乳酸菌代谢γ-谷氨酰二肽及其对馒头滋味特性的影响
2019-09-06闫博文杨化宇蔡一芥范大明连惠章赵建新
闫博文,杨化宇,蔡一芥,范大明,连惠章,陈 卫,张 灏,赵建新,*
(1.江南大学 食品科学与技术国家重点实验室,江苏 无锡 214122;2.江南大学食品学院,江苏 无锡 214122;3.苏州旅游与财经高等职业技术学校,江苏 苏州 215104;4.无锡华顺民生食品有限公司,江苏 无锡 214218)
馒头作为我国传统发酵主食拥有悠久历史,且深受广大消费者的喜爱[1]。随着工业水平的不断发展与进步,馒头的发酵主要采用即发型活性酵母粉,其具有发酵速率快且产品批次稳定性高等优点。然而,我国山东和河南等北方地区,仍传承着传统酸面团发酵工艺,制得馒头产品风味浓郁,质地更佳,始终是消费者的第一选择[2-3]。这主要是由于酸面团发酵过程中伴随着内源性谷物蛋白酶对大分子蛋白质聚合物的降解作用,产生大量的游离氨基酸,而酸面团中微生物可通过自身代谢将其生成具有改善产品风味的活性物质[4]。研究表明乳酸菌可代谢合成具有浓厚味(Kokumi)的滋味活性肽,此类物质可有效提高产品的滋味特性,如甜味、咸味以及酸味等,对产品感官品质有积极的作用[5-6]。
谷胱甘肽作为一种γ-谷氨酰三肽,是最早被发现具有浓厚味的活性滋味肽,其主要来源于酵母提取物和部分谷物[7]。谷胱甘肽的合成主要与酵母菌的自溶以及谷胱甘肽合成酶有关,且研究发现其对帕尔马干酪的滋味提升有重要作用[8-9]。然而,谷胱甘肽在γ-谷氨酰转移酶的作用下,可生成多种γ-谷氨酰二肽,其是一类具有浓厚味的滋味活性二肽,其滋味阈值在0.005~0.01 μmol/kg[5],广泛存在于奶酪、酱油和酸面团等发酵制品中[8,10-11]。由于其滋味阈值较低,且具有改善发酵产品滋味特性的潜在能力,因此近年来受到学者们的广泛关注。Zhao等[10]采用罗伊氏乳杆菌发酵酸面团观察其对γ-谷氨酰二肽合成的影响,发现乳酸菌代谢合成γ-谷氨酰二肽存在株间水平差异。然而,针对酸面团中常见的不同乳酸菌菌种对γ-谷氨酰二肽代谢合成能力的相关研究尚鲜见报道,且含有不同生成量γ-谷氨酰二肽的馒头制品在滋味上的差异性尚不明确。因此,本研究采用分离自酸面团中的常见乳酸菌发酵,观察比较不同乳酸菌在酸面团发酵过程中对γ-谷氨酰二肽合成量的种间水平差异,及其对产品滋味特性的影响。
1 材料与方法
1.1 材料与试剂
罗伊氏乳杆菌分离自酸面团,受赠于加拿大阿尔伯塔大学;发酵乳杆菌、植物乳杆菌、食窦魏斯氏乳杆菌、旧金山乳杆菌分离自酸面团,江南大学食品学院生物技术中心保藏;即发型活性干酵母 安琪酵母股份有限公司;小麦粉 加拿大Robin Hood公司;γ-谷氨酰二肽分析纯标准品(γ-谷氨酰谷氨酸、γ-谷氨酰缬氨酸、γ-谷氨酰异亮氨酸和γ-谷氨酰亮氨酸)由美国生物公司合成制备。
1.2 仪器与设备
SM-25搅面机、SK-12P醒发箱 新麦机械(无锡)有限公司;HB24D552W电蒸箱 博西家用电器(中国)有限公司;高效液相色谱、液相色谱-二级质谱联用仪 美国赛默飞世尔科技公司;Smart Tongue电子舌系统 上海瑞玢智能科技有限公司。
1.3 方法
1.3.1 菌种活化与酸面团的制备
将-80 ℃保菌管内菌液进行菌落划线,置于厌氧培养箱内37 ℃培养18 h,挑取单菌落接种至1 mL mMRS液体培养基中,37 ℃培养16 h,活化2 代,备用。取500 μL乳酸菌发酵液接种至50 mL mMRS液体培养基中,37 ℃培养16 h,得到菌液6 000×g离心10 min,去上清液,加入等体积生理盐水洗涤菌泥2 次,而后加入无菌水50 mL和小麦粉50 g,混匀,37 ℃培养48 h,制得酸面团备用[12]。化学酸化面团,即在面团制备过程中添加乳酸和乙酸混合溶液(体积比4∶1),调节初始面团pH 3.5。
1.3.2 酸面团pH值和微生物菌落总数的测定
采用酸面团中分离筛选得到的5 株常见乳酸菌发酵酸面团,并对其pH值和菌落总数进行比较分析。称取10 g酸面团样品,加入90 mL蒸馏水,搅拌20 min,静置5 min,测定酸面团pH值;称取10 g酸面团样品,加入90 mL无菌生理盐水,均质机拍打10 min,通过梯度稀释,涂布,厌氧培养,测定酸面团中微生物菌落总数[13]。
1.3.3 酸面团中游离氨基酸总量的测定
游离氨基酸是γ-谷氨酰二肽的重要合成底物,采用不同乳酸菌发酵酸面团考察其对游离氨基酸总量的影响。称取酸面团样品10 g,加入5%三氯乙酸溶液50 mL,高速匀浆1 min,再以10 000 r/min离心10 min,将上清液用两层滤纸过滤,滤液以10 000 r/min离心10 min后用邻苯二醛进行柱前衍生,然后高效液相色谱进样进行游离氨基酸分析,每个样品至少重复2 次操作[14]。
色谱条件:采用ODS Hypersil毛细管色谱柱(250 mm×4.6 mm,5 μm),流动相为20 mmol/L乙酸钠的甲醇-乙腈溶液(体积比1∶2),设置流速为1.0 mL/min,柱温为40 ℃,紫外检测器波长338 nm。
1.3.4 面团模拟体系和酸面团中γ-谷氨酰二肽含量的测定
采用面团模拟体系对乳酸菌发酵合成γ-谷氨酰二肽进行初步的验证。面团模拟体系参考Zhao等[10]的方法:将活化后的乳酸菌菌液接种至缓冲盐溶液中,37 ℃培养24 h。其中缓冲盐溶液中含有5 g/L麦芽糖和10 g/L氨基酸混合溶液(谷氨酰、胺谷氨酸、缬氨酸、异亮氨酸和亮氨酸)。为验证乳酸菌菌种是否可自身转化生成γ-谷氨酰二肽,将活化后的乳酸菌菌液接种至无氨基酸底物的缓冲盐溶液中进行分析比较。分别取0 h和发酵24 h后的模拟缓冲溶液,离心,过滤,滤液用于液相色谱-二级质谱分析γ-谷氨酰二肽的含量。
采用C18高效液相色谱柱(150 mm×2.1 mm,2.7 μm),流动相A为0.1%甲酸-水溶液,流动相B为0.1%甲酸-乙腈溶液,洗脱速率0.2 mL/min,设置程序:0~10 min,95%~75% A;10~15 min,75%~0% A;15~25 min,0%~95% A;平衡10 min。采用多重反应监测的方法对γ-谷氨酰二肽进行检测分析,4 种目标物质离子对参数见表1。将标准品溶于甲醇-水-甲酸溶液(30∶69.9∶0.1,V/V)中,用于标准曲线的绘制与计算。其中,标准品质量浓度为0.1~100 μg/L。

表1 γ-谷氨酰二肽在液相色谱-二级质谱中测定参数Table 1 LC-MS/MS parameters for detection of γ-GPs
1.3.5 酸面团发酵馒头样品的制备
制备500 g面团,酸面团发酵组和空白对照组配方见表2。将配料置于搅面机中先慢速搅拌4 min,再快速搅拌3.5 min,将成型面团取出,切分成80 g/个,手工搓圆,置于醒发箱内,35 ℃、相对湿度85%条件下发酵50 min。醒发结束后,将面团置于电蒸箱内,100 ℃加热20 min,室温冷却,备用。

表2 馒头制作配方Table 2 Steamed bread recipes
1.3.6 基于电子舌对酸面团发酵馒头滋味特性的测定
根据何晓赟[15]的方法,并适当修改:称取不同乳酸菌发酵酸面团制得的馒头100 g,粉碎,并加入300 mL去离子水,高速匀浆1 min后,置于离心机中3 000×g离心10 min,取上清液。量取上清液100 mL,数据采集中以清洗液(蒸馏水)和待测样品萃取液交替进行,为使传感器响应值趋于平稳,每个样品采集时间为120 s,每隔1 s采集一个数据,采集完毕清洗20 s。为减少测量误差,本试验取最后10 s测量值的平均值作为各传感器的后续处理数据。
1.4 数据处理
研究数据处理均采用Origin 8.0软件分析作图,SAS 8.0进行统计学数据分析,采用ANOVA方法进行显著性差异分析,P<0.05,差异显著。结果均为3 次实验平均值。
2 结果与分析
2.1 不同乳酸菌发酵酸面团的理化特性
由表3可知,不同乳酸菌发酵酸面团48 h后pH值均可达到3.50左右,菌落总数可达到108CFU/g以上。酸面团pH值的降低主要是由于乳酸菌生长代谢产生的乳酸等有机酸成分积累造成[16]。然而,过多的H+和有机酸组分对菌株的生成代谢有强烈的抑制作用[17]。因此,不同乳酸菌在长时间酸面团发酵过程中,pH值和菌落总数在菌种间并无显著性差异。
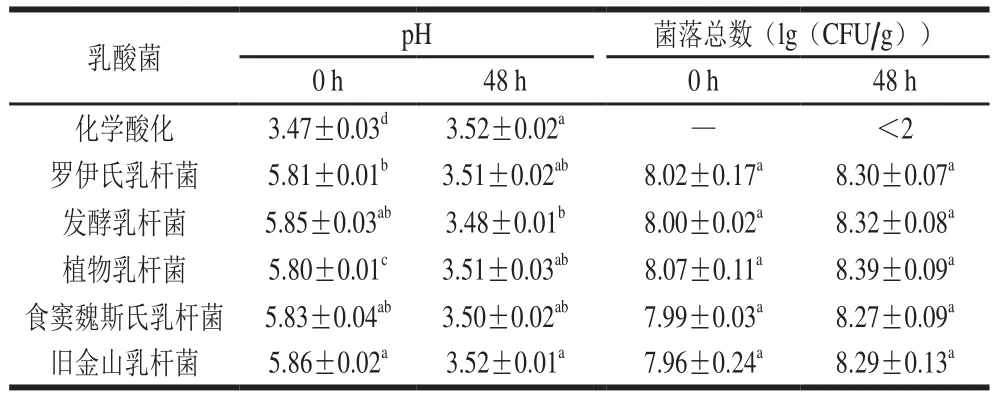
表3 不同乳酸菌发酵对酸面团pH值和菌落总数的影响Table 3 Effect of various lactobacilli on pH and total bacterial count of sourdough
2.2 乳酸菌发酵酸面团对游离氨基酸总量的影响
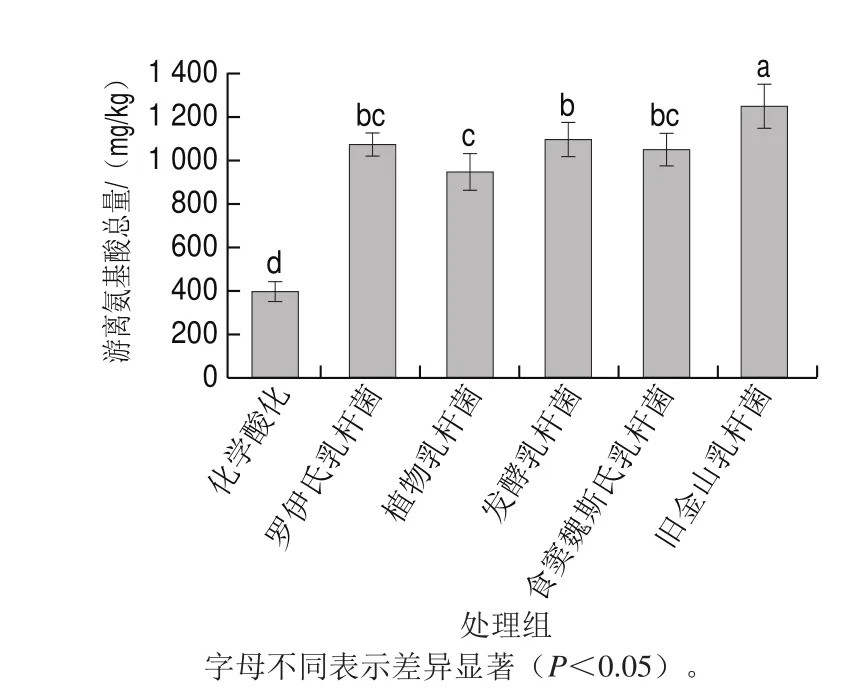
图1 不同乳酸菌发酵酸面团的游离氨基酸总量Fig. 1 Effect of various lactobacilli on the total content of free amino acids in sourdough
如图1所示,与化学酸化相较,乳酸菌发酵可显著提高面团中游离氨基酸总量。在酸面团发酵过程中,由于谷物中的氮源主要以无机形式存在,乳酸菌自身蛋白水解系统无法利用,对此类氮源的降解能力较为有限[18]。其主要是通过乳酸菌代谢产酸创造的低酸环境激活谷物中的内源性蛋白酶,进而促进小麦粉中大分子蛋白聚合物的降解,产生大量的游离氨基酸[19]。乳酸菌发酵酸面团是一个缓释酸化的过程,谷物蛋白酶最适pH 4.0,而化学酸化面团pH值始终保持在3.5左右,因此面团中游离氨基酸总量显著低于乳酸菌发酵组[20]。此外,结果还表明不同乳酸菌发酵酸面团中游离氨基酸总量存在显著差异,这主要是由于乳酸菌对面团中小分子蛋白,如清蛋白、球蛋白和谷蛋白等,具有良好的水解能力[21]。根据本实验结果可推测乳酸菌对小分子蛋白质的水解能力存在种间差异性,进而导致面团中游离氨基酸总量存在显著差异性,这也为γ-谷氨酰二肽的生成提供了物质基础。
2.3 酸面团模式体系下乳酸菌对γ-谷氨酰二肽合成的影响
基于酸面团中游离氨基酸单体含量的比较,本研究选择将γ-谷氨酰谷氨酸、γ-谷氨酰缬氨酸、γ-谷氨酰异亮氨酸和γ-谷氨酰亮氨酸作为γ-谷氨酰二肽代表性目标产物,用于阐明乳酸菌对γ-谷氨酰二肽合成的影响。首先,乳酸菌接种在无游离氨基酸模拟溶液中,γ-谷氨酰二肽并未检出,由此说明乳酸菌在无外源底物的情况下,通过自身代谢无法生成γ-谷氨酰二肽。而由图2可知,不同乳酸菌对γ-谷氨酰二肽的合成存在种间差异性,其中罗伊氏乳杆菌发酵合成γ-谷氨酰二肽含量最高。根据γ-谷氨酰二肽单体合成量的比较可以发现,不同乳酸菌发酵液中γ-谷氨酰谷氨酸和γ-谷氨酰异亮氨酸合成量显著高于γ-谷氨酰亮氨酸和γ-谷氨酰缬氨酸。Zhao等[10]在比较同种不同株水平上罗伊氏乳杆菌对γ-谷氨酰二肽代谢合成的影响中也得到了类似的结果。由此可推测乳酸菌在利用游离氨基酸合成γ-谷氨酰二肽时存在选择性的差异,其通过自身代谢酶系优先选择谷氨酸和异亮氨酸用于相应γ-谷氨酰二肽的合成。针对乳酸菌合成γ-谷氨酰二肽的形成过程,现有研究认为主要与γ-谷氨酰转移酶、γ-谷氨酰半胱氨酸合成酶以及谷胱甘肽合成酶有关[22-23]。γ-谷氨酰半胱氨酸合成酶和谷胱甘肽合成酶均与谷胱甘肽的合成有关,而谷胱甘肽可通过γ-谷氨酰转移酶生成不同的γ-谷氨酰二肽类物质[24]。基于NCBI数据库,将γ-谷氨酰半胱氨酸合成酶调控基因序列与不同种乳酸菌的基因序列进行比对发现,除旧金山乳杆菌以外,本实验所用乳酸菌均含有1~3 个γ-谷氨酰半胱氨酸合成酶调控基因[25]。然而,Kai等[26]的报道显示旧金山乳杆菌中含有谷胱甘肽合成酶,这可能与其代谢合成γ-谷氨酰二肽有重要的关系。
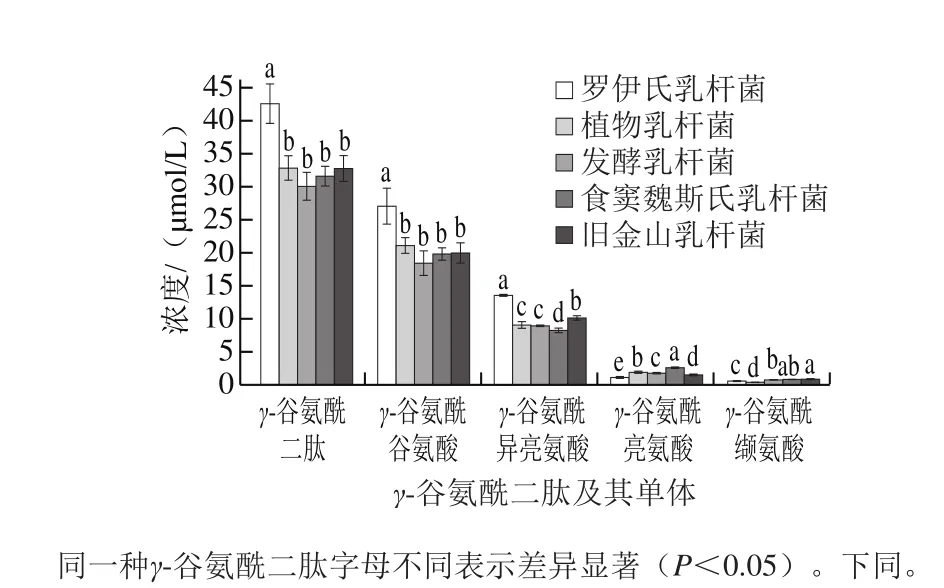
图2 不同乳酸菌发酵对酸面团模拟体系中γ-谷氨酰二肽含量的影响Fig. 2 Effect of various Lactobacillus strains on the concentration of γ-glutamyl dipeptides in buffer-based model dough
2.4 乳酸菌发酵对酸面团中γ-谷氨酰二肽含量的影响
为进一步验证乳酸菌发酵面团对γ-谷氨酰二肽合成的影响,采用不同种乳酸菌发酵酸面团比较发酵过程中γ-谷氨酰二肽的合成量。由表4所示,发酵初始阶段,面团中可测得少量的γ-谷氨酰谷氨酸、γ-谷氨酰异亮氨酸和γ-谷氨酰亮氨酸,而γ-谷氨酰缬氨酸并未检出,这可能与小麦粉中谷氨酸含量较高有关[27]。酸面团经过48 h发酵后(表5),4 种γ-谷氨酰二肽均增加,且γ-谷氨酰谷氨酸和γ-谷氨酰异亮氨酸合成量增加更为显著,这一结果与面团模拟发酵液所得到的结果一致。由γ-谷氨酰二肽总量可知,罗伊氏乳杆菌具有更优的γ-谷氨酰二肽合成能力,再次验证了乳酸菌在酸面团发酵过程中对于γ-谷氨酰二肽的合成存在种间差异性。此外,采用额外添加蛋白酶的方式,通过提高面团体系中游离氨基酸总量,观察4 种γ-谷氨酰二肽含量的变化(数据未列出)。结果表明,游离氨基酸底物浓度的增加可有效促进4 种γ-谷氨酰二肽的合成量,由此说明γ-谷氨酰二肽的合成与发酵过程中蛋白质水解程度有紧密的关系。
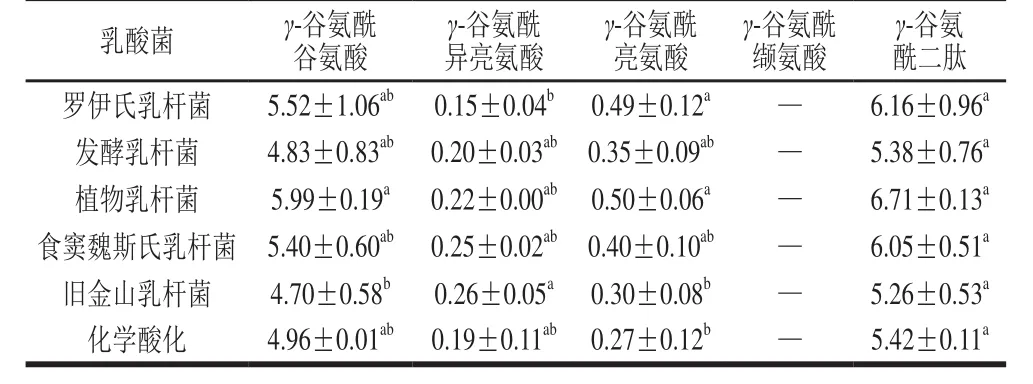
表4 不同乳酸菌发酵0 h对酸面团中γ-谷氨酰二肽浓度的影响Table 4 Effect of various Lactobacillus strains on the concentration of γ-glutamyl dipeptides in unfermented sourdough μmol/L
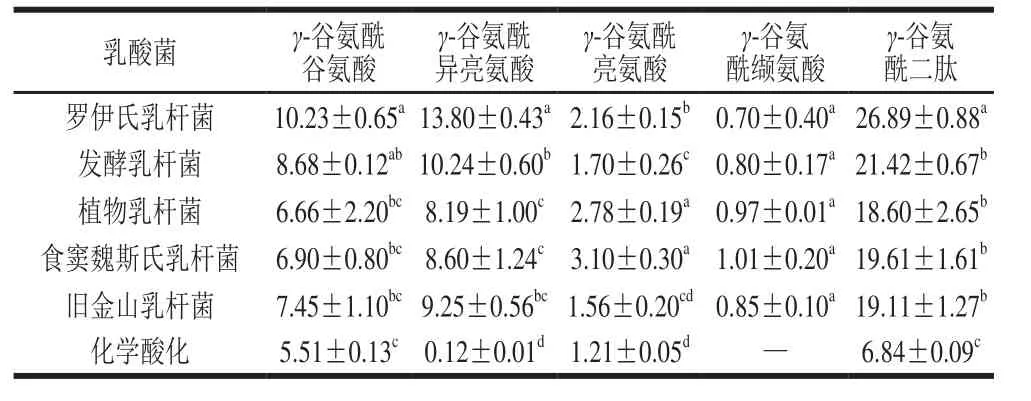
表5 不同乳酸菌发酵48 h对酸面团中γ-谷氨酰二肽浓度的影响Table 5 Effect of various Lactobacillus strains on the concentration of γ-glutamyl dipeptides in 48 h fermented sourdough μmol/L
2.5 乳酸菌发酵酸面团制备馒头样品的电子舌滋味分析
研究表明,具有浓厚味的活性物质可有效提高产品的其他滋味强度,如酸味、鲜味和咸味等,进而显著改善提升产品的滋味特性。为进一步验证乳酸菌发酵酸面团代谢合成γ-谷氨酰二肽对馒头滋味特性的影响,研究采用电子舌对不同乳酸菌发酵酸面团制得馒头的滋味特性进行分析比较,由图3可知,与空白对照组相较,乳酸菌发酵酸面团制得馒头在酸味、鲜味和咸味上存在显著差异。乳酸菌发酵可代谢积累乳酸等有机酸成分,因此制得产品相较空白组酸味显著增强。其中,由于植物乳杆菌生长速率快,产酸能力较强,因此其制得产品酸味值较高[28]。针对咸味方面,由于产品配方中并未添加任何咸味成分,其咸味可能与发酵过程中代谢形成的咸味氨基酸或小分子肽有关[29-30]。然而,通过与空白组比较,结合不同乳酸菌发酵酸面团对γ-谷氨酰二肽合成量的分析发现,罗伊氏乳杆菌发酵过程中γ-谷氨酰二肽合成量最高,其制得产品咸味值也是最强的。由此推断,γ-谷氨酰二肽与馒头咸味的增强有一定的联系。随着消费者对低钠摄入健康饮食的不断推崇,这为乳酸菌发酵高产γ-谷氨酰二肽制备低钠发酵食品奠定了理论基础,也为米面发酵食品优质菌种的筛选提供了理论依据。
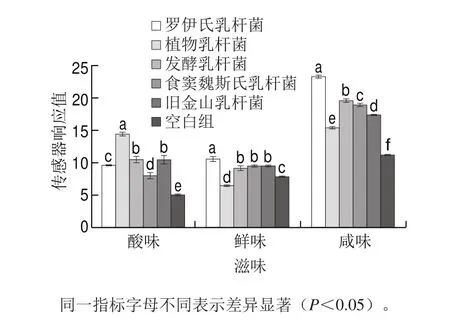
图3 不同乳酸菌发酵酸面团制得馒头滋味特性的比较Fig. 3 Effect of sourdough fermented by various Lactobacillus strains on the taste of steamed bread
3 结 论
研究表明,不同乳酸菌发酵酸面团均可通过自身代谢合成γ-谷氨酰二肽,但在发酵过程中对合成底物具有优先选择性,且其代谢合成量种间存在显著差异。在酸面团常见发酵菌种中,罗伊氏乳杆菌合成γ-谷氨酰二肽能力最优。此外,旧金山乳杆菌虽然不含γ-谷氨酰半胱氨酸合成酶,但仍可代谢合成γ-谷氨酰二肽,说明γ-谷氨酰二肽的代谢调控可能与γ-谷氨酰半胱氨酸合成酶和谷胱甘肽合成酶有关。电子舌分析数据表明,γ-谷氨酰二肽可显著提升产品的咸味强度,有效提升产品的滋味特性,这也为酸面团发酵制得馒头产品风味浓郁提供了新的理论佐证。
