不同年龄腰突出椎间盘组织CCNDBP1表达及意义
2019-09-10慈延东邱晨生吴晓淋方晓露相宏飞陈伯华
慈延东 邱晨生 吴晓淋 方晓露 相宏飞 陈伯华
[摘要]目的比较细胞周期D1型结合蛋白1(CCNDBP1)在不同年龄阶段腰椎间盘突出症病人椎间盘中的表达,探讨其与腰椎间盘退变的关系。方法收集不同年龄阶段腰椎间盘突出症病人共42例,其中30~岁、40~岁、50~岁年龄段病人各14例。取病人突出的椎间盘组织,苏木精`-伊红染色观察其退变情况,采用免疫组化法、PCR法、Western印迹法从细胞、基因、蛋白水平檢测CCNDBP1的表达量。结果苏木精`-伊红染色结果显示,各个年龄阶段腰椎间盘突出症病人腰椎间盘组织均有不同程度退变,退变程度与年龄呈正相关。免疫组化法、PCR法和Western印迹法检测结果显示,腰椎间盘组织CCNDBP1的表达随病人年龄的增加而减少,其中30~岁年龄段病人CCNDBP1的表达显著高于40~岁年龄段病人,差异有统计学意义(F=15.42~168.30,t=3.06~6.78,P<0.05),40~岁年龄段病人CCNDBP1的表达显著高于50~岁年龄段病人,差异有统计学意义(t=2.58~9.91,P<0.05)。结论人腰椎间盘髓核组织退变过程中有CCNDBP1的参与,随着年龄的增长,CCNDBP1在退变腰椎间盘组织中的表达量逐渐减少。
[关键词]椎间盘移位;腰椎;细胞周期蛋白质类;年龄因素
[中图分类号]R681.53[文献标志码]A[文章编号] 2096`-5532(2019)02`-0128`-05
doi:10.11712/jms201902002[开放科学(资源服务)标识码(OSID)]
EXPRESSION AND SIGNIFICANCE OF CYCLIN`-D1`-BINDING PROTEIN 1 IN THE INTERVERTEBRAL DISC OF PATIENTS WITH LUMBAR DISC HERNIATION AT DIFFERENT AGES CI Yandong, QIU Chensheng, WU Xiaolin, FANG Xiaolu, XIANG Hongfei, CHEN Bohua(Department of Spinal Surgery, The Affiliated Hospital of Qingdao University, Qingdao 266100, China)
[ABSTRACT]ObjectiveTo investigate the expression of cyclin`-D1`-binding protein 1 (CCNDBP1) in the intervertebral disc of patients with lumbar disc herniation at different ages and its association with lumbar disc degeneration. MethodsA total of 42 patients with lumbar disc herniation at different ages were enrolled, with 14 patients each in the 30-, 40-, and 50- year groups. The prominent intervertebral disc tissue was collected, and HE staining was used to observe disc degeneration. Immunohistoche`-mistry was used to measure the expression of CCNDBP1 in cells, and PCR and Western blot were used to measure the mRNA and protein expression of CCNDBP1. ResultsHE staining showed varying degrees of lumbar disc degeneration in the patients with lumbar disc herniation at different ages, and the degree of degeneration was positively correlated with age. Immunohistochemistry, PCR, and Western blot showed that the expression of CCNDBP1 in the intervertebral disc decreased with age; the 30- year group had significantly higher expression of CCNDBP1 than the 40- year group (F=15.42-168.30,t=3.06-6.78,P<0.05), and the 40- year group had significantly higher expression of CCNDBP1 than the 50- year group (t=2.58-9.91,P<0.05). ConclusionCCNDBP1 is involved in the degeneration of human lumbar disc nucleus pulposus, and the expression of CCNDBP1 in the degenerated intervertebral disc gradually decreases with age.
[KEY WORDS]intervertebral disc displacement; lumbar vertebrae; cell cycle proteins; age factors
腰椎间盘位于椎体之间,由髓核、纤维环及软骨终板组成。腰椎间盘突出症已成为中老年人群较为常见的脊柱退行性疾病之一[1],也是导致腰腿痛的主要原因[2]。近年来,我国腰椎间盘突出症的发病率较以往有所提高,发病年龄有提前的趋势[3`-4]。随着对腰椎间盘突出症病理研究的逐渐深入,细胞因子、炎性递质等在椎间盘突出症中作用的研究日益增多,目前腰椎间盘突出症发病机制的研究主要集中于损伤、退变、炎症、免疫、遗传等方面,但相应的病因机制还不完全清楚[4]。近期有研究表明,细胞周期D1型结合蛋白1(CCNDBP1)可能参与了腰椎间盘退行性改变[5],但其在椎间盘内表达情况及在椎间盘退行性变中的作用尚不明确。本研究拟检测CCNDBP1在不同年龄段腰椎间盘突出症病人腰椎间盘组织中的表达,分析CCNDBP1与腰椎间盘退变的关系,为延缓椎间盘退变提供新的方向。
1资料与方法
1.1一般资料
选取2017年11月—2018年3月在我院行手术治疗的腰椎间盘突出症病人42 例。病人年龄为30~60岁,平均43岁,其中30~岁、40~岁、50~岁年龄段病人各14例。术前行腰椎 X 线片及腰椎CT检查诊断为单纯腰椎间盘突出,并经腰椎MRI检查证实,腰椎间盘退变临床分级均为Pfirrmann Ⅲ~Ⅴ级。病人查体符合腰椎间盘突出症临床诊疗共識标准,且均有1~2项肯定的对应神经症状及体征,均无腰椎侧弯、腰椎滑脱和腰椎骨折等其他腰椎病变。本研究经青岛大学附属医院伦理委员会批准,病人及家属均签署知情同意书。
1.2标本处理
术中取出的椎间盘组织用灭菌生理盐水清洗后,取部分送病理检查,其余组织置液氮中保存待用。选择术中收集的部分腰椎间盘组织(选取L3/4、L4/5、L5/S1节段),用40 g/L的甲醛溶液固定,脱水,石蜡包埋,5 μm厚切片。所有标本均选取5张有髓核细胞切片,其中2张切片行苏木精`-伊红染色,另3张切片进行免疫组化染色。
1.3主要材料
TRIzol Reagent 购自TaKaRa公司;CCNDBP1一抗购自美国Abcam公司;CCNDBP1二抗和内参一抗购自武汉博士德生物技术有限公司;DAB显色试剂盒购自美国 Millipore公司。
1.4检测方法
1.4.1苏木精`-伊红染色椎间盘组织石蜡切片行苏木精`-伊红染色后,在高倍(200倍)光学显微镜下观察椎间盘细胞及细胞外胶原纤维形态学变化。
1.4.2免疫组化染色检测CCNDBP1的表达将椎间盘组织切片置于荷电切片上,在60 ℃下烘烤40 min,行去亲合、再水化,放入沸水中2 min,用柠檬酸钠缓冲液(pH值6.0)进行热抗原修复40 min,冷却至室温后,用蒸馏水和PBS洗涤15 min,将体积分数为0.03的过氧化氢滴加至切片上,37 ℃孵育10 min,加入CCNDBP1一抗(1∶500)孵育1 h,用蒸馏水和PBS洗涤后,加入CCNDBP1二抗孵育40 min,组织电位暴露1 min,苏木精作用50 s,分化液作用1 s,滴加中性树胶,封片后显微镜下观察。
1.4.3PCR方法检测CCNDBP1 mRNA的表达从液氮中取出椎间盘组织制作匀浆,取100 mg加1 mL TRIzol提取总RNA。震荡混匀后室温静置5 min,移入EP管中,4 ℃下以12 000 r/min离心5 min,吸取上清液,移入新的EP管中,加入200 μL的氯仿,上下混匀后室温静置5 min,在4 ℃下以12 000 r/min离心15 min,吸取上层水相移入新的EP管中,加入0.5 mL的异丙醇混匀,室温放置10 min,4 ℃下以12 000 r/min离心10 min,去除上清液体,用1 mL体积分数0.75的乙醇洗涤沉淀,悬浮后4 ℃下以7 500 r/min离心5 min,弃上清,将沉淀物室温干燥 10 min,溶于DEPC 水中。按照反转录试剂盒说明书将mRNA反转录得到cDNA。取5 μL PCR产物进行琼脂糖凝胶电泳,用凝胶成像系统记录电泳结果,计算机进行图像处理。另取2 μL PCR产物进行qRT`-PCR。CCNDBP1上游引物为5′`-ACAAGTCCATGCTGCCATCAAGG`-3′,下游引物为 5′`-GAGCTGAGCCATGCCATCCAC`-3′。按照转录试剂盒说明书上样,在 PCR 仪上进行基因扩增。采用2-△△Ct法计算各组样本CCNDBP1的表达量。实验重复3次,取均值。
1.4.4Western法检测CCNDBP1蛋白的表达将椎间盘组织从液氮中取出,用剪刀剪碎,加入蛋白提取裂解液RIPA和蛋白酶抑制剂,用组织匀浆器充分研磨组织,转入EP管中,4 ℃下以12 000 r/min离心20 min,吸取上清液移入新的EP管中,提取出组织中总蛋白。蛋白用紫外线分光光度计进行定量,加入蛋白上样缓冲液后保存于-80 ℃冰箱中。取等量蛋白质用40~120 g/L的SDS`-PAGE凝胶电泳,将蛋白印迹转至PVDF膜上(恒压110 V,电流300 mA,1.5 h),加入50 g/L脱脂奶粉`-TBST封闭液常温封闭1.5 h。加GAPDH单克隆一抗(1∶3 000)、CCNDBP1一抗(1∶1 000),置4 ℃冰箱中过夜。加入二抗室温下摇1 h,用化学发光试剂显影成像。计算机图像处理后测定各个条带的灰度值,目的蛋白表达量以目的条带与内参照条带灰度值的比值表示。实验重复3次,取均值。
1.5统计学分析
应用SPSS 22.0软件进行统计学分析,实验数据均以±s表示,多组均数比较采用方差分析。
2结果
2.1苏木精`-伊红染色
各个年龄段腰椎间盘突出症病人腰椎间盘组织结构均有不同程度的破坏,细胞核和细胞质模糊程
130青岛大学学报(医学版)55卷
度以及细胞稀疏程度随年龄的增大逐渐加重,髓核细胞数量逐渐减少,髓核细胞形态不规则程度加重。
2.2免疫组化染色
所有病人椎间盘组织中均有CCNDBP1的表达,30~岁年龄段病人椎间盘组织中CCNDBP1阳性细胞数量最多,颜色最深;40~岁年龄段病人阳性细胞数量次之,颜色较浅;50~岁年龄段病人阳性细胞数量最少,颜色最浅。
2.3不同年龄段病人腰椎间盘组织中CCNDBP1 mRNA表达比较
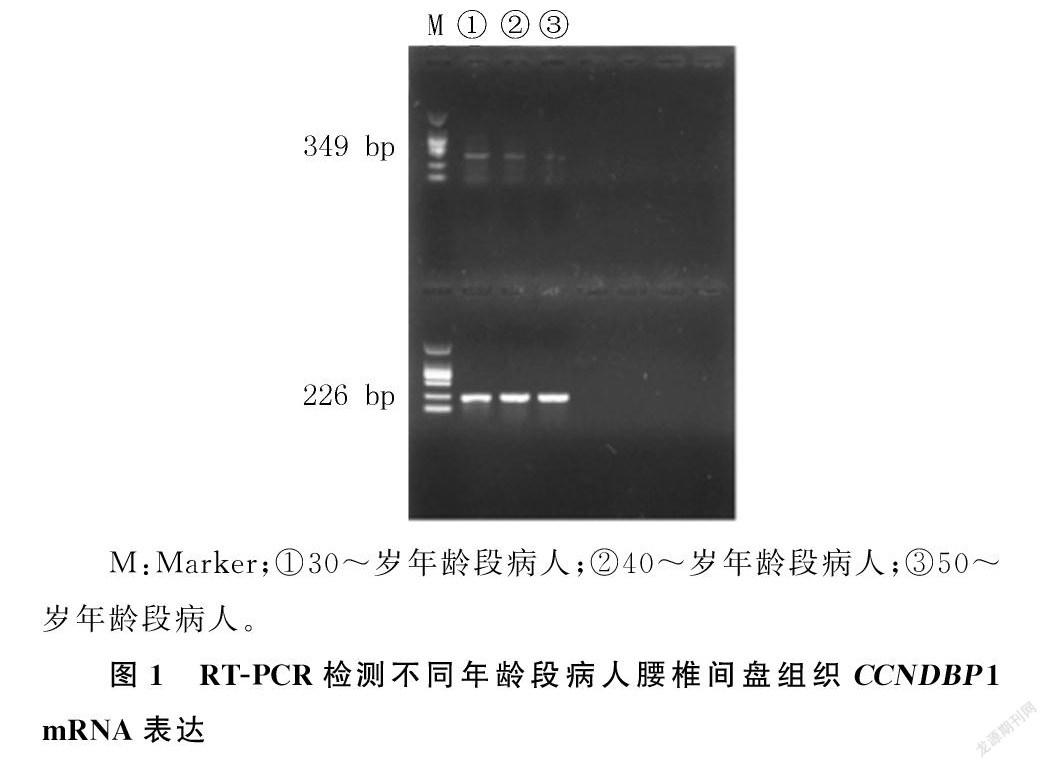
2.3.1RT`-PCR检测30~岁、40~岁、50~岁年龄段病人椎间盘组织均有一定程度的CCNDBP1 mRNA表达,且其表达随腰椎间盘突出症病人年龄的增大而下降。见图1。
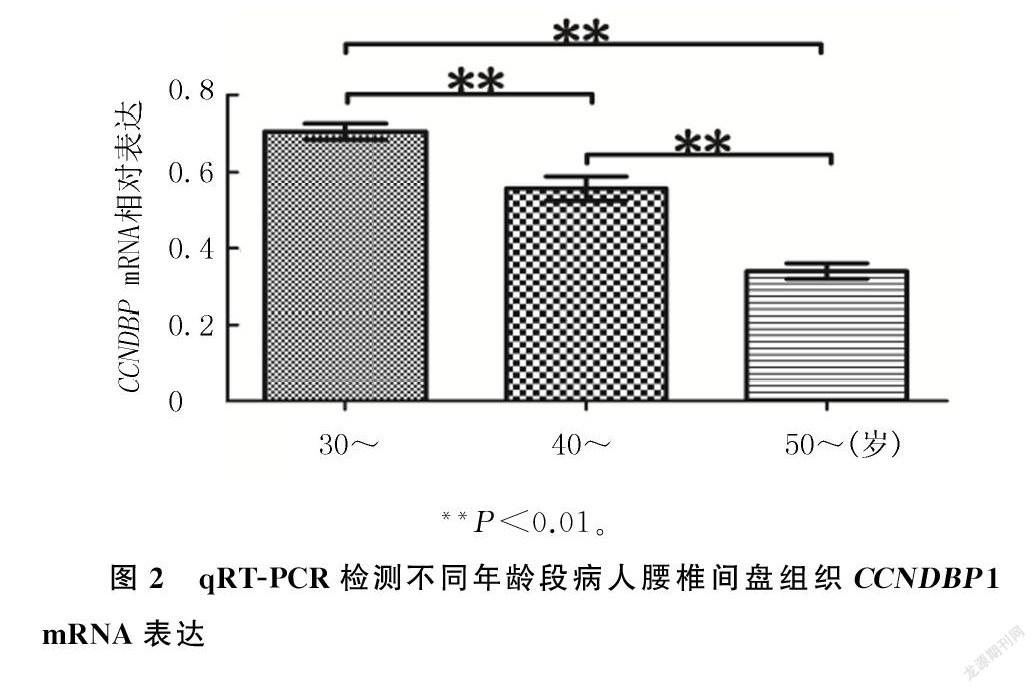
2.3.2qRT`-PCR检测腰椎间盘突出症病人年龄越大,其腰椎间盘组织CCNDBP1 mRNA的表达越低。30~岁年龄段病人CCNDBP1 mRNA表达水平显著高于40~岁年龄段病人(F=168.30,t=6.78,P<0.01),而40~岁年龄段病人CCNDBP1 mRNA的表达又显著高于50~岁年龄段病人(t=9.91,P<0.01)。见图2。
2.4不同年龄段病人腰椎间盘组织CCNDBP1蛋白表达比较
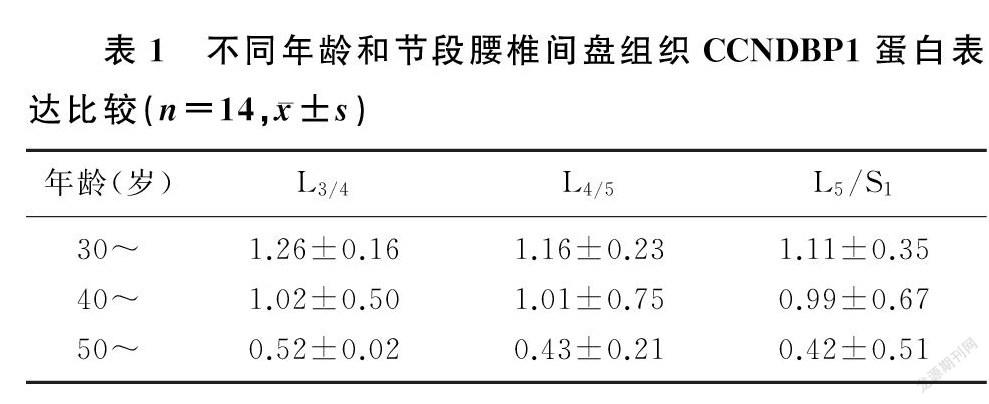
腰椎间盘突出症病人年龄越大,其腰椎间盘组织CCNDBP1蛋白的表达越低。30~岁年龄段病人不同节段腰椎间盘组织CCNDBP1的表达显著高于40~岁年龄阶段病人(F=15.42~23.38,t=3.06~3.83,P<0.05),40~岁年龄段病人CCNDBP1的表达显著高于50~岁年龄阶段病人(t=2.58~4.50,P<0.05)。同一年龄段病人不同节段腰椎间盘组织CCNDBP1蛋白的表达比较,差异均无统计学意义(P>0.05)。见表1。
3讨论
椎间盘退变被认为是椎间盘突出等疾病的病理基础,人们已经从分子生物学、生物化学和免疫学等方面对退变椎间盘进行了相关的研究,但椎间盘退变的确切机制仍存在争议,需要进一步的研究[6]。近年研究表明,细胞因子在腰椎间盘退变、突出过程中起着极为重要的作用[7`-8]。有研究发现,腰椎间盘突出症病人的椎间盘标本中存在免疫相关细胞因子的表达[9]。NAYLOR等[10]首先提出了椎间盘突出的自身免疫学说,认为椎间盘组织是一种潜在的抗原,当纤维环破裂后会产生自身免疫反应。研究证明,自身抗体通过巨噬细胞系统,并与细胞免疫共同作用于椎间盘,引起椎间盘组织的损伤,造成椎间盘生物力学改变,使椎间盘基质降解加速,促进椎间盘突出[11`-13]。KAWAGUCHI等[14]研究结果表明,椎间盘突出与炎症反应致组织损伤或化学刺激有关。TAPIA等[15]研究表明,椎间盘损伤修复过程中有巨噬细胞和肥大细胞浸润,释放大量的细胞因子,导致免疫反应引起组织损伤。
CCNDBP1最初由TERAI等[16]和YAO等[17]确定是细胞周期蛋白D(Cyclin D)与白细胞特异性适配蛋白(Grap2)相互作用结合的蛋白。XIA等[18]研究表明,CCNDBP1在肌肉、心脏、白细胞和大脑中的表达是最高的。CCNDBP1通过Grap2和Cyclin D介导的信号通路参与调节细胞的分化和增殖,
2期慈延东,等. 不同年龄腰突出椎间盘组织CCNDBP1表达及意义131
从而启动DNA的合成,促进细胞增殖,抑制细胞凋亡。CCNDBP1含有一个假定的HLH结构,没有基本的DNA结合结构域,可能作为转录因子(如ID蛋白家族的主要负调节因子)发挥作用[19`-22]。椎间盘髓核细胞衰老是已经确认的引起椎间盘退变的重要原因[23]。有研究表明,CCNDBP1在免疫系统细胞信号机制和转录调控肝脏特异性基因的突变机制中起作用[24]。BABU等[5]研究结果显示,正常椎间盘组织CCNDBP1的表达明显高于退变椎间盘组织。CCNDBP1表达可能与抑制细胞凋亡、促进细胞的分化和增殖有关[12]。本实验从细胞、基因、蛋白水平检测CCNDBP1的表达,结果显示,腰椎间盘突出症病人椎间盘组织中CCNDBP1的表达随年龄的增长而降低,提示CCNDBP1可能通过参与椎间盘退变过程中免疫反应并抑制椎间盘细胞的凋亡来延缓椎间盘的退变。
有研究表明,CCNDBP1在终末分化组织中高度表达,在增殖细胞中的表达减少[25]。目前,关于CCNDBP1的研究多集中在肿瘤领域,而在椎间盘方面的研究尚未见报道。MA等[26]研究显示,在结肠肿瘤中CCNDBP1通过特异性小干扰RNA表达促进了集落形成而表达减少,表明CCNDBP1可以作为潜在的肿瘤因子。还有研究表明,CCNDBP1可以在细胞分化和增殖控制方面发挥重要作用,已被确定为潜在的原癌基因[27`-29]。本研究通过苏木精`-伊红染色和免疫组化染色观察到,退变椎间盘组织细胞数量明显减少,细胞形态不规则,随年龄的增大细胞数量更少,表明CCNDBP1可能参与细胞的增殖过程。本文研究结果还显示,CCNDBP1在不同年龄阶段腰椎间盘突出症病人椎间盘组织中均有表达,随年龄增加其表达量逐渐减少,初步揭示了CCNDBP1在椎间盘退变中的作用。当退变或外伤导致椎间盘自身抗原成分与血液循环接触后,机体会产生自身免疫、炎症反应,腰椎间盘突出发生过程可能与免疫及炎症反应有关。NERLICH 等[30]研究结果表明,人类椎间盘髓核组织中存在CD68 阳性的炎性细胞,它们参与了椎间盘细胞外基质的吞噬降解过程。CCNDBP1可能通过参与免疫反应,如通过影响CD68细胞抑制椎间盘细胞凋亡,促进椎间盘细胞的分化与增殖,缓解椎间盘退变,今后可研究椎间盘组织中的炎性细胞因子以进一步揭示CCNDBP1的作用机制。
综上所述,CCNDBP1在腰椎间盘组织中的表达随年龄的增长而减少。本实验结果初步揭示了CCNDBP1与椎间盘退变的关系,为延缓椎间盘退变提供了新的方向与线索。但椎间盘退变的机制较为复杂,CCNDBP1在椎间盘退變进程中的具体作用机制及其与其他因素的关联及相互影响仍需进一步探讨。
[参考文献]
[1]姜棚菲,马张稳,张民泽,等. 腰椎间盘退行性改变患者ADAMTS`-7表达及其机制研究[J]. 实用医院临床杂志, 2017,14(6):101`-104.
[2]殷相姣,宫赫,王丽珍,等. 椎间盘退行性变的影响因素及其相关形态结构变化的研究进展[J]. 生物医学工程与临床, 2016,33(1):112`-117.
[3]曾佳兴,梁斌,尹东,等. 青少年与中老年腰椎间盘突出相关因素分析[J]. 中国矫形外科杂志, 2013,21(11):1121`-1126.
[4]曾佳兴,梁斌,尹东,等. MMP`-3、IgG和CD68在青少年与中老年突出腰椎间盘组织中的表达分析[J]. 中国脊柱脊髓杂志, 2013,23(12):1109`-1115.
[5]BABU N S, KRISHNAN S S, CHERUKUVADA V B, et al. Quantitative proteomic analysis of normal and degenerated human intervertebral disc[J]. Spine Journal, 2016,16(8):989`-1000.
[6]王宗亮,劉雅,蔡明,等. 组织蛋白酶K与人椎间盘退变相关性研究[J]. 创伤外科杂志, 2009,11(3):245`-247.
[7]MAITRE C L, FREEMONT A J, HOYLAND A A. The role of interleukin`-1 in the pathogenesis of human intervertebral disc degeneration[J]. Arthritis Research & Therapy, 2005,7(4):R732`-R745.
[8]HARO H, KOMORI H, KATO T, et al. Experimental stu`-dies on the effects of recombinant human matrix metalloprotei`-nases on herniated disc tissues-how to facilitate the natural resorption process of herniated discs[J]. Journal of Orthopaedic Research, 2010,23(2):412`-419.
[9]ROBERTS S, EVANS H, MENAGE J, et al. TNFα`-stimulated gene product (TSG`-6) and its binding protein, IαI, in the human intervertebral disc:new molecules for the disc[J]. European Spine Journal, 2005,14(1):36`-42.
[10]NAYLOR A, HAPPEY F, TURNER R L, et al. Enzymic and immunological activity in the intervertebral disk[J]. The Orthopedic Clinics of North America, 1975,6(1):51`-58.
[11]HABTEMARIAM A, VIRRI J, GRONBLAD M, et al. The role of mast cells in disc herniation inflammation[J]. Spine, 1999,24(15):1516`-1520.
[12]王沛,董强,雪原. 腰椎间盘突出病变部的压力测定及其病理学意义[J]. 中华骨科杂志, 2002,22(3):129`-133.
[13]GEISS A, LARSSON K, RYDEVIK B, et al. Autoimmune properties of nucleus pulposus:an experimental study in pigs[J]. Spine, 2007,32(2):168`-173.
[14]KAWAGUCHI S, YAMASHITA T, YOKOGUSHI K, et al.
132青岛大学学报(医学版)55卷
Immunophenotypic analysis of the inflammatory infiltrates in herniated intervertebral discs[J]. Spine, 2001,26(11):1209`-1214.
[15]TAPIAPEREZ H. Intervertebral disc pathologies from an immunological perspective[J]. Revista de Neurologia, 2008,46(12):751`-757.
[16]TERAI S, AOKI H, ASHIDA K, et al. Human homologue of maid:a dominant inhibitory helix`-loop`-helix protein associated with liver`-specific gene expression[J]. Hepatology, 2000,32(2):357`-366.
[17]YAO Y, DOKI Y W, IMOTO M, et al. Cloning and characterization of Dip1, a novel protein that is related to the Id fa`-mily of proteins[J]. Experimental Cell Research, 2000,257(1):22`-32.
[18]XIA C, BAO Z, TABASSAM F, et al. GCIP, a novel human grap2 and cyclin D interacting protein, regulates E2F`-mediated transcriptional activity[J]. Journal of Biological Chemistry, 2000,275(27):20942`-20948.
[19]BENEZRA R, DAVIS R L, LASSAR A, et al. The protein Id:a negative regulator of helix`-loop`-helix DNA binding proteins[J]. Cell, 1990,61(1):49`-59.
[20]CHRISTY B A, SANDERS L K, LAU L F, et al. An Id`-related helix`-loop`-helix protein encoded by a growth factor`-inducible gene[J]. Proceedings of the National Academy of Sciences of the United States of America, 1991,88(5):1815`-1819.
[21]DEED R W, BIANCHI S M, ATHERTON G T, et al. An immediate early human gene encodes an Id`-like helix`-loop`-helix protein and is regulated by protein kinase C activation in diverse cell types[J]. Oncogene, 1993,8(3):599`-607.
[22]RIECHMANN V, VAN C I, SABLITZKY F. The expression pattern of Id4, a novel dominant negative helix`-loop`-helix protein, is distinct from Id1, Id2 and Id3[J]. Nucleic Acids Research, 1994,22(5):749`-755.
[23]常獻,陈斌,李长青. 髓核细胞老化机制研究进展[J]. 中国矫形外科杂志, 2014,22(1):40`-42.
[24]MEZIANI R, YAMADA R, TAKAHASHI M, et al. A tran`-sethnic genetic study of rheumatoid arthritis identified FCGR2A as a candidate common risk factor in Japanese and European populations[J]. Japanese Journal of Rheumatology, 2012,22(1):52`-58.
[25]XIANG Hua, WANG Juan, MAO Yingwei, et al. Human telomerase accelerates growth of lens epithelial cells through regulation of the genes mediating RB/E2F pathway[J]. Oncogene, 2002,21(23):3784`-3791.
[26]MA W, STAFFORD L J, LI D, et al. GCIP/CCNDBP1, a helix`-loop`-helix protein, suppresses tumorigenesis[J]. Journal of Cellular Biochemistry, 2010,100(6):1376`-1386.
[27]RUZINOVA, MARIANNA B, BENEZRA, et al. Id proteins in development, cell cycle and cancer[J]. Trends in Cell Biology, 2003,13(8):410`-418.
[28]SIKDER H A, DEVLIN M K, DUNLAP S, et al. Id proteins in cell growth and tumorigenesis[J]. Cancer Cell, 2003,3(6):525`-530.
[29]WANG Q, HII G, SHUSTERMAN S, et al. ID2 expression is not associated with MYCN amplification or expression in human neuroblastomas[J]. Cancer Research, 2003,63(7):1631`-1635.
[30]NERLICH A G, WEILER C, ZIPPERER J, et al. Immunolocalization of phagocytic cells in normal and degenerated intervertebral discs[J]. Spine, 2003,28(15):2484`-2490.
.
