得到较好的开发,其中角孔海胆属包括3个种:模式角孔海胆、条纹角孔海胆、杂色角孔海胆(刘瑞玉, 2008)。有关杂色角孔海胆生物学方面的研究主要集中在发育生物学方面,Rahman等(2014、2016)研究了杂色角孔海胆个体发育过程,Subramaniam (1934)发现人工养殖条件下杂色角孔海胆最适合以红藻为食,付婉莹(2015)观察了杂色角孔海胆棘的显微结构,而在系统发育学和性腺营养成分组成方面的研究较少。随着生物技术的发展,16S rDNA在水产动物中的分类应用研究越来越重要,已经广泛应用于各种水生生物的分类研究,对水产养殖业的发展起到了重要的作用(周先文等, 2009)。
本研究对杂色角孔海胆开展形态学观察并测定其线粒体DNA 16S rDNA部分基因序列,分析与其他棘皮动物亲缘关系的远近,同时对杂色角孔海胆性腺营养成分进行分析。研究结果为角孔海胆属的生物学研究提供基础,为进一步开发利用角孔海胆属的海胆提供信息。
1 材料与方法
1.1 实验材料
实验所用杂色角孔海胆样品于2018年8月采自海南三亚市。取鲜活海胆的性腺、管足及口器,迅速投入液氮中冷冻后,–80℃保存以供后续进行性腺营养成分测定和线粒体DNA 16S rDNA部分基因序列分析。海胆壳用海水浸泡3 d,待其表面的棘全部脱落,再用清水冲洗干净,晾干保存用于海胆形态观察,同时对海胆的壳性状和性腺性状进行测量。
1.2 实验方法
1.2.1 形态学观察 对10个海胆壳进行测定,测定了22个性状。使用数显电子游标卡尺(Mahr 16ER,精确度0.01 mm)分别测量壳长、壳宽、壳高、口径、步带(赤道部)宽、间步带(赤道部)宽、顶系长、顶系宽、口器高、口器宽。用电子天平(G&GJJ100,精准度0.01 g)测量分别测量体重、壳干重、口器重、性腺重。性腺颜色用分光色差仪CM-2600d/2500d测定。采用Nikon DS100数码相机拍照。
1.2.2 线粒体16S rDNA基因片段的PCR扩增及序列测定 采用生工Ezup柱式动物组织基因组DNA抽提试剂盒提取海胆管足中的DNA。16S rDNA扩增引物序列为:F: 5¢-CGCCTGTTTAACAAAAACAT-3¢,R: 5¢-CCGGTCTGAACTCAGATCATG-3¢(Gao, 2003)。反应总体系为50 μl,其中20~50 ng/μl模板DNA 2 μl、10 μmol引物F 2 μl、10 μmol引物R 2 μl、10 mmol/L dNTP(mix)2 μl、10×Buffer 5 μl、5 U/μl酶0.5 μl、ddH2O补足至50 μl。反应条件为:95℃预变性3 min,94℃变性30 s,55~60℃退火30 s,72℃延伸30 s后进行35个循环,72℃修复延伸5~8 min,4℃保存。PCR产物取5 μl,用1%琼脂糖凝胶电泳观察,电泳参数为150 V,100 mA,20 min。PCR产物回收纯化,将纯化后的产物进行测序。
采用MEGA 5.0软件分析序列的碱基组成以及它们的遗传距离,按照Kimura双参数法计算物种间的DNA序列差异及种间遗传距离。调取GenBank中其他棘皮动物16S rDNA序列(表1),采用邻接法(Neighbor-Joining, NJ)和最大简约法(Maximum parsimony, MP)分别构建系统发育树,各分支的置信度采用重抽样法(Boot-strap),通过1000次循环评估系统发育树的可靠性。
1.2.3 性腺营养成分测定 性腺水分含量测定参照GB 5009.3-2016《食品中水分的测定》;粗灰分含量测定参照GB 5009.4-2016《食品中灰分的测定》;粗蛋白质含量测定参照GB 5009.5-016《食品中蛋白质的测定》;粗脂肪含量测定参照GB 5009.6-2016;氨基酸含量测定参照GB 5009.124-2016《食品中氨基酸的测定》,将样品经盐酸水解后利用氨基酸自动分析仪测定16种氨基酸,采用分光光度计法测定色氨酸的含量;脂肪酸含量测定参照GB 5009.168-2016《食品中脂肪酸的测定》,样品经过甲酯化后利用气相色谱进行分析,根据出峰时间定性、峰面积定量来测定脂肪酸含量。
2 结果与分析
2.1 海胆形态特征
杂色角孔海胆10个个体的测量数据经整理分析,22个外部形态可度量特征的变化范围和平均值见表2。步带(赤道部)宽和间步带(赤道部)宽的平均值为10.99、25.74,步带区宽约为间步带的2/5~1/2。口径/壳长的平均值为0.28。
2.1.1 外部形态特征 杂色角孔海胆体色为灰橄榄色,壳从呈低半球形到高圆锥形(图1A)。口面平坦且稍向内凹,围口部明显凹陷,略呈五角形(图1B),反面较隆起。步带区由反口面至口面逐渐展宽,步带区与间步带区幅宽不等,赤道部以上步带区宽约为间步带的2/5~1/2,两区隆起程度不同,因此海胆壳外形自口面观接近于圆形。步带区每个板上有1个大疣,在步带排列为一列;板内侧有1个中疣,与大疣平行排为一列;大疣上方及两侧分布着一些小疣。间步带板在赤道部以上每个板上有1个大疣,排列为一列;板的外侧有个1中疣,平行于大疣排列。间步带赤道部以下,各个板上有3个疣排列为一横行。顶系较小,被小棘所掩盖,与围肛部相连接,眼板不接触围肛部,各生殖板上分布着许多疣。大棘一般为绿色,带有深色横带,围口部的大棘略带紫色,中棘大多为赤褐色。反口面的大棘比较尖细,口面大棘稍长,末端扁成刃形。壳表面颜色不均匀,变化较为明显,反口面呈暗绿色或暗灰色,口面颜色较浅多为淡灰色或淡绿色甚至近白色,颜色从反口面向口面逐渐变浅。管足孔每3对排列为一弧,各个步带板的外缘有3~4个小的圆形凹孔(图1C)。
表1 用于系统分析的棘皮物种

Tab.1 Species used for the phylogenetic analysis
表2 杂色角孔海胆形态性状的变化范围和平均值

Tab.2 Variation range and mean value of morphological characters of S. sphaeroides variegate

图1 杂色角孔海胆的图片
A: 反口面;B: 口面;C: 顶系;D: 圆形凹孔;E: 口器;F: 骨片
A: Aboral surface; B: Surface; C: Apical system; D: Circular pit; E: Diameter; F: Ossicle
2.1.2 内部构造特征 本次测量的口径与壳长的比值为0.27~0.30,口位于腹面的正中围口膜的中央部,口器的5个大型齿顶露于口外。口器由5个尖锐的大齿、5个大齿骨、若干个小齿骨组成,齿骨之间有肌肉束相连(图1D)。管足壁内的骨片为C形(图1E)。杂色角孔海胆的体腔隔膜较为坚硬,将体腔分隔成若干部分,分别形成食道腔、围肛腔、生殖腔等(图1F)。食道腔位于围口部内侧,口器周围。围肛腔在顶系的内侧,生殖腔偏向于反口面的各个间步带的区域。
2.2 杂色角孔海胆与其他棘皮动物亲缘关系
2.2.1 线粒体DNA 16S rDNA的PCR结果及序列
DNA模板琼脂糖电泳结果显示DNA条带完整、清晰、无杂带,PCR扩增的片段位于200~500 bp。PCR产物直接测序得到杂色角孔海胆的16S rDNA基因片段长度为444 bp(Genbank登陆号为MK723850)。该序列的A、T、C、G的含量分别为33.8%、27.7%、18.0%和20.5%;A+T及C+G的含量为61.5%和38.5%。
2.2.2 基于16S rDNA序列构建杂色角孔海胆的系统发育树 采用MEGA 5.0软件构建基于NJ法和MP法分别构建系统进化树(图2和图3)。基于2种方法构建的系统进化树基本一致,聚类分析结果为:杂色角孔海胆和疏棘角孔海胆聚为一支,再与芮氏刻肋海胆、哈氏刻肋海胆、细雕刻肋海胆聚为一个亚群,然后是海刺猬、白棘三列海胆,再与马粪海胆、中间球海胆、光棘球海胆和紫球海胆聚为一支,与海星纲的长棘海星、海燕和海参纲的糙刺参、绿刺参及刺参较远。
用MEGA 5.0软件计算16种棘皮动物间的遗传距离(表3)。由表3可见,在16S rDNA水平上各种间的遗传距离为0.04~0.68。其中,光棘球海胆和马粪海胆、光棘球海胆和中间球海胆、杂色角孔海胆和疏棘角孔海胆间距离最小,均为0.04;海燕和糙海参距离最远,为0.68,表现为最大的遗传距离。
2.3 性腺营养成分测定结果
2.3.1 一般营养成分测定结果 杂色角孔海胆性腺中的水分含量为86.1%,灰分为3.0%,粗脂肪为2.0%,蛋白质为8.01%。
2.3.2 性腺氨基酸测定 杂色角孔海胆性腺中的氨基酸种类齐全,共有16种(表4),总含量为38.71 g/100 g,必需氨基酸(EAA)含量为14.32 g/100 g;非必需氨基酸(NEAA)含量为19.93 g/100 g,半必需氨基酸(CEAA)含量为4.46 g/100 g。其中,必需氨基酸含量占氨基酸总量(EAA/TAA)的36.99%,必需氨基酸与非必需氨基酸的比值(EAA/NEAA)为71.84%。杂色角孔海胆的呈味氨基酸(DAA)含量为14.39%,占总氨基酸含量的37.17%。

图2 16种棘皮动物的16S rDNA基因片段的NJ系统树

图3 16种棘皮动物的16S rDNA基因片段的MP系统树
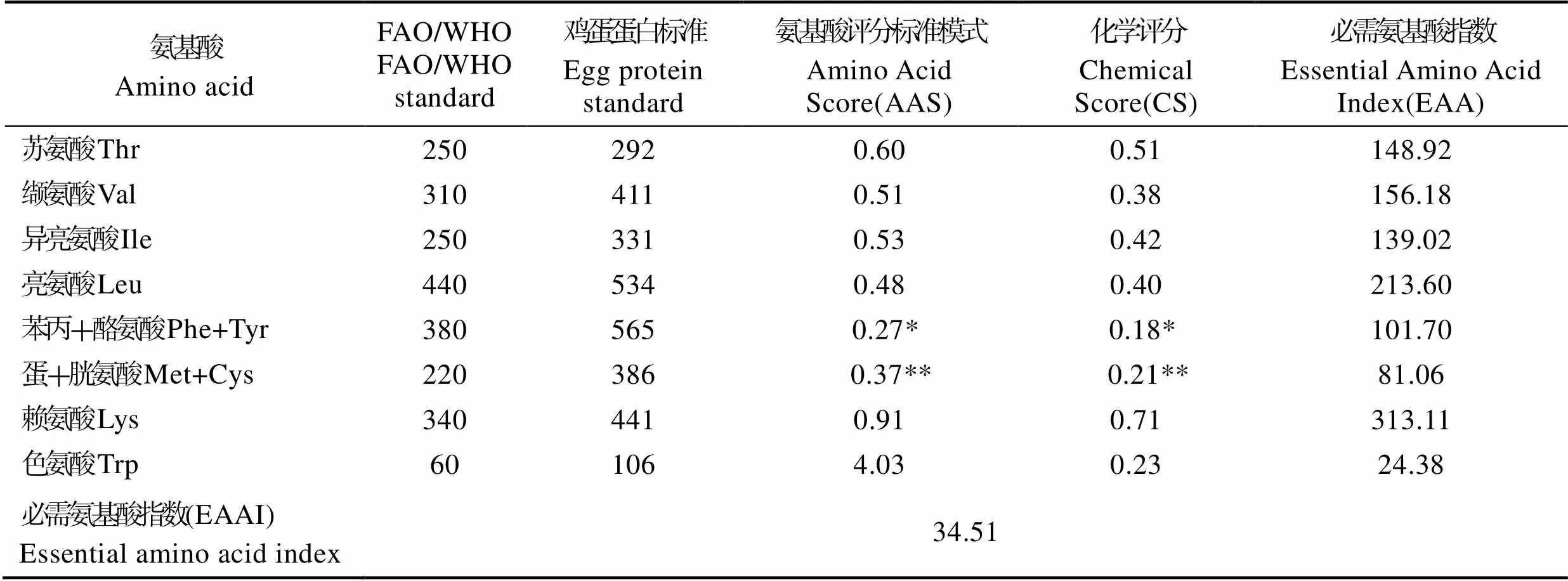
根据联合国粮农组织和世界卫生组织(FAO/WHO)的建议(Vinay, 2008),按照每克氮中氨基酸评分标准模式(Amino Acid Score, AAS)、化学评分(Chemical Score, CS)以及必需氨基酸指数(Essential Amino Acid Index, EAA)进行比较对杂色角孔海胆性腺进行相应的营养评价(见表5)。性腺中必需氨基酸中含量最高的是赖氨酸;非必需氨基酸中含量最高的是甘氨酸,从AAS和CS中得出第一限制氨基酸是苯丙氨酸和酪氨酸,第二限制氨基酸是蛋氨酸和胱氨酸,EAAI值为34.51。
2.3.3 性腺脂肪酸含量测定 杂色角孔海胆性腺中脂肪酸种类较丰富,共检测到16种(表6),总含量为8600.20 mg/kg。杂色角孔海胆性腺中脂肪酸含量最高的依此是棕榈酸C16:0、硬脂酸C18:0、EPA C20:5、花生四烯酸C20:4,杂色角孔海胆的n-3 PUFA/n-6 PUFA比值为0.79。5种饱和脂肪酸、2种单不饱和脂肪酸、 9种多不饱和脂肪酸分别占总脂肪酸的47.5%、4.9%、47.6%,饱和脂肪酸、单不饱和脂肪酸多不饱和脂肪酸分别以C16:0、C20:2、EPA C20:5为主。杂色角孔海胆的DHA含量占总脂肪酸含量的4.7%。
表3 16种棘皮动物16S rDNA基因片段的遗传距离

Tab.3 Genetic distance of 16S rDNA gene fragments between 16 echinoderms
注: 1. 细雕刻肋海胆; 2. 芮氏刻肋海胆; 3. 光棘球海胆; 4. 中间球海胆; 5. 马粪海胆; 6. 紫球海胆; 7. 海刺猬; 8. 糙刺参; 9. 仿刺参; 10. 长棘海星; 11. 白棘三列海胆; 12. 杂色角孔海胆; 13. 疏棘角孔海胆; 14. 海燕; 15. 绿刺参; 16. 哈氏刻肋海胆
Note: 1.2.3.4.5.6.7.8.9.10.11.12.13.14.15.16.
表4 杂色角孔海胆的氨基酸组成(湿重, g/100 g)

Tab.4 Amino acid composition of S. sphaeroides variegate (Wet weight, g/100 g)
注: *为必需氨基酸;**为非必需氨基酸;***为半必需氨基酸;a为鲜味氨基酸
Note: *: Essential amino acids (EAA); **: Non-essential amino acids (NEAA); ***: Semi-essential amino acids (CEAA); a:Delicious amino acid (DAA)
表5 必需氨基酸质量评价
Tab.5 Evaluation of EAA compositions of S. sphaeroides variegate (mg/g N)
注: *为第一限制氨基酸;**为第二限制氨基酸
Note: * means the first limiting amino acids; ** means the second limiting amino acids
3 讨论
3.1 杂色角孔海胆外观形态及系统发育分析
杂色角孔海胆的口器较小,杂色角孔海胆的口径/壳长为0.28,低于马粪海胆(0.31~0.33) (刘成龙等, 2015)、细雕刻肋海胆(0.324)、哈氏刻肋海胆(0.301)以及芮氏刻肋海胆(0.337),这说明杂色角孔海胆的口器与其他几种海胆相比较小。体腔隔膜较为坚硬,这是不同于其他海胆的特征之一。每个步带板的外缘有3~4个小的圆形凹孔,这是角孔海胆属的海胆特有的形态特征。管足孔每3对排列为一弧,这与刻肋海胆中其他几种海胆管足孔的排列方式是一致的。张文峰(2011)研究发现细雕刻肋海胆棘上有深色横带,而其他海胆没有。这与杂色角孔海胆的特征相同,说明杂色角孔海胆与刻肋海胆属亲缘关系较近。在张文峰(2011)的研究中发现赤道部以上步带区宽约为间步带的1/2,但本研究发现这一数值小于1/2。对于这一差异的产生,很有可能是观察设备、样品来源及海胆大小不同造成的。
本研究通过16S rDNA基因片段的系统发育分析表明杂色角孔海胆与疏棘角孔海胆的亲缘关系最近,同属于角孔海胆属,其次是与刻肋海胆的哈氏刻肋海胆、芮氏刻肋海胆和细雕刻肋海胆亲缘关系最近,与疣海胆科、毒棘海胆科、球海胆科和长海胆科的关系较远,但它们都属于海胆纲,与海星纲和海参纲的亲缘关系最远。这与Lee等(2003)应用COI和NDI得到的结论相一致,说明杂色角孔海胆与刻肋海胆属同属一个科。这一结果和曾晓起等(2012)研究中利用16S rDNA和COI基因片段进行的聚类分析结果是一致的。传统意义上的分类都认为杂色角孔海胆隶属刻肋海胆科、角孔海胆属(张凤瀛等, 1964),这与本研究综合形态学和16S rDNA基因片段系统发育分析两方面得到的结果是一致的。
表6 杂色角孔海胆的脂肪酸组成(湿重, mg/kg)

Tab.6 Fatty acid composition of S. sphaeroides variegate (Wet weight, mg/kg)
3.2 杂色角孔海胆性腺营养成分分析
杂色角孔海胆性腺中蛋白质含量为8.01%,低于光棘球海胆(13.46%)、马粪海胆(13.30%) (徐华等, 2018)和中间球海胆(9.66%~14.55%) (丁君等, 2011)、罗氏海盘车() (16.6%) (曹吉超等, 1995)和陶氏太阳海星(Verrill) (17.2%) (滕瑜等, 2016),但高于刺参(3.74%~4.43%) (韩华, 2011)和蓬莱玉参(5.25%)(刘长琳等, 2015)。杂色角孔海胆粗脂肪含量为2.0%,高于刺参(0.60%~1.00%)和蓬莱玉参(0.23%) (刘长琳等, 2015),而低于光棘球海胆(3.29±0.21)% (徐华等, 2018)、马粪海胆(3.45±0.24)% (徐华等, 2018)、罗氏海盘车(11.4%) (曹吉超等, 1995)和陶氏太阳海星(11.8%)(滕瑜等, 2016),由此可见,杂色角孔海胆的蛋白质和脂肪均高于刺参和蓬莱玉参。
杂色角孔海胆黄中的氨基酸种类齐全,共有 16种。杂色角孔海胆性腺中氨基酸中含量较高的是甘氨酸、谷氨酸和赖氨酸。氨基酸组成的重要性主要体现在营养和风味2个方面(徐华等, 2018)。本研究中杂色角孔海胆性腺中EAA/TAA为36.99%,从人类理想营养需求角度来讲,杂色角孔海胆作为蛋白来源是比较理想的,必需氨基酸占总氨酸的比例符合WHO/ FAO理想蛋白源的要求(必需氨基酸含量40%)(徐清云等, 2017),低于中间球海胆(34.01%~39.42%)、光棘球海胆(38.80%)、马粪海胆(39.91%)。EAA/NEAA为71.84%,高于FAO/WHO理想模式对EAA/NEAA的要求(60%以上),远远高于紫海胆(60.62%)、光棘球海胆(63.39%)、马粪海胆(65.50%)(徐华等, 2018)。研究表明,赖氨酸可以改善蛋白质的消化吸收,食用海胆性腺能弥补机体中的赖氨酸,从而加快人体对于食物中蛋白质的消化吸收;谷氨酸有益于改进和维持脑功能。从风味层面看,甘氨酸和丙氨酸起甜味作用,缬氨酸起苦味作用,谷氨酸起甘鲜作用。杂色角孔海胆性腺中DAA/TAA为37.17%,略高于紫海胆(36.19%)、马粪海胆(36.21%),但比光棘球海胆(40.51%)和中间球海胆(38.14%~51.84%)低。杂色角孔海胆中的谷氨酸、甘氨酸为含量最丰富的氨基酸,为海胆黄的鲜甜可口奠定基础。因此在医学和营养学上都有很高的研究价值。
丁君等(2011)发现光棘球海胆和海刺猬性腺中主要的多不饱和脂肪酸是C20:5,饱和脂肪酸以C16:0为主,杂色角孔海胆中这2种脂肪酸同样占据主导地位。杂色角孔海胆的n-3/n-6为0.79,远高于FAO/ WHO推荐的n-3/n-6日常膳食比值0.1~0.2。本研究结果发现,杂色角孔海胆中EPA和DHA的含量较高,特别是DHA的含量显著高于我国主要养殖品种中间球海胆。有研究表明,血小板磷脂中n-3/n-6比例越低,冠状动脉心脏病致死的几率越大,而EPA和DHA具有降炎的作用,杂色角孔海胆中较高的EPA和DHA含量可预防冠状动脉心脏病(Wang, 2016)。杂色角孔海胆性腺的的蛋白质含量丰富,氨基酸种类多、比例合理,脂质含量高且富含EPA和DHA等多不饱和脂肪酸,具有较高的营养价值和保健功能。
本研究通过观察杂色角孔海胆的形态,以线粒体16S rDNA基因部分序列为标记,分析其与其他棘皮动物的亲缘关系,研究结果为角孔海胆属的生物学研究提供基础。除此之外,对杂色角孔海胆的性腺营养成分进行测定分析,研究表明,海胆性腺具有较高的营养价值和保健功能,对杂色角孔海胆的综合开发利用具有很大的意义。
Cao JC, Li H, Zhang TM,. An analysis of inorganic elements, amino acids in the genital gland ofBell. Chinese Journal of Marine Drugs, 1995(1): 7–8 [曹吉超, 李华, 张天民, 等. 海星类动物的应用研究. Ⅱ罗氏海盘车生殖腺中无机元素及氨基酸分析. 中国海洋药物, 1995(1): 7–8]
Chang YQ, Ding J, Song J,. Biological research and cultivation of sea cucumber and sea urchin. Beijing: Ocean Press, 2004 [常亚青, 丁君, 宋坚, 等. 海参、海胆生物学研究与养殖. 北京: 海洋出版社, 2004]
Chen GQ , Xiang WZ , Lau CC ,. A comparative analysis of lipid and carotenoid composition of the gonads of,and. Food Chemistry, 2010, 120(4): 973–977
Ding J, Chang YQ, Hao ZL,. Comparative studies on urchin gonad fatty acid composition and β-carotene content in north China sea section. Journal of Agricultural Science and Technology, 2011, 13(3): 122–128 [丁君, 常亚青, 郝振林, 等. 北方海区经济海胆性腺脂肪酸组成与β-胡萝卜素含量的比较研究. 中国农业科技导报, 2011, 13(3): 122–128]
Fu WY. The study on microstructure of spines from four sea urchins and phylogenetic based on complete mitochondrial genome. Master’s Thesis of Chinese Ocean University, 2015, 1–84 [付婉莹. 四种海胆棘的显微结构和基于线粒体基因组全序列的系统发育学研究. 中国海洋大学硕士研究生学位论文, 2015, 1–84]
Gao TX, Li J, Wang QY,. Sequence analysis on mitochondrial 16S rDNA gene in. Journal of Fishery Sciences of China, 2003, 10(5): 359–364
Han H. Analysis and evaluation of nutritive composition in body walls for different age(Selenka). Marine Environmental Science, 2011, 30(3): 404–408 [韩华. 不同年龄刺参体壁营养成分分析及评价. 海洋环境科学, 2011, 30(3): 404–408]
Lee KZ, Sommer RJ. Operon structure and trans-splicing in the nematode. Molecular Biology and Evolution, 2003, 20(12): 2097–2103
Liu CL Wang YG, Qin B,. Analysis and evaluation of nutrient composition in the body wall of Penglai sea cucumber (sp.). Progress in Fishery Sciences, 2015, 36(5): 111–118 [刘长琳, 王有廷, 秦搏, 等. 蓬莱玉参(sp.)体壁的营养成分分析及评价. 渔业科学进展, 2015, 36(5): 111–118]
Liu CL, Zeng XQ. A comparative study on the morphological characteristics offrom different populations. Journal of Ocean University of China (Natural Science), 2015, 45(4): 40–45 [刘成龙, 曾晓起. 马粪海胆不同地区群体形态学比较研究. 中国海洋大学学报(自然科学版), 2015, 45(4): 40–45]
Liu RY. Marine life in China. Beijing: Science Press, 2008 [刘瑞玉. 中国海洋生物名录. 北京: 科学出版社, 2008]
Rahman MA, Yusoff FM, Arshad A,. Embryonic, larval, and early juvenile development of the tropical sea urchin,(Echinodermata: Echinoidea). Scientific World Journal, 2014, 2012(4): 1–9
Rahman MA, Yusoff FM, Arshad A,. Growth and survival of the tropical sea urchin,fed with different macroalgae in captive rearing condition. Journal of Environmental Biology, 2016, 37(4): 855–862
Subramaniam MK. The oogenesis of(Agassiz) with a suggestion as to the function of Golgi bodies. Proceedings of the Indian Academy of Sciences - Section B, 1934, 1(6): 291–317
Teng Y, Yuan DS, Wang DS,. Nutritional evaluation onVerrill. Agricultural Products Processing, 2016(1): 37–39 [滕瑜, 苑德顺, 王东升, 等. 陶氏太阳海星的营养评价. 农产品加工, 2016(1): 37–39]
Vinay BJ , Kanya TCS. Effect of detoxification on the functional and nutritional quality of proteins of karanja seed meal. Food Chemistry, 2008, 106(1): 77–84
Wang L, Manson JAE, Rautiainen S,. A prospective study of erythrocyte polyunsaturated fatty acid, weight gain, and risk of becoming overweight or obese in middle-aged and older women. European Journal of Nutrition, 2016, 55(2): 687– 697
Xu H, Wang YP, Yang DM,. Analysis and evaluation of nutritional components of two kinds of sea urchin gonads. Acta Nutrimenta Sinica, 2018, 40(3): 307–309 [徐华, 王云鹏, 杨德孟, 等. 两种海胆性腺营养成分分析及评价. 营养学报, 2018, 40(3): 307–309]
Xu QY, Pan N, Wu JN,. Analysis and evaluation of nutritionalcomposition of sea urchin (). Fishery Modernization, 2017, 44(2): 50–55 [徐清云, 潘南, 吴靖娜, 等. 紫海胆黄基本营养成分的分析与评价. 渔业现代化, 2017, 44(2): 50–55]
Zeng XQ, Zhang WF, Gao TX. Molecular phylogenetics of genusbased on 16S rRNA and COI sequences. Journal of Ocean University of China (Natural Science), 2012, 42(6): 47–51 [曾晓起, 张文峰, 高天翔. 基于线粒体16S rDNA与COI基因序列的刻肋海胆属系统发育研究. 中国海洋大学学报(自然科学版), 2012, 42(6): 47–51]
Zhang FY, Wu BL, Cheng LR. Animal atlas of China: Echinodermata. Beijing: Science Press, 1964, 74–101 [张凤瀛, 吴宝铃, 程丽仁. 中国动物图谱: 棘皮动物. 北京: 科学出版社, 1964, 74–101]
Zhang WF. Studies on the morphology and genetics of genetics ofand the taxonomy of temnopleuridae inshore water of China sea. Master’s Thesis of Chinese Ocean University, 2011, 50–51 [张文峰. 3种刻肋海胆的形态学与遗传学及中国刻肋海胆科分类学研究. 中国海洋大学硕士研究生学位论文, 2011, 50–51]
Zhou XW, Wang XQ, Ma X. 16S rRNA gene and its application in the classification of aquatic animals. Fisheries of Heilongjiang, 2009(2): 12–13 [周先文, 王晓清, 马晓. 16S rRNA基因及其在水产动物分类研究中的应用. 黑龙江水产, 2009(2): 12–13]
Zuo RT, Hou SQ, Chang YQ,. Research advance of nutritional physiology in sea urchins: A review. Journal of Dalian Ocean University, 2016, 31(4): 463–468 [左然涛, 侯受权, 常亚青, 等. 海胆营养生理研究进展. 大连海洋大学学报, 2016, 31(4): 463–468]
Morphological Observation, Relationship and Nutritional Composition Analysis of Gonadal of
ZHAO Wenfei, LI Zhe, HUANG Xiaofang, ZHANG Yang, DING Jun①, CHANG Yaqing①
(Key Laboratory of Mariculture & Stock Enhancement in North China's Sea, Ministry of Agriculture and Rural Affairs, College of Fisheries and Life Science, Dalian Ocean University, Dalian 116023)
In this study,was used as the research material to observe its morphology. Based on mitochondrial 16S rDNA gene partial sequencing, the genetic relationship betweenand other echinoderms was analyzed. The nutritional composition ofgonad was analyzed to accumulate a mass of data for germplasm improvement and resource evaluation of. The results showed that the main morphological characteristics of thewere as follows: the apical system was small, the outer edge of each step band plate had 3~4 small circular pits, and the bone fragment in the wall of the pedicle foot was C-shaped. The mitochondrial 16S rDNA gene sequences of other echinoderms and that obtained fromwere compared by Mega 5.0, and the phylogenetic trees were constructed based on distance method (NJ) and maximum likelihood method (MP). The results showed that thewas closely related to the, followed by the three species of sea urchins, the Temnopleuridae. An amount of water 86.1%, ash 3.0%, crude fat 2.0% and protein 8.01% was found in the sex glands of. There were 16 kinds of amino acids in the sex glands of, the total amino acid content was 38.71 g/100 g, EAA/TAA was 36.99%, EAA/NEAA was 71.84%, and DAA/TAA was 37.17%. A total of 15 fatty acids were detected, with the highest content being palmitic acid C16:0, stearic acid C18:0, EPA C20:5 and arachidonic acid C20:4, and the lowest content was that of C20:3.showed a higher content of DHA than in other sea urchins. The results will provide basic data for the biological study ofand can be used for the development and application of sea urchin ofeconomic value, as well as provide a theoretical basis for distant hybridization breeding and the germplasm identification of economically viable sea urchins.
; Morphology; Gonadal nutrition; Mitochondria; 16S rDNA
S917.4
A
2095-9869(2019)06-0163-10
10.19663/j.issn2095-9869.20190220001
http://www.yykxjz.cn/
赵文飞, 李喆, 黄小芳, 张杨, 丁君, 常亚青. 杂色角孔海胆形态学观察、亲缘关系及性腺营养成分分析. 渔业科学进展, 2019, 40(6): 163–172
Zhao WF, Li Z, Huang XF, Zhang Y, Ding J, Chang YQ. Morphological observation, relationship and nutritional composition analysis of gonadal of. Progress in Fishery Sciences, 2019, 40(6): 163–172
* 国家重点研发计划项目(2018YFD0901601)资助[This work was supported by the National Key Research and Development Program of China (2018YFD0901601)].赵文飞,E-mail: 2895741046@qq.com
丁 君,研究员,E-mail: dingjun1119@dlou.edu.cn; 常亚青,教授,E-mail: yqchang@dlou.edu.cn
2019-02-20,
2019-04-25
DING Jun, E-mail: dingjun1119@dlou.edu.cn; CHANG Yaqing, E-mail: yqchang@dlou.edu.cn
(编辑 陈辉)
