INPP5E基因影响小鼠胚胎神经管闭合的分子机制研究*
2019-10-24秦佳星王秀伟官臻牛勃郭金王建华钟儒刚
,秦佳星 ,王秀伟 ,官臻 ,牛勃,郭金,王建华,钟儒刚
(1.北京工业大学 环境与病毒肿瘤学北京市重点实验室,北京 100124;2.首都儿科研究所 转化医学研究室,北京 100020)
肌醇多聚磷酸5-磷酸酶(INPP5E)基因编码INPP5E蛋白,该基因敲除后小鼠出现无脑和露脑的神经管缺陷(neural tube defects,NTD)表型[1-3]。NTD为胚胎中枢神经系统先天畸形,是环境因素与遗传因素复杂交互作用的结果,环境因素包括母体营养、地理及社会经济状况等,而母体叶酸(folic acid,FA)与NTD的发生密切相关[4-8]。FA在体内以四氢叶酸的形式参与一碳单位的代谢,与DNA甲基化过程等表观遗传修饰相关。本研究通过FA缺乏NTD小鼠模型,研究该基因影响胚胎神经管闭合的分子机制,为NTD的防治提供实验依据。
1 材料与方法
1.1 材料
1.1.1 动物7~8 周体重 18~20 g 的无特定病原体级C57BL/6J成年健康雌鼠20只(北京维通利华实验动物技术有限公司)。在20~24℃恒温、40%~60%恒湿的条件下,鼠繁殖饲料喂养(北京维通利华实验动物技术有限公司),常规饮水,自由取食。动物房中不定时提供食物和水。本研究已获得首都儿科研究所实验动物伦理委员会的批准(批准号:DWLL2016004)。
1.1.2 试剂和仪器兔源甘油醛 -3- 磷酸脱氢酶(glyceraldehyde-3-phosphate dehydrogenase,GAPDH)、INPP5E单克隆抗体(美国Abcam公司),蛋白上样缓冲液(上海碧云天生物技术公司),脱脂奶粉(美国BD Difco 公司),预染 Marker(美国 Thermo Scientific公司),30%聚丙烯酰胺(上海碧云天生物技术公司),一步法动物组织活性蛋白提取试剂盒(上海生工生物工程股份有限公司),DNA提取纯化试剂盒(德国QIAGEN公司),DNA修饰试剂盒(美国Chemicon公司)。EG1150石蜡包埋机(德国Leica公司),Model200/2.0蛋白质电泳仪(美国Bio-Rad公司),221BR Trans-Blot SD Semi-Dry Transfer Cell(美国 Bio-Rad 公司),Image Quant LAS 4000mini(美国General Electric 公司),酶标仪(美国 Perkin Elmer公司),7500实时荧光定量聚合酶链反应(quantitative real-time polymerase chain reaction,qRT-PCR)仪(美国Applied Biosystems公司),多用凝胶成像分析系统(美国Bio-Rad公司),超高效液相色谱-串联四极杆质谱联用仪(美国Waters公司)等。
1.2 方法
1.2.1 动物模型及标本采集雌鼠体重达 20 g 后,与雄鼠合笼过夜(18:00至次日6:00),次日观察是否出现阴栓,出现阴栓可认为12:00时为胚胎发育第0.5天。
取16只孕鼠,在胚胎发育第7.5天时随机分为对照组和NTD组进行处理,每组8只。对照组腹腔注射0.9%氯化钠NaCl,NTD组采用腹腔注射4.5 mg/kg甲氨蝶呤,干扰FA代谢从而复制NTD小鼠模型[9-10]。在胚胎发育第11.5天颈部脱臼处死小鼠,取胚胎,体视显微镜下观察胚胎畸形情况。
1.2.2 苏木精 - 伊 红(hematoxylin-eosin staining,HE)染色 将胚胎固定于100 g/L 甲醛溶液中,在乙醇溶液中脱水,在二甲苯溶液中浸泡,之后放入石蜡嵌入包埋,将石蜡块用组织切片机进行神经管组织部位切片,厚度为5 mm,将切片放入二甲苯和乙醇溶液中脱蜡,随后放入苏木精染色液中约10 min,水冲洗后用伊红液浸染3 min,再置于水中洗去浮色。
1.2.3 超高效液相色谱串联质谱法取 20 ml小鼠血浆,加入1.0 g活性炭,冰上轻柔搅拌1 h,4℃、20 000 r/min 离心 5 min,上清为无 FA 小鼠血浆结合酶,分装储存于-70℃备用。第11.5天用提取缓冲液对对照组及NTD组小鼠胚胎神经组织样品进行匀浆。100℃加热提取 15 min,20 000 r/min 离心 15 min,在上清中加入血浆结合酶,将多聚谷氨酸FA分解为小分子的单谷氨酸FA,37℃孵育1.5 h后100℃加热5 min,20 000 r/min 离心 20 min,用超高效液相色谱串联质谱法检测上清液。
1.2.4 重亚硫酸盐测序法在两组小鼠胚胎发育第11.5天采用重亚硫酸盐测序法检测胚胎神经组织中INPP5E基因启动子区DNA甲基化水平。使用DNA提取纯化试剂盒提取基因组DNA于-20℃保存。NTD组DNA采用DNA修饰试剂盒进行重亚硫酸盐处理,将NTD组中未发生甲基化的胞嘧啶(C)转变成尿嘧啶(U),PCR扩增后转化为胸腺嘧啶(T),反之发生甲基化的C无变化。利用UCSC在线查询INPP5E基因 CpG 岛,使用 Methyl Primer Express V1.0软件设计重亚硫酸盐测序法引物,正向引物:5’-GGTTTAGTTATTGGGAAGGAGT-3’,反向引物:5’-AAATTAAACAAAATTAAATCCACCAAA-3’。 扩增产物长度为328 bp,共检测该区域8个CpG甲基化位点。PCR反应体系为50μl,反应条件:95℃预变性 10 min,95℃变性 30 s,55℃退火 30 s,72℃延伸30 s,共40个循环,72℃继续延伸7 min。PCR产物经2%琼脂糖凝胶电泳,在300 mm紫外灯下观察结果,凝胶成像分析系统记录处理数据,纯化后经Sanger法测序,使用BiQ Analyzer软件进行分析,将C信号的峰高与C+T信号的峰高进行比较,来定量每个CpG二核苷酸位点中的甲基化水平。
1.2.5 Westernblotting 通过 Western blotting 检测INPP5E蛋白的相对表达量。在胚胎发育第11.5天对发育正常小鼠胚胎及发育缺陷小鼠胚胎进行解剖,收集神经组织,用CelLytic MT蛋白裂解液提取正常胚胎神经组织总蛋白,使用布拉德福德光谱蛋白质法测蛋白含量。对蛋白进行SDS-PAGE后取下凝胶,进行电转移,电转移后的聚偏二氟乙烯膜(polyvinylidene fluoride,PVDF)在10%脱脂牛奶中室温轻摇,封闭1.5 h。将PVDF放入玻璃皿内,加入一定体积稀释的I抗(1∶1 000),4℃过夜。之后用抗鼠II抗(1∶2 000)室温孵育2 h。用增强化学发光液ECL显色,使用影像处理软件分析,GAPDH作为内参蛋白。
1.2.6 qRT-PCR采用qRT-PCR检测两组胚胎发育第11.5天小鼠胚胎神经组织中INPP5EmRNA的相对表达量。20μl PCR 反应液 :SYBR® Premix DimerEraser™(2×)10μl、PCR 正向引物(10μmol)0.6μl、PCR 反 向 引 物(10μmol)0.6μl、ROX 参比染料(50×)0.4μl、模板 2μl、ddH2O 6.4μl,采用序列分析和熔解曲线分析对PCR产物的特异性进行检验。反应条件:95℃预变性30 s,95℃变性3 s,55℃退火 30 s,72℃延伸 32 s,共 40个循环,72℃继续延伸 34 s。
1.3 统计学方法
数据分析采用SPSS 23.0软件。计量资料以均数±标准差(±s)表示,比较用t检验;计数资料以率(%)表示,比较用χ2检验或Fisher确切概率法,P<0.05为差异有统计学意义。
2 结果
2.1 两组小鼠胚胎表型及 HE 染色结果
对照组小鼠胚胎发育第11.5天胚胎外观较为饱满圆滑,头部前、中及后脑泡均已经发育形成,脊柱表面完整无裂口(见图1A);可见明显的耳泡、眼泡,尾部弯曲细长。HE染色显示神经管发育良好,细胞排列密集整齐(见图1B、C)。
NTD组小鼠胚胎的后脑部位神经管未闭合,神经组织裸露在胚胎外(见图1D)。HE染色显示后脑顶板未闭合,神经腔面未完整,且细胞排列疏松(见图1E、F)。
2.2 两组小鼠胚胎一般资料比较
两组胚胎的顶臀长度比较,采用t检验,差异有统计学意义(t=24.480,P=0.002),NTD组短于对照组。两组胚胎的体重比较,采用t检验,差异有统计学意义(t=41.860,P=0.001),NTD组低于对照组。两组胚胎畸形率比较,采用Fisher确切概率法,差异有统计学意义(P=0.000)。见表1。
2.3 两组INPP5E基因启动子区甲基化水平比较
在引物扩增的328 bp区域中,包含8个CpG二核苷酸。对照组与NTD组CpG甲基化水平分别为40.6%和9.4%,NTD组与对照组甲基化程度的比率为23.2%。两组INPP5E基因启动子区的甲基化水平比较,采用χ2检验,差异有统计学意义(χ2=33.330,P=0.000),NTD组低于对照组。见图2。

图1 两组小鼠胚胎发育第11.5天的胚胎表型及HE染色结果
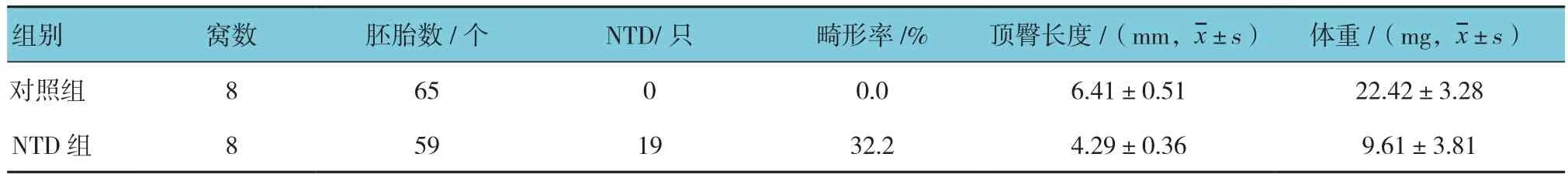
表1 两组胚胎一般资料比较
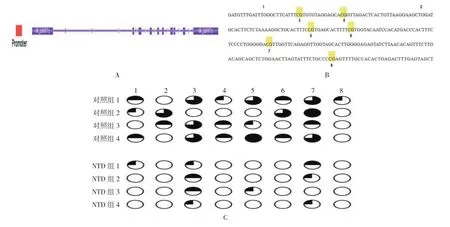
图2 INPP5E 基因 CpG 岛的甲基化
2.4 两组小鼠胚胎组织中INPP5E相对表达量比较
对照组和NTD组小鼠胚胎发育第11.5天时INPP5E蛋白相对表达量分别为(1.000±0.230)和(0.482±0.080),经t检验,差异有统计学意义(t=5.981,P=0.027),NTD组低于对照组。对照组和NTD组INPP5E mRNA相对表达量分别为(1.000±0.117)和(0.623±0.034),经t检验,差异有统计学意义(t=7.867,P=0.016),NTD组低于对照组。NTD组小鼠胚胎神经组织中,INPP5E基因启动子区的低甲基化可能是引起INPP5E mRNA表达降低的原因,从而影响胚胎神经管的发育。见图3、4。
2.5 两组小鼠胚胎神经组织中FA及其相关代谢产物水平比较
在最佳的超高效液相色谱条件下分离检测FA及其相关代谢产物5-甲基四氢FA(5-methyltetrahydrofolate,5-MeTHF)、5- 甲酰四氢 FA(5-formyltetrahydrofolate,5-FoTHF)和同型半胱氨酸(Homocysteine,Hcy)水平,分别在2.82、2.53、2.88和0.71 min被洗脱,进一步通过串联质谱进行定量分析。见图4。

图3 两组小鼠胚胎组织中 INPP5E 蛋白和mRNA 表达水平比较(n =8,±s)
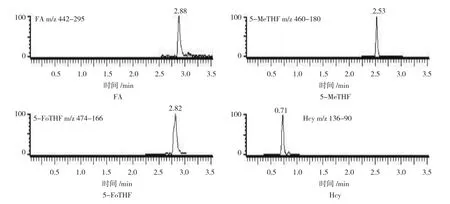
图4 FA及其相关代谢产物质谱多反应监测色谱图
FA在对照组及NTD组中的含量分别为(2.76±0.98)和(4.39±1.41)mg/g,5-MeTHF的含量分别为(9.55±2.38)和(4.62±1.98)mg/g,5-FoTHF的含量分别为(4.86±1.02)和(3.19±0.75)mg/g,Hcy的含量分别为(4.33±1.42)和(7.06±1.38)mg/g。两组小鼠胚胎神经组织中FA、5-MeTHF、5-FoTHF及Hcy水平比较,经t检验,差异有统计学意义(t=6.566、21.350、10.710 和118.200,P=0.022、0.002、0.009和0.000),NTD组小鼠胚胎神经组织中FA和Hcy水平高于对照组,NTD组小鼠胚胎神经组织中5-MeTHF和5-FoTHF水平低于对照组。见图5。
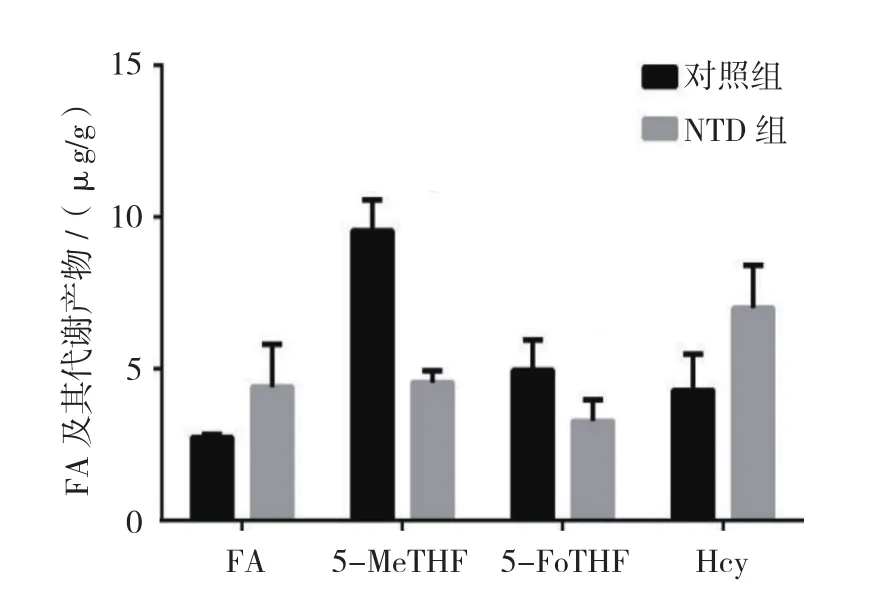
图5 两组小鼠胚胎神经组织中FA及其相关代谢产物水平比较(n =8,±s)
3 讨论
INPP5E基因调节肌醇信号通路活性,参与胚胎神经发育,其编码的INPP5E蛋白存在于神经细胞的细胞质、细胞骨架、纤毛轴丝、高尔基体、高尔基体膜、外周膜蛋白及质膜内侧[11-14]。有研究发现,INPP5E基因突变导致茹贝尔综合征等疾病[15-17]。有研究显示,INPP5E基因表达影响初级纤毛稳定性,进而调控神经管的闭合[18-20]。有学者报道INPP5E基因突变与神经管畸形相关,神经管畸形是纤毛相关疾病的主要表型之一,是一种严重的出生缺陷,是由多种因素导致的神经管部分或完全闭合不全,进而影响神经系统发育[21]。INPP5E基因突变会引起PI(3,4,5)P3和PI(4,5)P2的生成产物减少,并通过改变PI3K信号通路影响细胞增殖、存活、凋亡及初级纤毛稳定性,从而导致后脑畸形及纤毛相关疾病[22-25]。
DNA甲基化是一种重要的表观遗传修饰,参与基因表达调控,影响胚胎生长发育[26]。低甲基化的DNA在有丝分裂中易发生重组,导致基因发生缺失或易位,最终影响基因表达[27]。有研究报道NTD小鼠胚胎中,与细胞增殖相关的Ptch1、Pla2g4a、Foxgl基因的甲基化水平与对照组相比显著降低,且qRT-PCR结果与甲基化水平改变相关,推测甲基化的改变在NTD发生过程中发挥重要作用[28]。本研究采用腹腔注射4.5 mg/kg甲氨蝶呤,干扰FA代谢从而复制NTD小鼠模型,通过检测对照组及NTD组小鼠胚胎中的FA及其代谢产物,发现NTD组小鼠胚胎神经组织的FA含量明显升高,且代谢产物5-MeTHF和5-FoTHF的含量明显降低,说明此时FA代谢呈现紊乱趋势;INPP5E基因启动子区甲基化水平的显著降低,推测FA代谢失衡引起甲基化供体不足,导致INPP5E基因启动子区甲基化水平显著降低,从而影响该基因的正常表达,干扰神经管的闭合。以上结果表明在神经发育早期,需要INPP5E基因在一定水平上表达调控神经管正常闭合。
本研究发现NTD小鼠胚胎INPP5E蛋白和mRNA相对表达量低于正常小鼠。说明INPP5E基因在发育早期影响神经系统发育,本研究结果表明NTD小鼠胚胎神经组织的INPP5E蛋白和mRNA相对表达量下降,参与FA代谢紊乱神经管闭合失败,造成小鼠露脑畸形。INPP5E基因表达水平的改变是否通过PI3K信号通路活性影响初级纤毛形成,其具体机制尚需在细胞内干扰INPP5E基因的表达,如用CRISPR-Cas9基因编辑技术在小鼠胚胎神经干细胞上进行INPP5E基因敲除实验。同时,检测PI(3,4,5)P3和PI(4,5)P2及初级纤毛的形成可进一步研究该基因对胚胎神经发育的影响。
因此,INPP5E基因在调节胚胎神经发育中起重要作用,NTD组小鼠胚胎神经组织中该基因的表达降低,推测由于FA代谢失衡引起甲基化供体不足,使该基因启动子区发生低甲基化,进而引起基因异常表达导致NTD的发生。对INPP5E基因调控胚胎神经发育分子机制的进一步阐明,对神经系统发育缺陷相关疾病的早期诊断及防控具有重要的意义。
