蔬菜-牧草轮作5年草地土壤微生物量变化及其群落结构分异
2019-11-18林栋张德罡McCulleyRebecca
林栋,张德罡,McCulley Rebecca L.
(1.甘肃农业大学草业学院,甘肃 兰州 730070;2.美国肯塔基大学植物与土壤科学系,肯塔基州 列克星敦 40546-0091)
作物长期连作会导致土壤质量下降,营养元素失衡,自毒物质和病原微生物积累[1],尤其在蔬菜类作物连作中更为常见[2]。而蔬菜与牧草轮作能够有效缓解连作障碍,改善土壤理化性状和微生态环境,提高农业生态系统的生产力[3]。采用轮作种植模式是全球温带地区可持续农业重要的土地利用策略之一[4-5],该轮作方式在全球范围内被证实有诸多好处[6],如增加土壤有机质和生物量,形成潜在的土壤碳汇,减缓气候变化,对实现农牧业可持续生产具有重要意义,是集生态效益、经济效益、社会效益于一体的草地农业生产模式,具有广泛的推广应用前景[7]。Conant等[8]对全球115项有关耕地或原生植被转变为草地,以及提高草地管理措施的研究结果进行了统计分析,结果表明耕地转变为草地显著增加了土壤碳沉积。Franzluebbers[9]在美国东南部对作物-家畜系统的研究表明,被侵蚀的农田种植多年生牧草后土壤有机质、土壤营养循环、生物和物理状况显著提升。中国春秋战国时期伴随着土地连作制度的出现,轮作就在古代农耕制度中逐渐形成,雏形是作物和绿肥轮作[10],目前已广泛应用于各地区农牧业生产和生态环境保护。李静等[11]对黄土区不同退耕年限草地土壤理化性质的研究结果表明,随着退耕年限的增加,草地土壤分离速率逐渐降低,不同退耕年限草地土壤容重、水稳性团聚体含量、根系生物量密度均有不同程度的改变。
土壤微生物直接参与土壤有机质积累和矿化等过程,影响土壤养分循环及其有效性,调节和指示土壤功能[12],是反映生态系统受到一定干扰后土壤质量变化的指标之一[13-15]。土壤微生物生物量比土壤有机质变化更敏感,其群落结构能很好地区分不同草地生长年限土壤有机质存在的差异。土壤微生物对环境扰动的响应非常迅速,由于能够较早地预测土壤养分及环境质量的变化过程,被认为是最具潜力的敏感性指标之一[16]。传统耕作措施下的蔬菜生产,被证实会减少土壤微生物生物量[17-18]和真菌种群数量[19-20]。而放牧草地土壤相比蔬菜农田,能够维持更高的微生物生物量和新陈代谢潜力[14]。同时,研究表明草地土壤由于较高的微生物群落丰富度和多样性,而更能抵抗环境胁迫[21]。生态系统忍耐外界干扰的能力,部分依赖于土壤微生物组成[22]。定量分析土壤微生物生物量和群落结构,在探究土壤营养物质循环和积累过程中至关重要,然而在蔬菜-牧草轮作系统中,草地生长年际和季节间的变化规律尚缺乏系统和精确的分析结论。研究蔬菜-牧草轮作过程中牧草生长时间序列上土壤微生物群落结构与功能动态特征,有助于深入理解该轮作措施影响土壤状况和功能发挥的生物学机制。
1 材料与方法
1.1 研究区概况
研究区位于美国肯塔基州乔治敦县的埃尔姆伍德有机畜牧场(N 38°13′26″-38°14′00″、W 84°30′08″-84°30′53″),海拔165~323 m,处于亚热带湿润气候区的北缘[23],年均降水量1209 mm,年均气温12 ℃,无霜期163~192 d。该有机牧场不使用化肥和除草剂等农药,一直沿用3年低投入蔬菜生产和5年多年生牧草轮作的生产方式。研究地坡度约2%~6%,土壤类型为淋溶土(国际土壤分类系统),采样点土壤类型、坡度、朝向、植被总盖度等基本立地条件保持一致[24]。
1.2 试验设计
运用空间代替时间的方法,选取蔬菜-牧草轮作系统中处于不同建植年限的多年生草地作为研究对象,以永久性草地为对照,分析连续5年牧草轮作过程中不同生长年限和不同季节之间草地土壤微生物群落动态特征。研究区轮作草地前茬蔬菜作物种类一致,主要为:甘蓝(Brassicaoleracea)、甜菜(Betavulgaris)和胡萝卜(Daucuscarota)等。多年生草地主要物种为:苜蓿(Medicagosativa)、草地早熟禾(Poapratensis)、高羊茅(Festucaarundinacea)、鸭茅(Dactylisglomerata)、白三叶(Trifoliumrepens)、红三叶(Trifoliumpratense)和梯牧草(Phleumpratense)。轮作草地生长年限从第1~5年的处理,分别标记为Y1、Y2、Y3、Y4、Y5,近50年未曾耕种过的永久性草地(permanent pasture)标记为P。从每个相同生长年限的草地中随机选取立地条件基本一致的3个小区(5 m×5 m)作为重复,每个小区内以“S”形布置6个采样点,于2015年4个季节(春、夏、秋、冬)分别采集0~15 cm土层土壤,6土钻混合为一袋样品,塑料自封袋封装后用冰盒带回实验室,过5.66 mm筛,剔除可见植物根茎组织后进行检测。
1.3 高通量磷脂脂肪酸分析
运用改进后的Buyer等[25]的方法进行高通量磷脂脂肪酸(phospholipid fatty acid, PLFA)分析,检测研究区草地土壤微生物群落。将新鲜土样冷冻干燥后取1.0~1.5 g在玻璃试管中溶于4 mL的Bligh-Dyer提取剂(氯仿∶甲醇∶0.05 mol·L-1磷酸钾缓冲液,体积比1.0∶2.0∶0.8,pH 7.4),含内标(19∶0, 1, 2-dinonadecanoyl-sn-glycero-3-phosphocholine)0.5 μL·mL-1,测试组前中后分别加入3个空白对照。通过超声波降解,加入氯仿和去离子水涡旋振荡,并离心浓缩进行分离。用1 mL甲醇淋洗50 mg硅胶固相萃取(solid phase extraction,SPE)96孔板(Phenomenex,Torrance,美国)3次,再用1 mL氯仿淋洗3次。加入分离出的样品后,用1 mL氯仿和1 mL丙酮间隔5 min淋洗,再用0.5 mL甲醇∶氯仿∶水为5∶5∶1的溶剂洗提磷脂。脂肪酸甲酯(fatty acid methyl esters,FAMEs)提取用0.2 mL酯交换反应试剂(0.285 g KOH溶于25 mL甲醇后加8.3 mL甲苯),在39 ℃下培养15 min进行甲基化。加入0.4 mL 0.075 mol·L-1醋酸、0.4 mL氯仿,反复振荡分离后在真空离心浓缩系统中干燥30 min提取FAMEs,再用75 μL正己烷溶解。用搭载有自动进样器、Ultra 2毛细管柱、火焰离子化检测器和MIDI Sherlock系统(Microbial Identification System Inc.,版本6.2, Newark, DE)的安捷伦7890A气相色谱仪(Agilent Technologies,Wilmington,美国)测定脂肪酸种类和含量。脂肪酸命名参照Vestal等[26]和Frostegård等[27]的方法,通过MIDI系统微生物特征PLFA谱图数据库比对,参考已被广泛证实的微生物标志脂肪酸,并结合研究区同类土壤微生物区系分析,用特定FAMEs指示不同微生物类群。
1.4 数据统计
草地生长年限、季节及其交互效应对土壤中不同种类FAMEs含量、特定脂肪酸比例和微生物生物量的影响,用JMP统计分析软件(版本12.0, SAS软件研究所)进行双因素方差分析。草地不同生长年限和季节间土壤微生物群落结构的分异(或相似性),用多元统计分析软件PC-ORD 6.0(MjM软件,美国)进行判别,PLFA提取物中相对丰度>1%的FAMEs(n=22)用于非度量多维测度(nonmetric multidimensional scaling,NMS)排序。用Sørensen距离(Bray-Curtis法计算[28])确定样本间非线性梯度的相关性及方向,并将草地不同生长年限和季节土壤微生物群落结构差异转化为可视化的NMS排序图。模型不稳定性判别标准0.00001,步长为0.2,随机运行250次。同时,分析驱动微生物群落结构分异的主要环境因子。环境变量在NMS排序中显示为矢量图,箭头的长度和角度分别表示相关性强度和方向。多响应置换过程(multi-response permutation procedure,MRPP)是一种非参数多元分析方法,用于判别不同生长年限和季节间土壤各类FAMEs丰度比例的两两比较和组间差异显著性。多重比较的差异显著性P值进行Bonferroni校正,不同生长年限间15组,季节间6组。
2 结果与分析
2.1 不同年限草地土壤中FAMEs分布特征
从不同生长年限草地和永久性草地土壤中,共检测出C10到C24的98种FAMEs,其中草地生长第5年土壤中FAMEs种类最多,为92种,比草地建植初期高出19%,永久性草地土壤检测到85种FAMEs。脂肪酸10:0 3OH和13:1ω4c在草地生长第4和第5年土壤中出现,19:1ω6c、19:0cyclo ω9c、20:2ω6c、24:1ω9c和16:2DMA 5种脂肪酸仅在第5年草地土壤中出现,而14:1ω7c仅在永久性草地土壤中检测到。不同季节有部分特征脂肪酸出现,例如:10:0(癸酸)、14:1ω9c、19:0cyclo ω9c仅在春季土样中检测到,24:1ω9c仅出现在夏季土样中,16:3ω6c和16:2DMA仅在秋季土样中检测到,13:1ω4c和14:1ω7c仅在冬季土样中出现。夏季、秋季和冬季草地土壤中均检测到11:0、12:0DMA和19:1ω6c,而在春季土壤中未检测到。10:0 3OH、20:0iso、20:2ω6c、22:6ω3c和22:4ω6c 5种脂肪酸在春、夏、冬季草地土壤中均有分布,而在秋季土壤中消失。春、夏、秋季草地土壤中均含有10:0 2OH,冬季土壤中却未检测到。
随着草地生长年限的增加,土壤中各类主要脂肪酸比例也随之发生变化(表1)。草地生长年限对饱和脂肪酸与单一不饱和脂肪酸之比(saturated/monounsaturated,sat/mono)、革兰氏阳性菌与革兰氏阴性菌脂肪酸的比例(gram positive/gram negative,G+/G-)产生极显著影响(P<0.01),而对环丙烷脂肪酸与其前体脂肪酸之比(cyclopropane/precursors,cy/pre)无显著影响(P=0.1188)。永久性草地土壤sat/mono低于生长1~5年草地,第2年草地亦低于第1和4年草地。随草地生长年限的增加,G+/G-有逐渐减小的趋势,第5年草地G+/G-显著低于第1和3年,比草地建植初期降低8%,接近永久性草地土壤G+/G-。草地生长5年间异构与反异构脂肪酸之比(isomers/ante-isomers,i/a)无显著差异(P>0.05),但除第3年草地外,其他年限草地均显著高于永久性草地(P<0.01)。真菌脂肪酸与细菌脂肪酸的比例(fungi/bacteria,F/B),在不同生长年限草地土壤中也表现出显著差异(P<0.05),Y5比Y1、Y2、Y3、Y4均高13%,且与永久性草地土壤F/B无显著差异(P=0.6561)。
季节显著影响研究区草地土壤中sat/mono、cy/pre、i/a和G+/G-(P<0.01),sat/mono夏、秋季比春、冬季分别高出20%和27%,cy/pre和i/a夏、秋季亦显著高于春、冬季,G+/G-在各季节草地土壤中的高低顺序为秋季>夏季>冬季≈春季。F/B在不同季节草地土壤中,也表现出显著差异(P<0.05),秋季F/B均比春、夏、冬季低13%(表2)。
草地生长年限与季节对sat/mono(P=0.1215)、cy/pre(P=0.2802)、i/a(P=0.9663)、F/B(P=0.0506)和G+/G-(P=0.4700)的交互效应均不显著。
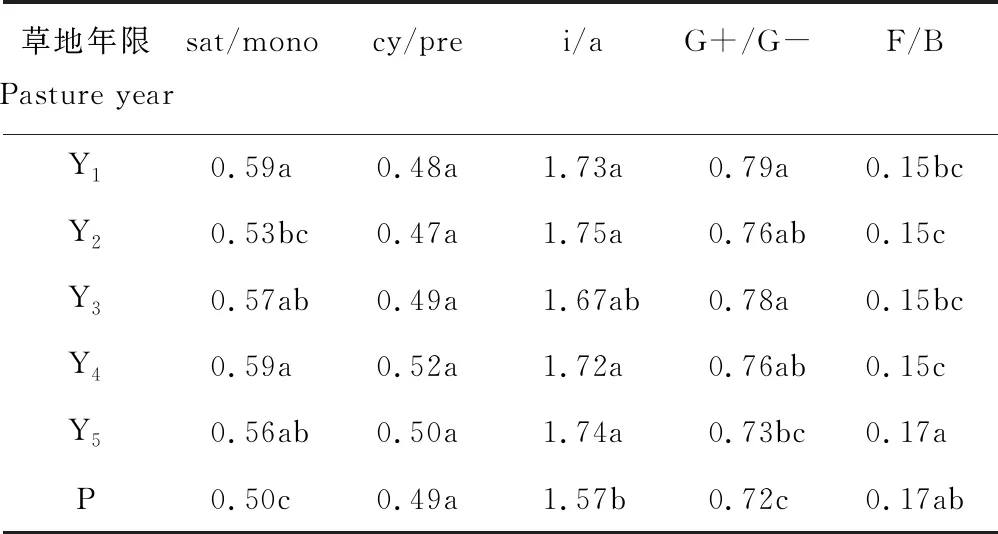
表1 不同生长年限草地土壤特定脂肪酸比例Table 1 Special FAMEs ratio of pasture soils in different growing years
注:sat/mono,饱和脂肪酸/单一不饱和脂肪酸;cy/pre,环丙烷脂肪酸/前体脂肪酸;i/a,异构脂肪酸/反异构脂肪酸;G+/G-,革兰氏阳性菌脂肪酸/革兰氏阴性菌脂肪酸;F/B,真菌脂肪酸/细菌脂肪酸;Y1~Y5,生长1~5年草地;P,永久性草地;不同小写字母表示在0.05水平上处理间差异显著,下同。
Note: sat/mono, ratio of saturated to monounsaturated fatty acids; cy/pre, ratio of cyclopropane to precursors fatty acid; i/a, ratio of isomers to ante-isomers fatty acid; G+/G-, ratio of gram positive to negative bacteria; F/B, ratio of fungi to bacteria; Y1-Y5, pastures 1 to 5 years old; P, permanent pasture. Different lowercase letters indicate significant differences between treatments (P=0.05), the same below.
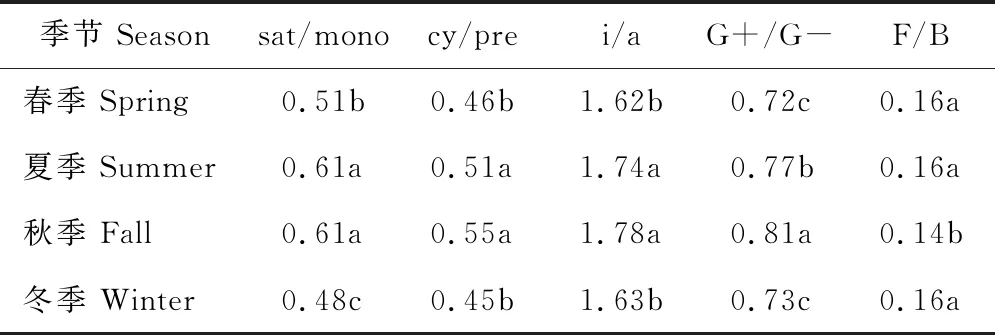
表2 不同季节草地土壤特定脂肪酸比例Table 2 Special FAMEs ratio in pasture soil in different seasons
2.2 不同年限草地土壤微生物量变化
草地生长年限显著影响土壤细菌、真菌、真核生物、原生动物及其总微生物生物量(P<0.01),随着草地生长年限的增加,土壤总微生物量逐年增加,从Y1到Y5增加了46 nmol·g-1土壤,增长了45%,Y5接近永久性草地土壤微生物总量(表3)。细菌和真菌生物量经过草地生长5年分别增长了44%和63%,变化趋势与土壤总微生物量相似,Y2均高于Y1、Y3和Y4,且与Y5和永久性草地土壤微生物量之间无显著差异(P>0.05)。真核生物生物量5年增加69%,Y5高于其他年份草地和永久性草地土壤。原生动物生物量在草地生长前4年无显著差异(P>0.05),生长5年后增加1倍,且显著高于生长1~4年草地和永久性草地(P<0.05)。
季节显著影响草地土壤中真核生物(P=0.0327)和原生动物(P<0.01)生物量的变化(图1)。春季草地土壤中真核生物生物量为1.8 nmol·g-1,比秋季草地土壤高20%。原生动物生物量在夏季(1.2 nmol·g-1)高于其他3个季节,且存在显著的草地生长年限×季节的交互效应(P<0.01),生长第5年草地在夏季的土壤原生动物生物量最高。

表3 不同生长年限草地土壤微生物生物量Table 3 PLFA concentrations of the major microbial groups in pasture soils in different growing years (nmol FAMEs·g-1 soil)
注:表中数据为平均值±标准误差;Y,草地生长年限;S,季节;Y×S,草地生长年限和季节的交互效应。**表示P<0.01,*表示P<0.05,NS表示不显著,下同。
Note: Mean±SE; Y, pasture growing year; S, season; Y×S, pasture growing year and season interaction. ** indicates significant difference atP<0.01, * indicates significant difference atP<0.05, NS indicates no significance, the same below.
由表4可知,草地生长5年,土壤细菌中革兰氏阳性菌(G+)和革兰氏阴性菌(G-)生物量分别增加了38%和49%。Y5草地土壤G+和G-生物量大于Y1、Y3和Y4,与Y2和永久性草地无显著差异。Y5草地土壤中的厌氧菌含量高于所有其他年限草地和永久性草地,比草地建植第1年增加67%,但与Y2和永久性草地的差异并不显著。放线菌在永久性草地土壤中含量最高,生长5年增加38%,Y4与Y5之间无显著差异。真菌中腐生真菌和丛枝菌根真菌经过草地生长5年,分别增加了75%和41%,并接近永久性草地含量。
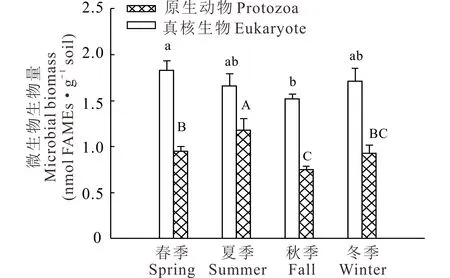
图1 不同季节草地土壤真核生物和原生动物生物量Fig.1 Microbial biomass of eukaryote and protozoa in pasture soil in different seasons 不同小写字母表示真核生物生物量在季节间存在显著差异(P<0.05),不同大写字母表示原生动物生物量在季节间存在显著差异(P<0.05)。Different lowercase letters indicate significant difference of eukaryote biomass between seasons while different capital letters indicate significant differences of protozoa biomass between seasons (P<0.05).
研究区草地土壤中的厌氧菌和腐生真菌生物量在不同季节间存在显著差异(图2),但草地生长年限×季节的交互效应均不显著(表4)。冬季草地土壤厌氧菌平均含量1.8 nmol·g-1,显著高于秋季(1.5 nmol·g-1)和夏季(1.4 nmol·g-1),春季亦比夏季高24%。腐生真菌生物量在秋季草地土壤中平均为7.7 nmol·g-1,比春季、夏季和冬季(平均9.7 nmol·g-1)低21%。
2.3 不同年限草地土壤微生物群落结构分异
用相对丰度>1%的22种FAMEs进行非度量多维测度(NMS)排序,其结果揭示了微生物群类在草地不同生长年限和季节间的分异。对不同生长年限草地土壤微生物结构进行NMS排序(图3),生长第5年草地和永久性草地土壤微生物结构与生长第1、2、3年草地分离,排序轴1和2累计解释了82.9%的变异。多响应置换过程(MRPP)分析显示,Y1、Y2、Y3与Y5、P差异显著(P<0.05),而Y1、Y2、Y3、Y4之间无显著差异(P>0.05),Y4、Y5、P之间差异亦不显著(P>0.05)。可见,草地生长4年改变了土壤微生物群落结构,并趋近于永久性草地土壤微生物群落结构。进一步用PC-ORD识别驱动土壤微生物群落结构在草地生长年际间分异的环境因子,环境变量包括土壤有机碳氮、颗粒与非颗粒有机质、碳矿化率、Ca、P、K、Mg、Zn元素含量及其交换量、阳离子交换量等,结果表明土壤碳矿化(carbon mineralization,CM)和颗粒有机质(particulate organic matter,POM)含量是主要驱动因子。

表4 不同生长年限草地土壤细菌和真菌生物量Table 4 PLFA concentrations of bacteria and fungi in pasture soils in different growing years (nmol FAMEs·g-1 soil)
对不同季节草地土壤微生物群落结构NMS排序(图4)结果表明,秋季草地土壤微生物群落结构与春季、夏季、冬季草地土壤微生物群落结构明显分离(P<0.01),排序轴1和2累计解释82.9%的变异。同时,MRPP分析结果显示,夏季草地土壤微生物群落结构与冬季草地土壤微生物群落结构差异显著(P<0.01),而春季与夏季(P=0.1003)、冬季(P=0.3664)差异均不显著。

图2 不同季节草地土壤厌氧菌和腐生真菌生物量Fig. 2 Microbial biomass of anaerobe and saprophytic fungi in pasture soil in different seasons 不同小写字母表示厌氧菌生物量在季节间存在显著差异(P<0.01),不同大写字母表示腐生真菌生物量在季节间存在显著差异(P<0.05)。Different lowercase letters indicate significant difference of anaerobe biomass between seasons (P<0.01) while different capital letters indicate significant differences of saprophytic fungi biomass between seasons (P<0.05).
对各季节草地土壤中不同种类FAMEs含量的方差分析表明,秋季草地土壤含有较丰富的G+、G-和放线菌的脂类标志物,例如:15:0iso、15:0anteiso、19:0cycloω7c和16:0 10-methyl等。而在冬季草地土壤中FAMEs类群向丛枝菌根真菌和放线菌生物标记转变,如16:0 10-methyl和16:1ω5c。腐生真菌(18:1ω9c)和原生动物(20:4ω6c)生物标记在夏季草地土壤中显著影响微生物群落,而在春季草地土壤中G-标志物(16:1ω7c、18:1ω7c等)含量显著增加。
3 讨论
3.1 草地生长年限和季节对土壤中FAMEs的影响
从检测到的FAMEs种类可以看出,轮作系统中随着牧草生长年限的增加,FAMEs种类不断增加,表明土壤微生物多样性随牧草种植年限的延长有提高的趋势,但永久性草地土壤FAMEs种类数低于生长5年的草地。白丽等[29]对黄土高原草地次生演替的研究表明,前30年土壤微生物多样性逐渐增大,之后缓慢下降。长期单一的土地利用方式相比草地轮作降低了土壤微生物多样性,此结果与张笑宇等[30]对烟草(Nicotianatabacum)轮作的研究结果一致,同时符合中度干扰假说[31]的生态学规律。
不同土地利用方式的土壤具有特定的脂肪酸类群和微生物群落结构[32],Acosta-Martínez等[14]报道了放牧草地与蔬菜农田相比,真菌PLFA标志物增加(例如18:2ω6c、18:1ω9c、16:1ω5c),造成了土壤微生物群落结构的差异。蔬菜轮作为牧草4年后,土壤中陆续出现一些新的脂肪酸种类,例如表征革兰氏阴性菌的10:0 3OH、13:1ω4c、19:1ω6c、19:0cyclo ω9c、24:1ω9c,表征厌氧菌的16:2DMA和表征真核生物的20:2ω6c,可以指示草地生长年限增加过程中土壤微生物的发生学关系,说明部分脂肪酸种类更适宜于较长生长年限的草地土壤微环境。研究表明[33],20:4ω6c与土壤酶活性和土壤微生物量密切相关。永久性草地土壤饱和脂肪酸与单一不饱和脂肪酸之比显著降低,可能是土壤有机质含量较高的结果。Bossio等[34]的研究表明,在养分充足的土壤环境中,不饱和脂肪酸的比例会更高。

图3 不同生长年限草地土壤微生物群落结构非度量多维测度排序Fig. 3 PC-ORD nonmetric multidimensional scaling (NMS) ordination biplots using relative abundance of FAMEs>1% in pasture soil in different growing years Y1~Y5:生长1~5年草地 Pastures 1 to 5 years old;P:永久性草地 Permanent pasture;POM:土壤颗粒有机质 Particulate organic matter;CM:土壤碳矿化 Carbon mineralization.

图4 不同季节草地土壤微生物群落结构非度量多维测度排序Fig. 4 NMS ordination biplots using relative abundance of FAMEs>1% in pasture soil in different seasons Sp:春季 Spring;Su:夏季 Summer;F:秋季 Fall;W:冬季 Winter.
不同季节草地土壤中出现了特异脂肪酸,或有部分脂肪酸在特定季节消失,可能是土壤微生物对季节间水热配比状况差异的响应。不同生长年限草地土壤在夏秋季饱和脂肪酸与单一不饱和脂肪酸之比显著高于春冬季,Moore-Kucera等[35]的研究结果也表明,夏季干旱胁迫引起饱和与不饱和脂肪酸比例升高。特定脂肪酸比例变化可以表征土壤微生物对环境胁迫的应答,但前提条件是环境压力导致了细菌细胞膜结构组成的改变[36]。
3.2 草地生长年限和季节对土壤微生物量的影响
蔬菜-牧草轮作系统中,随着草地生长年限的增加,土壤微生物量显著增加,并接近永久性草地土壤微生物含量。Acosta-Martínez等[33]在德州西部的研究表明,多年生牧草和黑麦(Secalecereale)-棉花(Gossypiumhirsutum)轮作土壤微生物量高于棉花连作。徐丽君等[37]对不同生长年限紫花苜蓿人工草地的研究结果表明,1~4年土壤微生物量逐渐增加,第5年开始减少。不同生长年限草地土壤的微生物总量、细菌、真菌和放线菌生物量在季节间无显著差异,暗示季节并不影响土壤中占比较大的微生物类群的生物量,大部分微生物量在季节间保持基本稳定。然而,占比较小的真核生物、原生动物、腐生真菌、厌氧菌生物量在不同季节间有显著差异。李振高等[38]和Bell等[39]指出,土壤温度和水分的季节变化直接控制着生态系统中微生物量和酶活性波动,土壤微生物量碳与土壤湿度、温度、降水和平均气温等非生物因素具有显著正相关关系。同时,土壤微生物量季节性涨落还与土壤中可利用碳和养分资源的限制、植物生长节律有关[40]。
3.3 草地生长年限和季节对土壤微生物群落结构和功能的影响
牧草种植5年土壤真菌与细菌比率显著提高,指示土壤功能发生重要变化[21]。研究表明,土壤微生物真菌途径对碳的存储能力较强[41],部分ε-变形菌属的细菌通过分泌特异性酶影响CO2固定能力[42]。真菌菌丝被认为是土壤中不稳定性碳和营养物质的重要来源,在细菌接近颗粒碳底物的过程中发挥着关键作用[43]。草地生长5年,丛枝菌根真菌含量增加,且在永久性草地土壤中最多。Bainard等[44]在加拿大草原的研究表明,对于长期放牧草地,丛枝菌根真菌能增强土壤氮和磷的吸收与转化,提高牧草耐受干旱胁迫的能力,促进土壤团聚体的形成和稳定性。随着草地生长年限的延长,革兰氏阳性菌占比逐渐减小,而革兰氏阴性菌比例增大。Romaniuk等[45]也证实革兰氏阴性细菌在生长10年的放牧草地土壤中较多,革兰氏阴性菌含量常随着土壤有机碳有效性的增强而增加[46]。Riah-Anglet等[21]对土壤微生物群落结构与功能的关系研究表明,永久性草地土壤微生物群落结构的改变与功能的变化紧密关联。草地生长第2年土壤微生物生物量显著增加,但群落结构在草地生长4年才发生显著变化,说明量变积累到一定阶段后才会发生结构性的质变。Xue等[47]对不同土地利用系统的研究也发现,微生物生物量的变化不一定总是伴随着微生物群落多样性和活性的变化。
季节变化影响草地土壤微生物群落[48],不同生长年限草地土壤微生物群落结构在秋季显著区别于其他季节,夏季与冬季不同。Slaughter等[49]在本试验区附近的草地研究结果也表明,季节显著影响草地土壤微生物群落结构。Stevenson等[48]对新西兰北岛温带气候区草地的研究也表明,秋季土壤细菌群落明显区别于其他季节,本研究结果与此一致。季节间土壤微生物群落结构的变化与非生物因素有关,例如土壤温度、湿度及其综合作用等。土壤微生物群落结构变化导致其分解功能稳定性发生改变[50],本研究中草地土壤微生物群落结构秋季不同于其他季节,秋季土壤碳矿化潜力也显著低于其他季节。
4 结论
随着蔬菜-牧草轮作系统中草地生长年限的增加,土壤微生物的脂肪酸种类和数量逐渐增加。草地生长4年后土壤中出现新的脂肪酸种类,不同季节亦有特征脂肪酸出现或消失。草地生长5年土壤微生物总生物量增加45%,接近永久性草地土壤微生物生物量。其中原生动物生物量增加1倍,腐生真菌增加75%。季节显著影响真核生物、原生动物、腐生真菌、厌氧菌生物量和特定脂肪酸比例。草地生长4年后土壤微生物群落结构发生显著改变,且趋近于永久性草地土壤微生物群落结构。土壤碳矿化和颗粒有机质含量的变化是驱动土壤微生物群落结构分异的主要环境因子。
