不同土壤基质下水分胁迫对黄檗幼苗形态和物质分配的影响
2019-09-05李绍臣杨雨春陆志民
及 利 ,李绍臣 ,王 君 ,王 芳 ,杨雨春 ,陆志民 ,孙 伟
(1.吉林省林业科学研究院,吉林 长春 130033;2.东北林业大学 林学院,黑龙江 哈尔滨 150040)
气候变化模型预测未来十几年内,许多地区可能会出现严重的极端气候事件,如干旱、热浪和洪水等,出现土壤水分亏缺和发生短期干旱的范围和频率将翻倍[1-2]。水分是影响植物生长的重要因素之一,缺水对植物生产的负面影响已成为林业可持续发展的一个主要问题[3-4]。因此,在苗木培育过程中,研究幼苗对水分亏缺的形态学、生理学等方面的响应机制变得愈发关键。
土壤水分是影响植物生长发育和形态建成的重要因子之一[5],土壤水分不足会延缓或破坏植物的正常生长,改变植物的形态和生物量分配[6-7],并最终导致植物死亡[5,8]。安玉艳等[9]发现,水分胁迫下的杠柳Periploca sepium生长减慢,总生物量降低,且生物量优先分配到根系生长,根冠比显著升高。徐飞等[10]认为,水分胁迫虽然对刺槐Robinia pseudoacacia幼苗的径向生长没有显著影响,但明显限制了刺槐幼苗的纵向生长,这种降低株高与基径比例的适应策略有利于刺槐获得更多的光能。但目前关于水分胁迫方面的研究多集中于单一土壤基质上,由于不同土壤基质间的容重、孔隙度和养分等方面存在一定的差异[11],因此探究不同基质条件下幼苗对水分亏缺的响应,对建立苗木栽培过程中建立精准的培育和管理措施提供依据。
黄檗Phellodendron amurense作为中国东北三大硬阔之一,是东北阔叶红松林的主要伴生树种之一,既是珍贵的用材树种也是重要的药材,具有较高的经济和生态价值[12]。但目前针对黄檗的研究多集中于其药用成分等方面[13-16],而在苗木栽培方面的研究较少,尤其是关于不同育苗基质方面。因此,本研究以3年生黄檗幼苗为研究对象,通过比较不同土壤基质间幼苗的形态建成和物质分配在面临水分胁迫时的变化,揭示黄檗幼苗对干旱胁迫的适应策略,以期为该树种培育过程中的基质选择和水分管理提供依据。
1 研究区概况与研究方法
1.1 研究区概况
研究区位于吉林省长春市净月开发区新立城镇(43°33′~ 44°41′N,125°19′~ 125°24′E),该地区气候属温带大陆性季风气候,年平均温度为4.6 ℃,无霜期140~150 d,年平均降水量为600~800 mm。为避免降水对幼苗干旱胁迫造成的影响,在该地区设置人工遮雨棚,遮雨棚两侧可打开通风。
1.2 试验材料
以吉林省露水河宏伟苗圃3年生黄檗幼苗为实验材料,于2017年4月中下旬移栽到砖红色塑料盆中(17 cm×12 cm),置于遮雨棚内进行缓苗。分别向盆中装入等体积的腐殖土、壤土和砂壤土(壤土:沙=1:1)3种土壤基质,其中腐殖土采于针阔混交林地,土壤类型为暗棕壤。
1.3 试验设计
2017年7月,取3种土壤基质样品,分别测定其全氮、全磷、速效磷和土壤物理性质(表1),随即选取生长一致的黄檗幼苗分别栽植到不同基质的盆中进行控水试验。以土壤基质和水分进行两因素完全正交试验设计,3种土壤基质分别设置4个水分胁迫梯度,水分梯度为CK(正常供水,80%田间持水量)、T1(轻度胁迫,60%田间持水量)、T2(中度胁迫,40%田间持水量)和T3(重度胁迫,20%田间持水量),共计12个处理,每个处理设置3个小区,每个小区20盆黄檗幼苗(选取长势均一的苗木),控水方式为整盆称重法,每小区20盆,计算出理论上不同处理的土、盆、苗和该水分下的总质量,隔天进行差额补水。每20 d取样1次,共取3次(取样时间分别为8月8日、8月29日和9月20日)。由于在胁迫过程中幼苗自身质量在不断变化,在每次取样时再次测定幼苗总质量,校正理论上的整盆总质量后再继续进行差额补水。
1.4 测定方法
在水分胁迫前分别测定两种土壤基质的土壤理化性质,土壤全氮用凯氏滴定法测定,土壤全磷用硫酸-高氯酸-钼锑抗比色法[17],土壤有效磷用双酸浸提法,土壤含水量和容重用环刀法测定[18],土壤总孔隙度、通气孔隙度、吸水倍数、渗水速率和蒸发速率参照卫星的方法测定[11]。

表1 3种土壤基质的理化性质†Table1 Physicochemical properties of three soil substrates
每次取样选取5盆幼苗测定苗高、地径和冠幅等指标,然后全株挖出后分开根、茎、叶3个器官取样装于信封中。每株叶片平铺于刻度纸上并用Epson Scanner扫描仪扫描,再用Photoshop CS 5.1软件进行计算总叶面积[19]。叶片扫描后,与根和茎放入变温烘箱中于105 ℃杀青30 min,之后调为85 ℃烘干至恒质量,测定各器官生物量。所有指标的取样均为5重复。由下列公式计算叶面积指数(LAI)、比叶面积(SLA)、根冠比(R/S)和各器官生物量比:
叶面积指数(LAI)=总叶面积/冠面积;
比叶面积(SLA)=叶面积/叶干重;
根冠比(R/S)=根生物量/(地上生物量);
根(茎、叶)生物量比(RMR、SMR和LMR)=根(茎、叶)生物量/总生物量。
1.5 数据分析
采用Microsoft Excel 2010整理数据;利用SPSS19.0进行单因素方差(One-way ANOVA)和多因素方差(Multi-way ANOVA)分析检验不同处理间的差异(LSD,α=0.05);叶面积用Adobe Photoshop CS 5.1计算;用Canoco for Windows 5.10软件对不同处理下的幼苗形态和生物量分配等指标进行主成分分析(Principal component analysis,PCA)和冗余分析(Redundancy analysis,RDA)。在Sigmaplot 12.5软件中作图。所有数据为平均值±标准误(Mean±SE)。
2 结果与分析
2.1 水分胁迫对黄檗生长指标的影响
在水分胁迫60 d后,3种土壤基质中,黄檗幼苗的苗高、地径和冠面积均随着水分胁迫强度的增加而呈显著降低的趋势(表2,P<0.05)。3种基质中,腐殖土中的幼苗的苗高、地径和冠面积在胁迫中变化最大,T3处理下的苗高、地径和冠面积分别比CK降低了21.05%、23.93%和31.02%(P<0.05),而T1处理下的苗高、地径和冠面积与CK的无显著影响。幼苗总叶面积大小随着水分胁迫程度的加深而降低,与CK相比,腐殖土、壤土和砂壤土在T3处理下的叶面积分别降低了15.65%、58.64%和69.87%(P<0.05)。
2.2 水分胁迫对黄檗生物量分配的影响
整个60 d胁迫周期内,水分胁迫对叶片生物量和总生物量有显著影响,幼苗叶片生物量和总生物量随胁迫强度的增加而呈逐渐降低的趋势(图1,P<0.05)。在胁迫60 d时,水分和土壤基质对黄檗幼苗各器官生物量均存在显著影响(P<0.05),其中,在腐殖土、壤土和砂壤土中,T3处理下的叶片生物量比CK降低了59.94%、38.65%和41.58%。幼苗茎生物量在胁迫20 d时受土壤和水分影响较小,3种土壤基质下各胁迫处理对茎生物量没有显著影响,但在胁迫60 d时,水分亏缺显著降低了茎生物量。壤土和砂壤土中,在胁迫20 d时水分胁迫对幼苗根系生物量不存在显著影响,随着胁迫时间的延长,胁迫40 d和60 d时,T1处理下的根系生物量明显高于CK处理,说明适当的胁迫促进了根系的生长发育以达到获取更多水分的目的。

表2 胁迫60 d后不同土壤基质和水分胁迫下幼苗生长指标的变化Table2 Changes of growth indexes of seedlings under different soil substrates and water stress after 60 days of stress
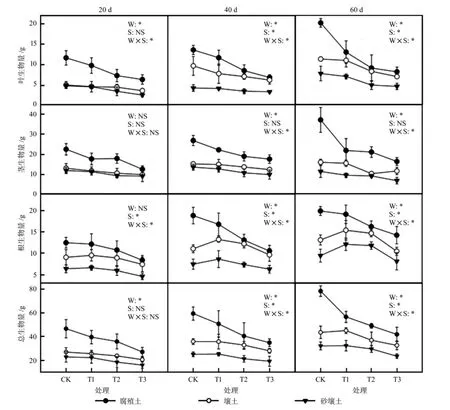
图1 不同土壤基质和水分胁迫下幼苗生物量分配的变化Fig.1 Changes of biomass allocation in seedlings under different soil substrates and water stress
水分胁迫和土壤基质显著改变了叶生物量比、根生物量比和根冠比(表3),其中,3种土壤基质中T2处理的叶片生物量比平均比CK降低了24.97%,而砂壤土土中T2处理的叶片生物量比较CK处理下降幅度最大,为32.08%(P<0.05)。经过60 d胁迫处理,幼苗根生物量比有明显提高,3种基质中T2处理下的根生物量比均显著高于CK(P<0.05),而腐殖土中T3处理的根生物量比最高,壤土和砂壤土中T2处理的根生物量比最高,说明不同土壤基质下幼苗对胁迫的耐受程度不同,壤土和砂壤土中的幼苗在胁迫一定程度后,根生物量分配不再继续增加。
随着胁迫程度的加深,腐殖土中幼苗根冠比呈逐渐增加的趋势,而壤土和砂壤土则是呈先增加后降低的趋势,在T2处理下达到最高(表3,P<0.05)。胁迫处理后,砂壤土中幼苗根冠比变化最大,T2处理比CK高59.50%(P<0.05)。各水分处理下,幼苗根冠比在3种土壤基质的顺序为:腐殖土<壤土<砂壤土,说明土壤物理结构和养分对根冠比影响较大。
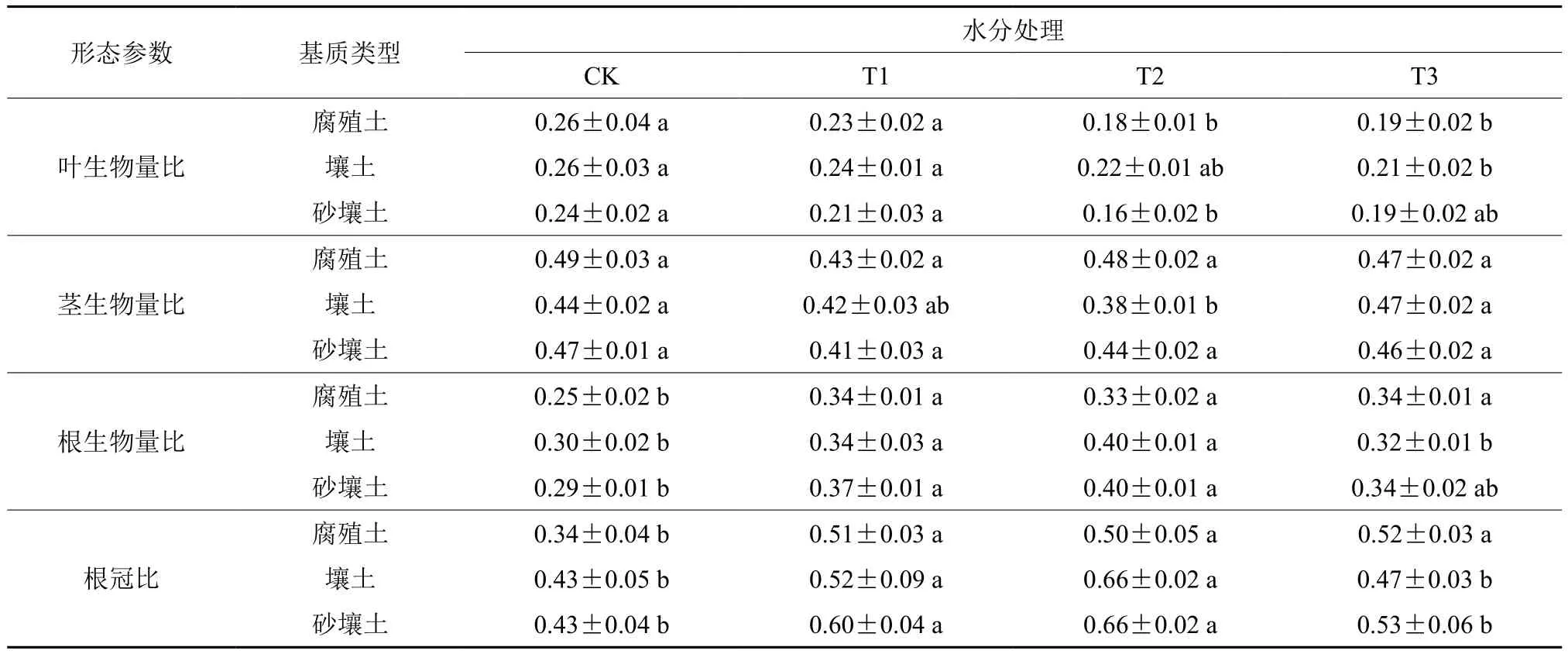
表3 不同水分胁迫条件下幼苗生物量的分配Table3 Allocation of seedling biomass under different water stress conditions
2.3 水分胁迫对黄檗比叶面积的影响
腐殖土中幼苗比叶面积(SLA)随胁迫程度加深而呈升高的趋势,胁迫60 d后,T3处理的SLA比CK高110.55%(图2)。总体上,在胁迫20 d后,壤土和砂壤土中幼苗SLA随胁迫程度加深而呈降低的趋势,同时这两种土壤基质下的T1和CK处理之间未达到显著水平,T2和T3处理下的SLA显著低于CK(P<0.05)。同时,3种土壤基质的SLA均随胁迫时间的延长而降低,且在胁迫60 d时最低,说明幼苗叶片对胁迫环境存在一个适应的过程,中度和重度胁迫显著改变了幼苗的SLA。

图2 不同土壤基质和水分胁迫下幼苗比叶面积的变化Fig.2 Changes of specific leaf area of seedlings under different soil substrates and water stress
2.4 土壤基质与幼苗形态建成的因子分析
3种土壤基质中的幼苗形态学和生物量分配参数表现出不同的变异程度,土壤基质和水分条件改变了幼苗的形态建成(图3)。两个主成分轴分别解释了总变异的83.2%和6.6%。各器官生物量与主成分一轴显著相关,叶面积指数(LAI)与主成分二轴显著相关。3种土壤基质的不同水分处理典型地分布在主成分轴的3个象限内,同时幼苗的形态和生物量分配等指标与腐殖土保持较高的正相关关系,与壤土和砂壤土则是明显的负相关。

图3 不同水分胁迫处理下3种土壤基质中黄檗幼苗生长的主成分分析Fig.3 Principal component analysis of growth of Phellodendron amurense seedlings in three soil substrates under different water stress treatments
形态建成的相关关系,两个排序轴共解释了总变异的90%以上(图4),把3种土壤基质下幼苗的生长形态指标与土壤理化指标进行偏蒙特卡洛置换检验,土壤蒸发速率(Reva)、全氮和全磷是第一排序轴的主要影响因子,其中,土壤全N、全P、蒸发速率和渗透速率与幼苗形态和生物量存在明显的正相关关系,与根冠比呈负相关关系,而土壤容重与幼苗形态之间呈负相关关系,与根冠比呈正相关关系。
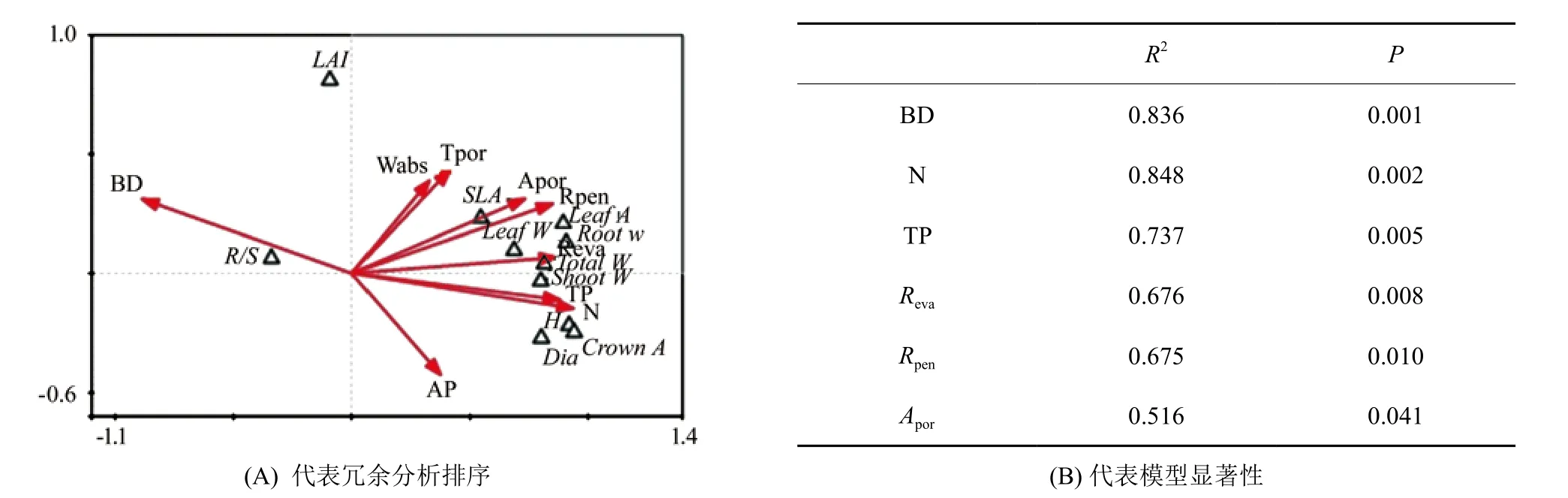
图4 黄檗幼苗生长和土壤因子的冗余分析Fig.4 The redundancy analysis of Phellodendron amurense seedling growth and soil factors
3 结论与讨论
土壤水分是影响植物生长发育和形态建成的重要因子之一,而土壤容重、孔隙度和土壤养分等土壤理化性质在影响着幼苗的形态学特征和物质的分配等方面同样发挥着重要的作用[5,11]。目前普遍认为,当育苗基质总孔隙度为70%~90%,通气孔隙度大于15%,密度为0.2~0.6 g·cm-3,pH值为6.0~7.5时,符合大多数植物的生长要求[20]。本研究中选取的3种土壤基质间的土壤理化性质存在显著差异(表1),土壤基质对黄檗幼苗的苗高、地径、冠面积和叶面积有显著的影响,总体上表现为腐殖土最好、壤土次之。同时,冗余分析的结果表明,土壤容重、蒸发速率和渗透速率是影响幼苗的根冠比和各器官生物量分配等指标的重要因素(图4)。卫星等[11]研究表明,不同土壤物理性质下的育苗基质对容器苗的生长指标存在显著影响,一定范围内较大的孔隙度利于根系的生长,但孔隙度过大不利于保水保肥的实现。这进一步说明了在研究水分亏缺对幼苗影响的同时,选择合理的土壤基质是至关重要的。
在面对水分亏缺时,植物通常通过调整自身生长和生物量分配来适应环境条件的变化,水分胁迫过程中幼苗的物质分配方式一定程度上体现着其生存对策[6,21-23]。单立山等[24]发现,适度的干旱处理增加了红砂幼苗根系总生物量、各级侧根生物量和主根生物量,以扩大其营养空间适应胁迫环境。本研究中,胁迫初期水分条件对幼苗茎和根的生物量没有显著影响,在胁迫40 d后,水分显著影响了幼苗各器官生物量,而幼苗叶片生物量在整个胁迫期间内都与胁迫强度存在显著关系,同时3种土壤基质中幼苗根冠比均随胁迫的加剧而升高,说明3种土壤基质下的黄檗幼苗在面临水分胁迫时,都是通过降低叶片生物量增加地下生长的行为,以达到减少水分消耗,逃避和耐受环境条件造成的胁迫伤害。
在水分亏缺条件下,由于植物为达到减少自身蒸腾面积,降低水分丧失等目的,水分胁迫降低了幼苗叶片数量、总叶面积和平均叶面积[25-26]。但在降低的叶面积的同时也损失了对光合作用产生的碳水化合物的获取,因此,比叶面积(SLA)成为评价水分胁迫下植物适应策略的重要指标[26-27]。本研究中,随胁迫时间延长,3种土壤基质下的幼苗SLA均呈逐渐降低的趋势,砂壤土中的幼苗SLA随胁迫强度增加而降低,而腐殖土的则是随胁迫增加而逐渐升高,说明腐殖土条件下幼苗叶片生物量丢失的速度要高于叶面积减小的,虽然胁迫条件下腐殖土的叶生物量降低,但未对其获取碳水化合物造成阻碍,降低叶片建成的投资成本,同时也提高了对碳的利用效率。
综上所述,水分胁迫显著降低了黄檗幼苗的叶面积、叶片生物量和总生物量,土壤全N、全P、蒸发速率、渗透速率和容重是影响黄檗幼苗生长和物质分配的重要因素。腐殖土条件下(即土壤容重、孔隙度和养分等条件合理的基质)适度的水分亏缺,有利于黄檗幼苗自身的形态建成,没有限制幼苗对碳水化合物的获取。本研究是基于水分和栽培基质两个因素探讨不同基质条件下水分胁迫强度对幼苗黄檗幼苗生长和物质分配的影响,还需结合生理方面以及根系和叶片对胁迫的协同变化机制进行综合地评价,同时,由于野外条件复杂,还需结合光照强度、干旱后复水以及淹水胁迫等试验完善黄檗对不同土壤基质和水分条件下的变化机制,以期为黄檗的培育、管理提供依据和理论支撑。
