中国近海棱梭群体形态学和耳石形态学研究
2019-08-30吴天昊谭颖琪龙丹丹高天翔
阳 敏,刘 璐,吴 桢,吴天昊,谭颖琪,龙丹丹,高天翔,宋 娜
(1.中国海洋大学水产学院,山东 青岛266003;2.浙江海洋大学水产学院,浙江 舟山316022)
形态学方法是鱼类群系划分的一种传统的、基本的方法[1],早期的形态学研究主要对样品的量度特征和分节特征等形态学指标进行统计分析[2]。在上世纪80年代出现的地标形态学则借助几何学方法对鱼体的二维或三维形态坐标点间的位置、距离等关系进行分析[3],两种方法结合使用可以更好地研究形态学差异[4]。硬骨鱼类矢耳石在形成过程中其形态结构十分稳定,且具有物种和群系特异性[5-6]。随着图像分析技术的进步,耳石形态被越来越广泛地应用于种内和种间的鉴别[7-9]。鱼类耳石外部形态特征的分析方法通常有传统耳石形态分析法和椭圆傅里叶分析法两种[2]。前者侧重对耳石体轴的分析,即对耳石形态量度特征(如耳石长、耳石宽、耳石面积等)或者将其进行几何学组合得到的形态指标(如椭圆率、矩形趋近率、半径比等)进行分析[10-11];后者则侧重对耳石外部轮廓进行分析[12-14]。
棱梭(Chelon affinis)隶属硬骨鱼纲,鲻形目,鲻科,梭属(Chelon)[15],作为梭属鱼类的一个重要鱼种,因其背部在第一背鳍之前有一条正中纵走棱脊而明显区别于其他梭属鱼类而得名。此外该鱼类国内又称棱鮻[16]、前鳞龟鮻[17-18]等;拉丁学名Liza affinis[16]、Chelon affinis、Planiliza affinis[19]等都作为棱梭的学名出现,本研究采用目前国内外普遍使用的Chelon affinis作为棱梭的学名。棱梭栖息于浅海或河口咸淡水交汇处,淡水至盐度达38的海水中均能生长。棱梭的耐寒能力较强,性活泼,春季游向近岸,冬季于深水处越冬。性成熟鱼年龄为3龄,产卵期为冬末春初,是重要的经济鱼类之一[20]。国内对与棱梭同为梭属的梭鱼(Chelon haematocheilus)的群体形态学研究已于2012年被报道[21],但关于棱梭的形态学和耳石形态学研究目前未有报道。本研究从形态学、耳石形态学两个角度对采自中国近海海域6个地点的棱梭群体的差异进行分析,以阐述棱梭种内不同地理群体间的分化水平,为进一步的棱梭群体遗传学研究提供参考,同时丰富鱼类群体耳石形态学的研究内容,比较形态学和耳石形态学两种方法在群体差异研究中的效果。
1 材料与方法
1.1 实验材料
本研究于2013年10月至2015年11月分别采集了金门、厦门、虎门、湛江、北海、防城港各邻近海域中共6个棱梭地理群体的样品(图1、表1)。样品由碎冰包埋处理后运输到中国海洋大学渔业生态学实验室,选取性成熟个体经形态学种类鉴定[22]后进行形态学测量,并摘取样品的左右矢耳石。

图1 棱梭样品采集站位分布Fig.1 Distribution of the sampling sites for Chelon affinis
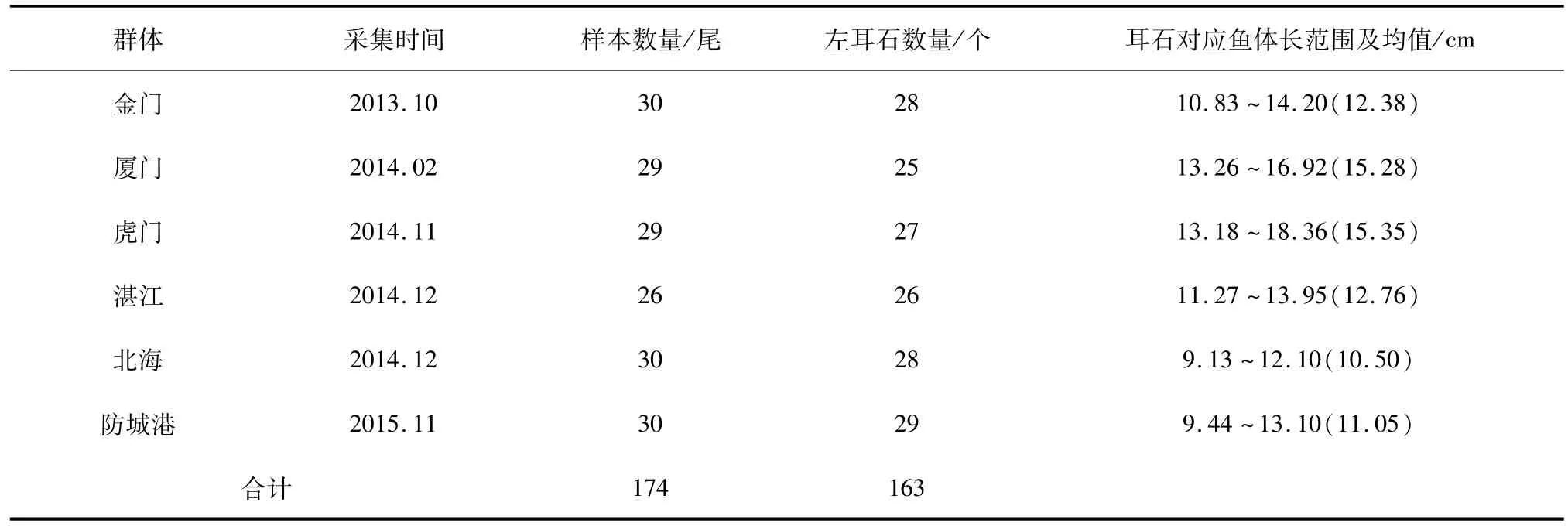
表1 棱梭样品及矢耳石采集信息Tab.1 Sampling information of C.affinis and their sagittal otoliths
1.2 实验方法
测定棱梭的14个可数特征(第一背鳍鳍棘数量、第二背鳍鳍棘数量/鳍条数量、胸鳍鳍棘数量、腹鳍鳍棘数量/鳍条数量、臀鳍鳍棘数量/鳍条数量、尾鳍鳍条数量,上/下鳃耙数量、脊椎骨数量、横列鳞数量、纵列鳞数量)和16个可量特征(体重、体高、全长、体长、第一背鳍基长、第二背鳍基长、胸鳍长、腹鳍长、臀鳍基长、头长、吻长、上颌长、眼径、眼间隔、尾柄长、尾柄高)。此外,选取棱梭外部形态的9个坐标点构建框架结构(图2)。根据设定的9个坐标点之间的直线距离,测量两两之间16个量度指标:D1-2(坐标点1与2之间的距离,以下类推)、D1-3、D2-3、D2-4、D2-5、D3-4、D3-5、D4-5、D4-7、D5-6、D6-7、D6-8、D6-9、D7-8、D7-9、D8-9。所有可数特征均由肉眼直接计数,体重由电子天平秤得(精确到0.1 g),其他量度指标均由游标卡尺测得(精确到0.01 cm)。
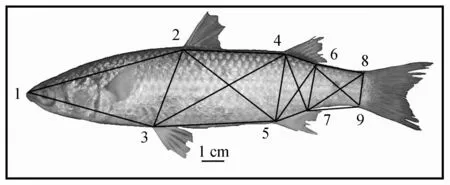
图2 棱梭框架结构测量示意Fig.2 Truss network for morphometrics measurements of C.affinis
将摘取的左右矢耳石置于已编号的离心管中,经超声波清洗,烘箱烘干[23]。用连于解剖镜的数码相机Nikon SMZ 1000对所有耳石的凹面进行拍照。使用Image-Pro Plus 6.0对获得的耳石照片进行校准、测量,得出8个耳石尺寸指标:耳石面积(A)、耳石周长(P)、耳石长(L)、耳石宽(W)、耳石最大Feret径长(Fmax)和最小Feret径长(Fmin)、耳石最大半径(Rmax)和耳石最小半径(Rmin)[23]。利用以上8个耳石尺寸指标得到8个耳石形状指标:椭圆率=(L-W)/(L+W);形态因子=4πA/P2;矩形趋近率=A/(L×W);环率=P2/A;圆度=4A/πL2;幅形比=L/W;耳石的Feret比=Fmax/Fmin;耳石的半径比=Rmax/Rmin[11,23]。耳石大小、耳石摆放位置不同等因素会对分析结果产生影响,这可以通过耳石形状指标消除[11]。
椭圆傅里叶分析方法侧重于耳石二维轮廓和形态分析[24]。首先应用Shape软件包[25]对耳石二维影像进行处理得到傅里叶谐函数,而后依次使用Chaincoder.exe、CHC2NEF.exe、Prin Comp.exe软件包将其转换为傅里叶谐值,得到77个椭圆傅里叶参数[24]。
1.3 数据分析
1.3.1 形态学数据分析 采用体长比值法(头部指标量度值除以头长、尾柄长除以尾柄高、其他参数值除以体长)[26-27]对测量数据进行校正以消除非同龄棱梭不同体长所带来的差异。
①主成分分析:对28组校正后的形态学量度指标进行主成分分析,根据前两个主成分得分绘制主成分散点图。
②聚类分析:采用组之间联接的方式,使用平方Euclidean距离系数对28组校正后的形态学量度指标平均值和4组可数特征的平均值进行聚类分析,构建聚类关系树。
③判别分析:对28组形态学量度指标进行判别分析,根据前两个判别函数值制成散点图。
④单因子方差分析:采用LSD法和Tamhane’s T2法对28组校正后的形态学量度指标进行单因子方差分析。
以上分析均通过SPSS 22.0软件完成。
1.3.2 耳石形态数据分析 使用SPSS 22.0软件对棱梭的8个耳石形状指标和77个椭圆傅里叶参数共85个耳石形态指标进行主成分分析、聚类分析和判别分析,具体方法参照“1.3.1”。
2 结果与讨论
2.1 棱梭形态学结果
对6个群体的14个可数特征进行分析(表2),结果显示:6个群体的腹鳍鳍棘数量、腹鳍鳍条数量、第一背鳍鳍棘数量、第二背鳍鳍棘数量、臀鳍鳍棘数量、臀鳍鳍条数量、尾鳍鳍条数量均为固定值,分别为1、5、4、1、3、9、14;纵列鳞数量介于26~43片之间,数值变化范围较大,分布相对较为平均。横列鳞数量介于10~17片之间,大多群体数量在11~14片之间,占总数的87.4%;胸鳍鳍棘数量在14~17条之间,大多群体数量在15~16条之间,其中以15条最多,占总数的76.9%;第二背鳍鳍条数量介于6~10条之间,大多群体数量为8条和9条,二者占总数的91.4%;上鳃耙数量介于25~28根之间,下鳃耙数量介于46~49根之间,分布均相对较为平均;脊椎骨数量介于22~25节之间,24节的个体最多,占总数的65.7%。不同棱梭群体之间的可数特征差异十分微小。

表2 棱梭群体可数特征Tab.2 Countable characteristics of C.affinis populations
使用体长校正法校正所有棱梭样本的传统形态学可量特征数据和框架数据,得到28个量度指标,对其进行主成分分析。量度指标包括体高/体长、第一背鳍基长/体长、第二背鳍基长/体长、胸鳍长/体长、腹鳍长/体长、臀鳍基长/体长、头长/体长、吻长/头长、上颌长/头长、眼径/头长、眼间隔/头长、尾柄长/尾柄高、D1-2/体长、D1-3/体长、D2-3/体长、D2-4/体长、D2-5/体长、D3-4/体长、D3-5/体长、D4-5/体长、D4-7/体长、D5-6/体长、D6-7/体长、D6-8/体长、D6-9/体长、D7-8/体长、D7-9/体长、D8-9/体长。将主成分根据其特征值大小排序,结果显示前两个主成分的贡献率分别为27.623%和10.429%,特征值大于1的前8个主成分累积贡献率为65.868%(表3)。第一主成分中框架数据载荷值较大,主要反映鱼体比例特征;第二主成分中胸鳍长/体长、腹鳍长/体长、头长/体长、D6-8/体长和D7-9/体长的载荷值较大,反映鱼体胸鳍长、腹鳍长、头长和尾柄长等特征。
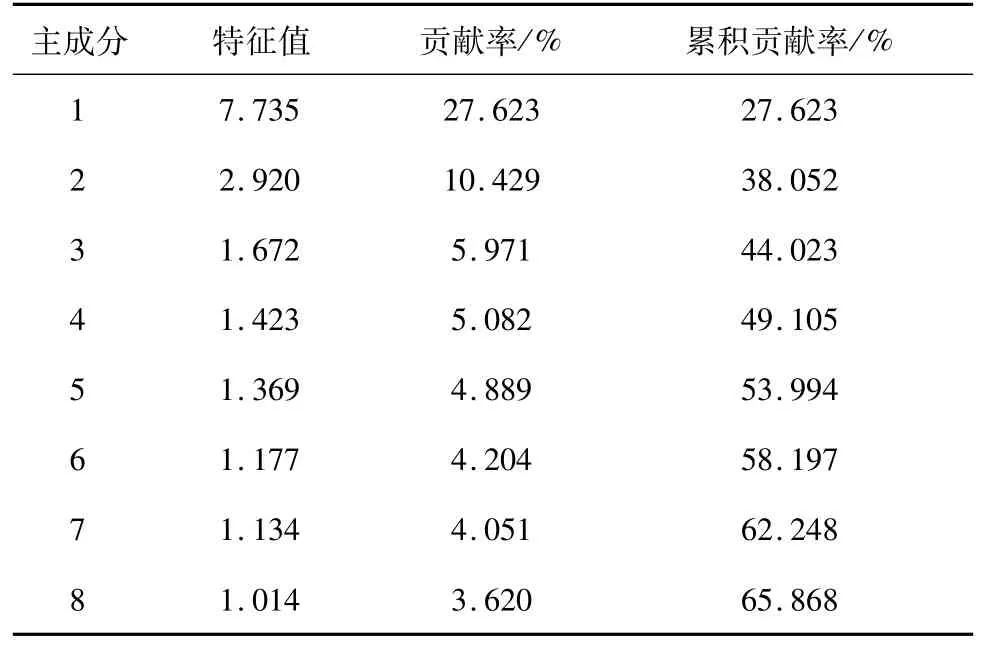
表3 前8个主成分的特征值和贡献率Tab.3 Eigenvalues and contributions of the first 8 principle components
根据第一主成分和第二主成分分析得分绘制散点图(图3),图3中显示,各群体有较大交叉,北海群体多位于散点图纵轴的正半轴,厦门群体多位于散点图纵轴的负半轴,其他群体则没有呈现出明显的分布倾向。
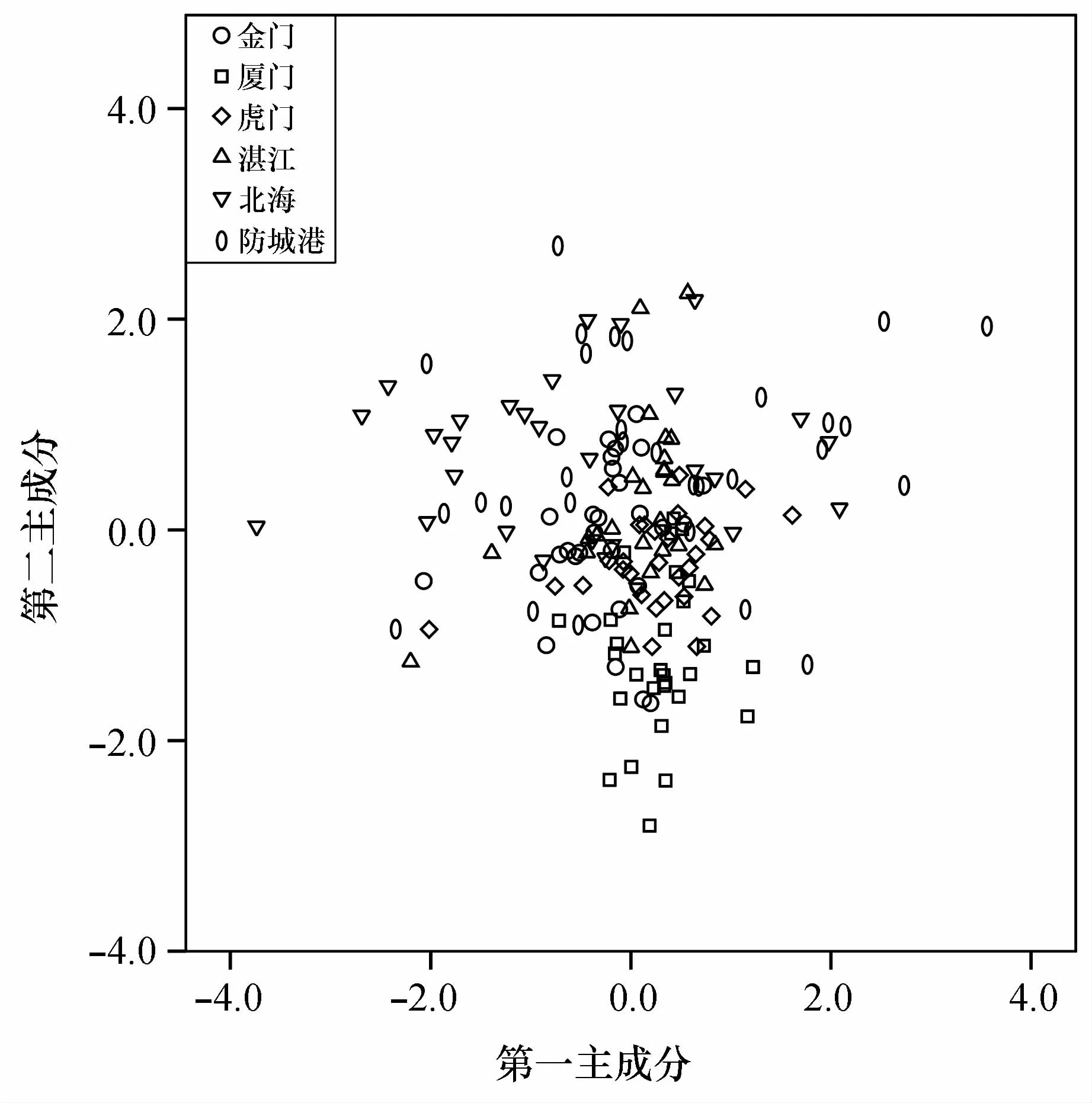
图3 形态量度指标的前两个主成分得分散点图Fig.3 Scatter plots based on scores of the first two metrical characteristics from PCA
对棱梭的28个量度指标和4个可数特征(胸鳍鳍条数量、第二背鳍鳍条数量、纵列鳞数量和脊椎骨数量)的平均值进行聚类分析,构建平均联接的树状图(图4)。图中显示金门群体和北海群体形态特征上十分相近,首先聚为一支,然后依次与虎门、湛江、厦门和防城港聚在一起。
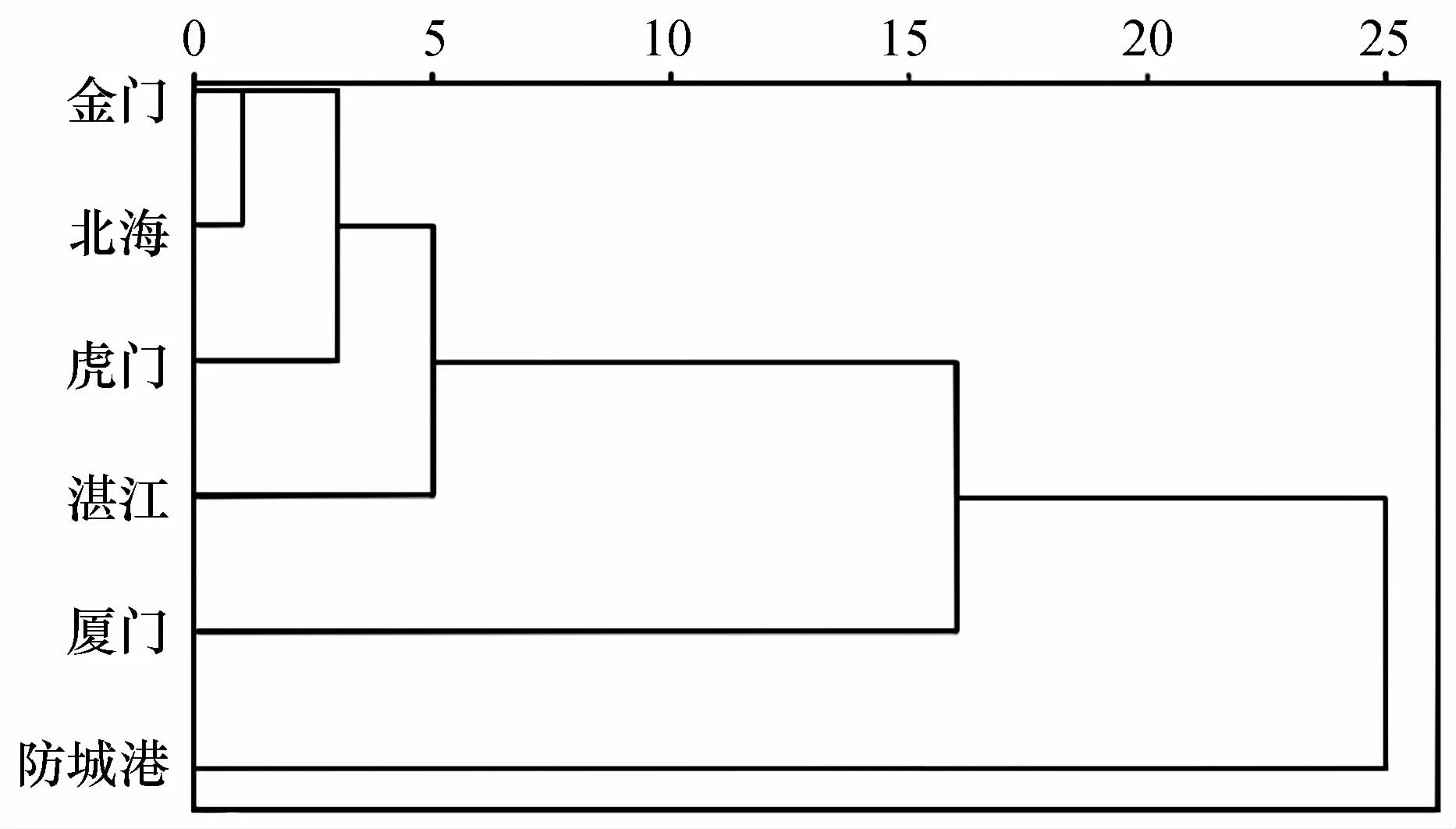
图4 基于形态特征的6个棱梭群体聚类关系Fig.4 Dendrogram of the relationship of six C.affinis populations based on morphological characteristics
对6个棱梭群体的28个量度指标进行判别分析,结果显示,6个群体的判别正确率在57.7%~93.1%之间,其中厦门群体判别正确率最高,为93.1%,湛江群体判别正确率最低,为57.7%,综合判别正确率为75.9%(表4)。判别分析过程中,为了建立判别公式而从28个指标中筛选出对判别贡献较大的9个参数,分别为体高/体长(A)、第二背鳍基长/体长(B)、腹鳍长/体长(C)、臀鳍基长/体长(E)、头长/体长(F)、上颌长/头长(G)、D1-2/体长(H)、D2-3/体长(I)、D7-9/体长(J)。所得判别正确率Y计算公式如下:
金门群体:
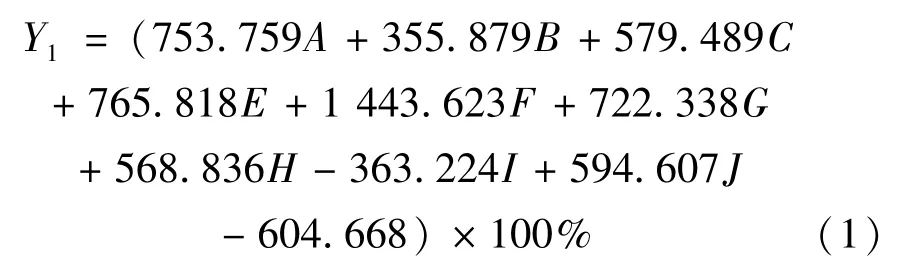
厦门群体:

虎门群体:

湛江群体:

北海群体:

防城港群体:

根据前2个判别函数值绘制散点图(图5),由图5可知,虎门和厦门两群体重叠严重,金门、防城港、北海3个群体相距较近,湛江个体则较为分散,与其他各个群体都有一定重叠。

表4 棱梭群体量度指标判别分析结果Tab.4 Results of discriminant analysis on metrical characteristics of C.affinis
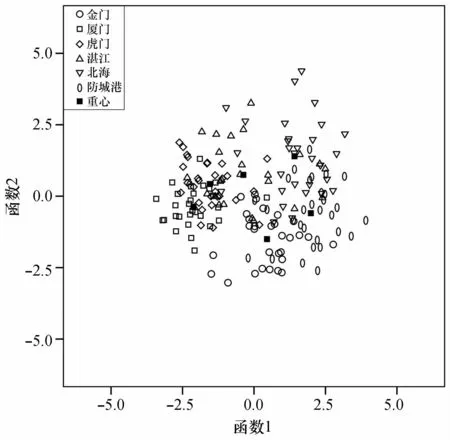
图5 基于量度指标的前两个判别函数值散点图Fig.5 Scatter plots based on the first two discriminant functions of metrical characteristics
对6个棱梭群体28组量度指标进行单因子方差分析,对具有方差齐性和不具有方差齐性的变量分别采用LSD法和Tamhane’s T2法分析,各组变量在0.05水平上显著性差异。结果表明,除了变量第一背鳍基长/体长、吻长/头长、尾柄长/尾柄高、D2-4/体长、D3-5/体长、D4-5/体长和D5-6/体长,不同群体之间的其他21个量度指标的均值都存在显著差异。结果显示,6个群体存在显著差异的指标个数较多,其中,厦门群体与金门、北海和防城港群体之间均有11个指标存在显著差异,为最多;湛江与金门群体之间仅有1个指标(即体高/体长)存在显著差异,为最少。差异系数依据Mayr等(1953)提出的75%识别与划分规则[28]求得,用差异系数大于等于1.28的亚种差异标准来衡量几个群体间的差异程度,结果显示所有两两群体之间、所有量度指标的差异系数小于1.28,介于0.000 3~1.035 1之间。
2.2 棱梭耳石形态
为了检验棱梭左右矢耳石的差异性,使用配对样本t检验,对6个棱梭群体样本的左右矢耳石(凹面)的8个原始耳石尺寸指标进行差异性分析。结果显示棱梭左右耳石差异不明显(p>0.05)。因此,在后续的分析中统一对棱梭的左耳石进行分析。
对6个棱梭群体的85个耳石形态指标进行主成分分析,结果显示特征值大于1的前23个主成分的累积贡献率为79.290%,其中前9个主成分的累积贡献率超过50%,为52.484%。第一主成分贡献率为11.841%,第二主成分贡献率为8.364%,第三主成分贡献率为6.258%。根据前两个主成分的得分情况绘制散点图,结果显示,湛江群体多位于散点图纵轴的正半轴,北海群体多位于散点图纵轴的负半轴。其他的群体则没有呈现出明显的分布倾向(图6)。
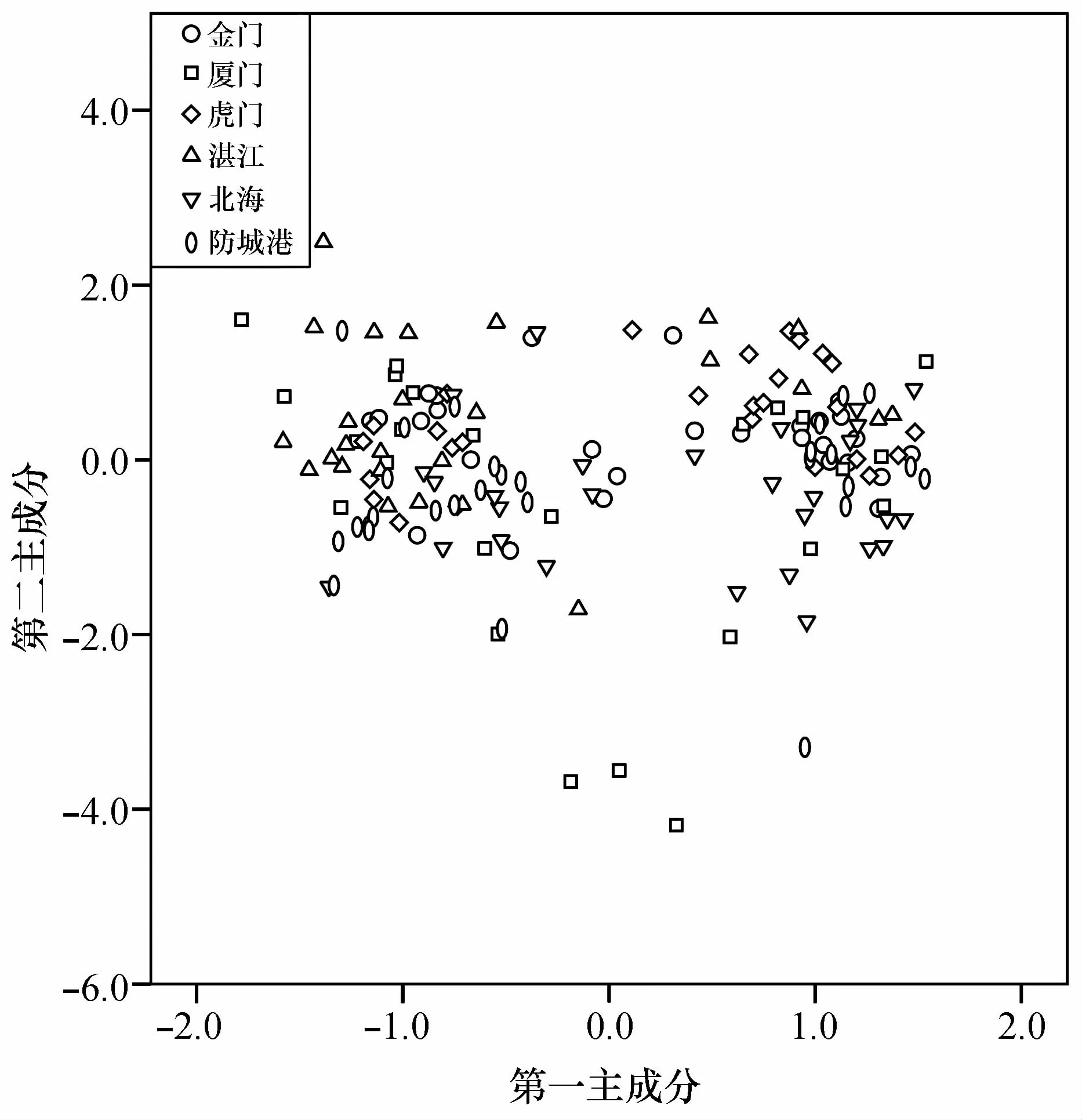
图6 耳石形态指标的前两个主成分得分散点图Fig.6 Scatter plots based on scores of the first two otolith morphological characteristics from PCA
对6个棱梭群体85个耳石形态指标的平均值进行聚类分析,构建平均联结的树状图。图中显示金门群体和北海群体首先聚为一支,然后依次为湛江、厦门、防城港,最后是和虎门聚在一起(图7)。
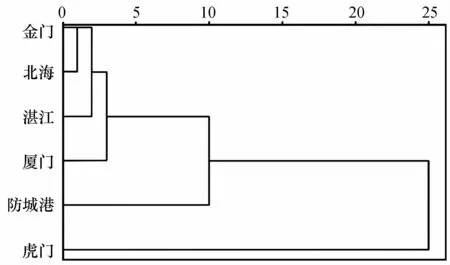
图7 基于耳石形态特征的棱梭群体聚类关系Fig.7 Dendrogram of the relationship of six C.affinis populations based on otolith morphological characteristics
对6个棱梭群体的85个耳石形态指标进行判别分析。结果显示,6个棱梭群体判别正确率仅在60.0%~78.6%之间,综合判别正确率为69.3%。其中判别正确率最高的是金门群体,为78.6%,防城港群体判别正确率次之,为75.9%,厦门群体判别正确率最低,为60.0%。每个群体都有个体被误判到金门群体、北海群体和防城港群体中(表5)。
根据典则判别函数散点图可知,虎门群体和湛江群体与其他群体有较明显的界线,其他4个群体交叉严重,没有明显的界线(图8)。

表5 棱梭群体耳石形态特征判别分析结果Tab.5 Results of discriminant analysis on otolith morphological characteristics of C.affinis
2.3 讨论
形态学研究将传统形态学和地标形态学相结合,对棱梭的6个群体174尾样品的形态学特征进行比较分析。从棱梭可数特征的实验结果来看,棱梭横列鳞数量为13片,腹鳍鳍棘数量为1条,腹鳍鳍条数量为5条,臀鳍鳍棘数量为3条,臀鳍鳍条数量为9条,第一背鳍鳍棘数量为4条,第二背鳍鳍棘数量为1条,尾鳍鳍条数量为14条,这些指标都为定值,且与1962年南海鱼类志[29]、1963年东海鱼类志[20]等相关分类资料中的记录相一致。不同棱梭群体之间的可数特征差异十分微小,因此可数特征很难作为这6个棱梭群体的区分依据。
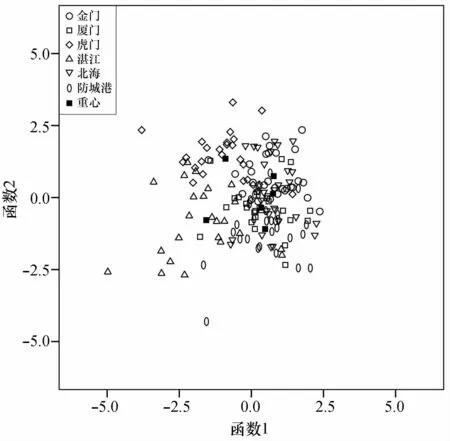
图8 基于耳石形态特征的前两个判别函数值的散点图Fig.8 Scatter plots based on the first two discriminant functions of otolith morphological characteristics
在形态学分析中主成分分析结果显示前8个主成分累积贡献率为65.868%,远小于临界值85.000%,由本结果可以推断这6个棱梭群体形态上的差异不能够单独依靠少数指标来判断[30]。贡献率最大的第1和第2主成分,主要反映了鱼体比例特征、鱼体胸鳍长、腹鳍长、头长和尾柄长等特征。主成分散点图显示金门、虎门、湛江和防城港4个群体的关系密切,厦门和北海分别与其他群体之间略有差异(图3)。聚类分析中金门、虎门、湛江和防城港4个群体的聚类结果说明群体间的主要指标差异的成因与地理距离有一定关联,但是距离很近的金门群体与厦门群体差异较大,相距很远的金门群体与北海群体却首先聚为一支(图4)。金门、北海近海海岸平直、海底地形平缓且水流通畅,其他4个地点包括厦门海岸线曲折、水动力环境复杂[31],再者棱梭成鱼为底层生活鱼类,以摄食底栖生物为主,据此推测金门与厦门近海海底地形地貌和水动力环境等非生物环境差异以及其导致的饵料组成的差异[32]可能是这两个棱梭群体形态差距大的主要原因,海底地形地貌和水动力环境也解释了金门与北海群体的高相似度。
形态学判别分析结果显示6个棱梭群体判别正确率在57.7%~93.1%之间,综合判别正确率为75.9%。结合前两个判别函数值绘制的散点图可得出如下结论:虎门和厦门两个棱梭群体关系密切,湛江与其他各群体均存在一定相似度(图5)。通过以上数据分析可知由形态学特征得到的分类函数并不能精确地将6个群体的棱梭分开。单因素方差分析显示湛江棱梭群体与其他群体在形态上存在显著差异的量度指标较少,而其他两两群体之间的形态差异指标较多,说明这些棱梭群体间出现了一定程度的形态学差异。其中,湛江与金门群体之间仅有1个指标存在显著差异,为最少。这与判别分析结果共同表明了地理位置位于6个地点中间的湛江群体与其他群体相似度高。通过差异系数分析发现,以上群体间量度指标的差异系数介于0.000 3~1.035 1之间,根据Mayr等提出的临界值1.28判断,这6个棱梭群体形态学差异并未达到亚种水平[28]。
对棱梭左右矢耳石(凹面)的6个尺寸指标进行配对t检验,结果表明棱梭左右矢耳石差异不显著,表明棱梭耳石在种内群体间具有高度同质性[2]。对6个棱梭群体163个耳石样本的85个耳石形态学参数进行主成分分析、聚类分析和判别分析,结果与形态学分析结果相近。主成分分析结果显示前23个主成分(特征值大于1)的累积贡献率为79.290%,说明不能使用少数因子判断6个群体的差异。有学者对中国沿海黄鲫(Setipinna tenuifilis)[2]和斑尾复鰕虎鱼(Synechogobius ommaturus)[33]不同地理群体的耳石形态差异进行研究,发现均能用少数因子判断群体间耳石形态的差异,造成研究间差别的原因可能与研究的鱼种、群体间地理远近程度等因素有关。基于第一、第二主成分的得分散点图显示6个群体的耳石形态差异不明显(图6)。聚类分析结果显示金门与北海群体最为相似,这一点与形态学研究结果相同(图7)。6个群体判别正确率仅在60.0%~78.6%之间,综合判别正确率为69.3%。基于前两个判别函数的散点图表明,虎门和湛江群体与其他群体有相对明显的界线(图8)。
3 结论
本研究结合传统形态学、地标形态学和耳石形态学,使用多元统计分析方法对中国东南沿海的6个棱梭地理群体的差异进行研究,鱼体形态和矢耳石形态的分析结果基本一致。两者共同显示各群体间存在一定差异,且聚类分析结果均表明各群体间的差异与地理距离有关联,此外栖息地环境、饵料组成和海流等可能是导致形态学差异和耳石形态差异形成的主要因素。同时,海流的通畅会使得各水域海水温度、盐度等理化因子相似,这会减弱群体间形态和耳石形态的差异,群体间的交流也是差异减弱的一个重要因素。通过两种方法的结合,更好地揭示了地理群体间的关系,丰富了群体间差异研究的案例,为棱梭种质资源的保护和利用提供了一定的研究基础。
